
Abstract
Four hundred and thirty Salmonella isolates, recovered from various food-animal products, were tested for nalidixic acid resistance, plasmid-mediated quinolone resistance, and genetic relationship. One hundred fifteen isolates (113 Salmonella serovar Enteritidis and two Salmonella serovar Typhimurium isolates) of 430 (26.7%) Salmonella isolates exhibited nalidixic acid resistance. Polymerase chain reaction screening for qnrA, qnrB, qnrS, qepA (encoding fluoroquinolones resistance) and rmtB (encoding aminoglycosides resistance) showed that 5 (1.16%) isolates were positive for qnr- and qepA-type genes, and the aac(6′)-Ib-cr gene was observed in two (1.7%) Enteritidis isolates concomitantly with qnrA or qnrB. The co-occurrence of qepA and rmtB in one Typhimurium isolate is noteworthy. Pulsed-field gel electrophoresis revealed a high genetic homogeneity of nalidixic-resistant isolates and the persistence of clonal clusters over 4 years in different regions in Tunisia and from various food-animal products. To the best of our knowledge, this is the first report of co-occurrence of qepA and rmtB in a Salmonella strain.
Introduction
Antimicrobial agents are not essential for the treatment of most Salmonella infections. However, treatment of severe infections caused by Salmonella spp. is necessary (Rice et al., 2003; Guerrant et al., 2001). Broad-spectrum cephalosporins are commonly used to treat invasive infections or severe diarrhea (Guerrant et al., 2001). However, cephalosporin-resistant Salmonella isolates in humans or animals have frequently been reported and have become increasingly common worldwide (Dierikx et al., 2010; Zou et al., 2012). Fluoroquinolones are one of last treatment options for Salmonella infection caused by multidrug-resistant isolates (Guerrant et al., 2001). However, in recent years, fluoroquinolone resistance has been increasingly reported throughout the world, especially in extended-spectrum β-lactamase–producing isolates of human or animal origin (Paterson, 2006; Lo et al., 2012). Resistance to fluoroquinolones is typically mediated by alterations in the target enzymes DNA gyrase (GyrA and GyrB) and topoisomerase IV (ParE and ParC) or by changes in drug entry and efflux (Ruiz, 2003; Robicsek et al., 2006; Cattoir and Nordmann, 2009). The emergence of plasmid-mediated quinolone resistance (PMQR) was complicated in the last 10 years (Cattoir and Nordmann, 2009; Strahilevitz et al., 2009). The first PMQR mechanism involves Qnr-type pentapeptide proteins QnrA (seven alleles), QnrB (48 alleles), QnrS (six alleles), QnrC (one allele), and QnrD 1 (one allele), which confer quinolone resistance by binding to target sites in DNA gyrase and topoisomerase IV and protect them from quinolones (Cattoir and Nordmann, 2009; Strahilevitz et al., 2009; Ruiz et al., 2012). Enzymatic inactivation of quinolones is the second mechanism, conferred by a variant of an aminoglycoside-modifying enzyme, AAC (6′)-Ib-cr. This variant can modify fluoroquinolones and thus yield fluoroquinolone resistance. The third PMQR determinant, QepA encoded by the qepA gene, is an efflux pump system conferring resistance to hydrophilic quinolones (i.e., norfloxacin and ciprofloxacin) (Cattoir and Nordmann, 2009; Strahilevitz et al., 2009; Ruiz et al., 2012). Aminoglycosides, such as gentamicin, amikacin, neomycin, and apramycin, are widely used in animals to prevent and control bacterial infections (Ungemach et al., 2006; Hur et al., 2012). Aminoside resistance is mainly due to modification of the target by mutation in ribosomal proteins or acquisition of various aminoside-modifying enzymes (N-acetylation, O-nucleotidylation or O-phosphorylation) (Doi et al., 2007). However, recently, a new mechanism conferring high-level resistance to all 4,6-disubstituted deoxystreptamine aminoglycosides has been reported. This mechanism is mediated by plasmid-encoded 16S rRNA methylases (armA, rmtA, rmtB, rmtC, rmtD, npmA, and rmtE) and has been increasingly reported in Enterobacteriaceae (Yamane et al., 2005; Doi et al., 2007).
In 2011, our laboratory—the National Reference Laboratory for Salmonella, Shigella, and Vibrio spp. at Pasteur Institute of Tunis—initiated a study including almost all Salmonella enterica serovar Enteritidis and Typhimurium isolates in Tunisia from food-animal products during the period from 2008 to 2011. According to our National Reference Center, these two serotypes were the top serotypes isolated from food-animal products during the last few years in our country (Ben Aissa et al., 2007). The aim of this study was to collect retrospective information on the emergence and occurrence of quinolone resistance in Enteritidis and Typhimurium isolates from different sources of food-animal products in Tunisia, to examine the occurrence of PMQR genes, and to establish the genetic relatedness of these isolates.
Materials and Methods
Salmonella isolates
A total of 430 Salmonella enterica isolates recovered from different food-animal products (chicken: 251, red meat: 126, seafood: 53) in Tunisia during the period 2008–2011 were submitted to our National Reference Laboratory for identification, serotyping, and genetic characterization (Table 1). Each isolate was provided with epidemiological data (source and year of isolation). All of the isolates were suspended in Luria-Bertani broth supplemented with 20% glycerol and stored at −80°C until analysis.
PMQR, plasmid-mediated quinolone resistance; MIC, minimum inhibitory concentration; NAL, nalidixic acid; CIP, ciprofloxacin; PFGE, pulsed-field gel electrophoresis; TET, tetracycline; OFX, ofloxacin; SSS, sulfonamide; SXT, trimethoprim–sulfamethoxazole; TIC, ticarcillin; AMX, amoxicillin; AMP, ampicillin; KAN, kanamycin; STR, streptomycin; CAZ, ceftazidime; CTX, cefotaxime; CEF, cephalothin; CHL, chloramphenicol; GEN, gentamicin.
Serotyping
Salmonella strains were positively identified and serotyped according to the Kauffmann–White scheme with the use of antiserum (Bio-Rad, Marnes-la-Coquette, France). Serotyping was performed by slide agglutination to identify the somatic O antigens and flagellar H antigens.
Antimicrobial susceptibility testing
Antimicrobial susceptibility testing was done using standard methods (disc diffusion method) using Mueller–Hinton agar, and interpreted according to the guidelines of the Antibiogram Committee of the French Society for Microbiology guideline (Soussy et al., 2010
MIC breakpoints used for susceptibility and resistance to nalidixic acid were ≤8 mg/L and ≥16 mg/L as recommended by CA-SFM; whereas for ciprofloxacin MICs ≤0.5 mg/L and >1 mg/L classified susceptible and resistant isolates, respectively, as recommended by European Committee on Antimicrobial Susceptibility Testing (
Screening procedure for PMQR genes
Only nalidixic acid–resistant isolates were included in this study. Rapid DNA preparation was performed by a boiling technique that includes a heating step at 100°C of a single colony in distilled water followed by a centrifugation step of the cell suspension as previously described (Al-Gallas et al., 2002). Screening for the qnrA, qnrB, and qnrS genes was carried out by multiplex polymerase chain reaction (PCR) as described previously (Cattoir et al., 2007). The used primers were able to amplify internal fragments of qnrA1 to qnrA6, qnrB1 to qnrB8, and qnrS1 to qnrS2, which are the prevalent alleles found in Enterobacteriaceae. The aac(6′)-Ib and qepA genes were amplified as previously described (Park et al., 2006; Yamane et al., 2008). Positive controls were used in all PCR experiments (Abbassi et al., 2010). aac(6′)-Ib-positive products were subjected to digestion with BstCI (New England Biolabs) to identify aac(6′)-Ib-cr variants. The rmtB gene, conferring resistance to aminoglycosides, was amplified in all quinolone-resistant isolates as reported previously (Doi et al., 2007).
Molecular typing by PFGE
DNA purification for macrorestriction analysis, restriction enzyme digestion, and pulsed-field gel electrophoresis (PFGE) were performed essentially as described elsewhere (Hunter et al., 2005; Ben Aissa and Al-Gallas, 2008). S. cholerasuis ATCC14028 strain was used as a control. PFGE was performed on a 1% agarose gel (Bio-Rad) using a CHEF DR III apparatus (Bio-Rad) in 0.5x Tris–borate–EDTA buffer at 14°C with 6 V/cm at a field angle of 120°. For better DNA bands separation, the total running time of electrophoresis was 19 h with two successive electrophoresis programs, a first running program of 8.5 h, with initial switching time of 7 s, to final switching time of 12 s, automatically followed by a second running program of 10.5 h with initial switching time of 20 s to final switching time of 40 s. After the electrophoresis, the gels were stained with ethidium bromide, and DNA was visualized under ultraviolet light. A lambda DNA ladder with a size range of 48.5 kb to 1 Mb (Amersham Biosciences, Buckinghamshire, UK) was used as a DNA size standard. The DNA patterns were compared using the BioNumeric software (Applied Maths, Kortrijk, Belgium). The relatedness of different patterns was analyzed by cluster analysis using the Dice coefficient for band matching with 1% position tolerance and the unweighted-pair group method with arithmetic average.
Results
Prevalence of quinolone resistance and antibiotic susceptibility
According to the number of isolates per year from different food-animal products we showed the following numbers (360 Enteritidis isolates, 70 Typhimurium isolates): 2008, 66 Enteritidis (chicken: 57, seafood: 9) and 13 Typhimurium (chicken: 11, red meat: 2); 2009, 89 Enteritidis (chicken: 54, red meat: 30, seafood: 5), and 11 Typhimurium (chicken: 8, seafood: 3); 2010, 134 Enteritidis (chicken: 53, red meat: 63, seafood: 18) and 31 Typhimurium (chicken: 17, red meat: 9, seafood: 5); 2011, 71 Enteritidis (chicken: 47, red meat: 13, seafood: 11) and 15 Typhimurium (chicken: 4, red meat: 9, seafood: 2).
Among 430 Salmonella isolates, 115 (27%) were resistant to nalidixic acid (NAL) as shown by MIC values or antibiogram, of which 20 were classified as ciprofloxacin resistant by MIC (>0.5 μg/mL). Antimicrobial susceptibility testing revealed that 31.4% (113/360) of Enteritidis were NAL resistant and only two of 70 (2.8%) Typhimurium isolates were NAL resistant. A gradual increase in the prevalence of NAL-resistant Salmonella serovar Enteritidis isolates was observed during the 4 years, being 16.6% in 2008 and 57.7% in 2011 (Table1). MICs of the 115 NAL-resistant Salmonella ranged from 32 μg/mL to >2048 μg/mL for NAL and from 0.003 μg/mL to 128 μg/mL for ciprofloxacin. The majority of isolates showed reduced susceptibility to ciprofloxacin (MIC≥0.125 mg/L), with the exception of 11 isolates that had ciprofloxacin MICs ranging from 0.003 μg/mL to 0.032 μg/mL. It is noteworthy that the majority of quinolone-resistant isolates were isolated from chicken. Furthermore, multiple drug resistance to three or more antimicrobial agents was detected in 17.4% (20/115) of these NAL- resistant isolates (Table 1). The “ACSSuT” penta-resistance pattern was observed in two Salmonella serovar Typhimurium isolates.
Occurrence of qnr, qepA, acc(6′)-Ib-cr, and rmtB genes
qnr genes were detected in five isolates (4.3%). These qnr genes were as follows: qnrA (one Enteritidis), qnrB (three Enteritidis), and qnrS (one Enteritidis) among the 115 strains resistant to NAL. The aac(6′)-Ib-cr gene was observed in two (1.7%) Enteritidis isolates concomitantly with qnrA or qnrB. The qepA gene was observed in one Typhimurium isolate in association with rmtB gene. It is worth noting that no isolates obtained from red meat or from clams harbored any PMQR genes.
Clonal relationship of Salmonella isolates
The 115 NAL-resistant Salmonella strains were analyzed by PFGE. XbaI digestion yielded 11–22 DNA fragments ranging in size from approximately 50 kb to 600 kb. DNA degradation was observed for three Enteritidis isolates because of nuclease activity, and the remaining 112 isolates were classified in 16 PFGE clusters (Fig. 1). Each cluster contained closely related isolates that differ by a single genetic event, indicated by two-to-three band differences (Tenover et al., 1995). Three major clusters (A, B, and D) encompassing 87 isolates were observed. The PFGE pattern A (31 isolates) and B (45 isolates) were subdivided to subtypes presenting closely related isolates. The similarity between PFGE A and B was 72%. Ten unique profiles were also observed. The qnrB-positive and qnrS-positive Enteritidis isolates were of pulsotype A1; however, qnrA- and qepA-harboring isolates were singletons.
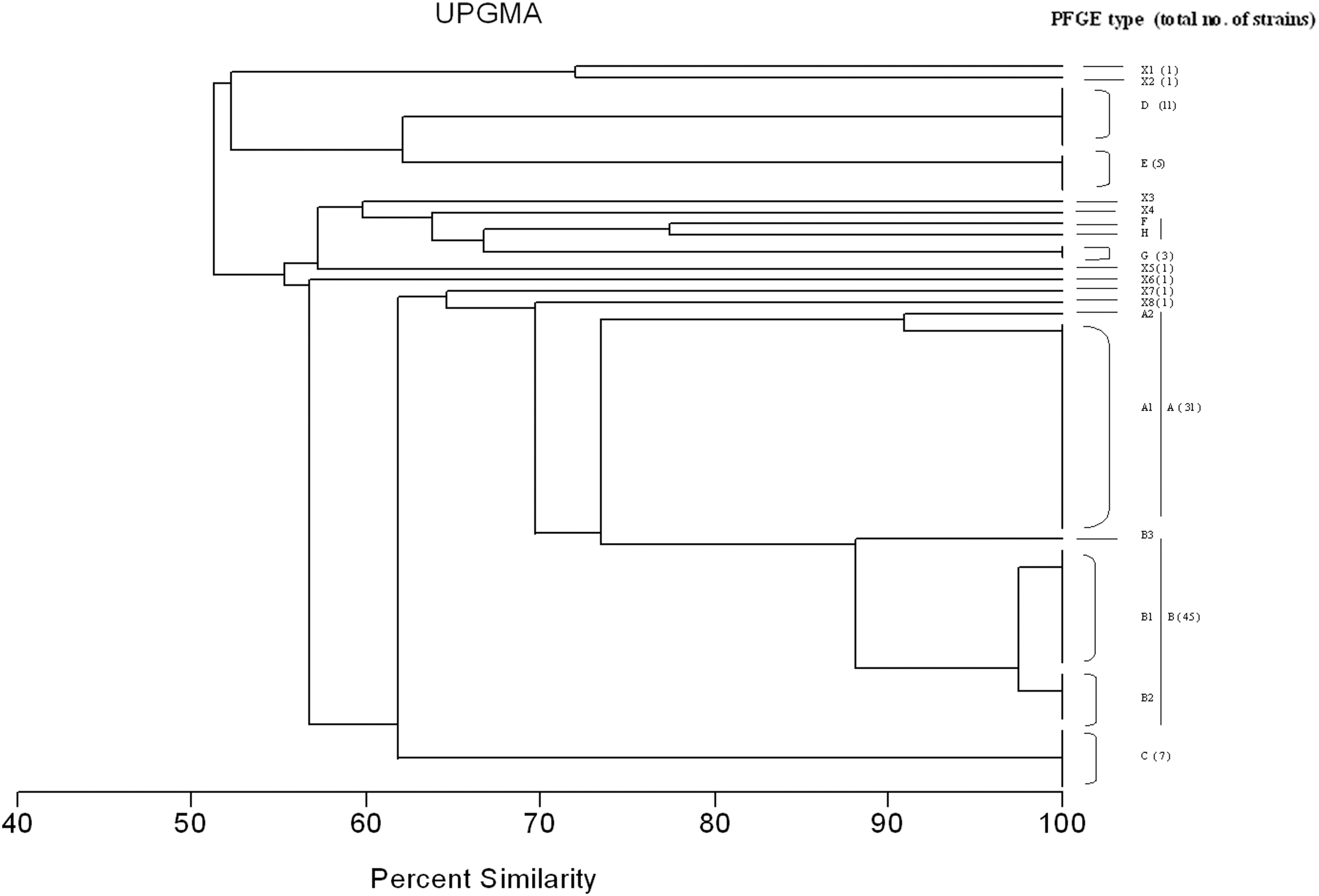
Dendrogram showing percent similarity calculated by the Dice similarity index of pulsed-field gel electrophoresis (PFGE) restriction endonuclease digestion profiles among the 115 quinolone-resistant Salmonella enterica serovars Enteritidis and Typhimurium strains isolated from food-animal products. The different patterns and number of strains are indicated. UPGMA, unweighted-pair group method with arithmetic mean.
Discussion
An increasing rate of antimicrobial resistance in Salmonella has been reported worldwide (Van et al., 2012), and resistance to combinations of several classes of antimicrobials has led to the emergence of multidrug-resistant strains. In Tunisia, little is known about the occurrence of antibiotic resistance in Salmonella from food-animal products as well as from humans (Guedda et al., 2009; Abbassi-Ghozzi et al., 2011). In this report, we focus on quinolone resistance in Salmonella isolated in Tunisia during a period of 4 years, which can give us a reliable data for the real epidemiological situation. The majority of Salmonella isolates were of Enteritidis serotype (360/430); indeed, this serotype has emerged as an important serotype isolated from chicken and all other food-animal products in Tunisia, as well as throughout the world (Ben Aissa et al., 2007; Guedda et al., 2009; Abbassi-Ghozzi et al., 2011; Soufi et al., 2012; Van et al., 2012). NAL resistance was observed in 27% (115/430) isolates and mainly in Salmonella serovar Enteritidis (98.2%, 113/115). Similar rates have been reported by other reports (Vo et al., 2010; Wannaprasat et al., 2011; de Jong et al., 2012).
Owing to the absence of an ecological barrier of human and animal ecosystems as well as the zoonotic property of Salmonella, the transit of quinolone-resistant Salmonella isolates to humans is a major health problem. This problem could even be more significant among humans with critical immunological status and would require antibacterial therapeutic intervention. Resistance to quinolone was clearly showed by antibiogram, and MICs were two- to 128-fold over the breakpoint (16 μg/mL). However, only five isolates were ciprofloxacin resistant by antibiogram and showed MICs between 32 μg/mL and 128 μg/mL. The remaining isolates showed two populations. The first population was isolates with full susceptibility to ciprofloxacin and presenting MICs ranging between 0.003 μg/mL and 0.032 μg/L, and the second population was presented by isolates with ciprofloxacin MICs ranging from 0.125 to <0.5 μg/mL and considered as nonwild susceptible isolates or intermediately resistants. Theoretically, NAL resistance also means cross-resistance to fluoroquinolone compounds, due to identical target site (Ruiz, 2003; Strahilevitz et al., 2009; Ruiz et al., 2012). In our study, PMQR genes were detected in six isolates. Owing to many limitations, we were unable to do sequencing of qepA and rmtB genes and to study their genetic background; however, positive amplifications were observed in three more separate PCR amplifications yielding the expected sizes of DNA amplicons. Indeed, the qnrB was identified in three Enteritidis; however, qnrA and qnrS1 each were identified in one Enteritidis isolate. The aac(6′)-Ib-cr gene was observed in two (1.7%) Enteritidis isolates concomitantly with qnrA or qnrB. The occurrence of qnr-type in Salmonella from animal or food-animal products has been reported previously with low rates (Veldman et al., 2011), in contrast to other reports showing total absence of any PMQR in NAL-resistant Salmonella (Kim et al., 2011; Thai et al., 2012). The qepA gene was found in one Typhimurium isolate. To the best of our knowledge, this is the second report of qepA gene occurrence in Salmonella spp. after Lunn et al. (2010), and the first report of concurrent occurrence of qepA and rmtB genes in Salmonella. Indeed, despite high reported prevalence of this gene in Enterobacteriaceae of human or animal origin (Paterson, 2006; Cattoir and Nordmann, 2009), it was not found in a large survey of non-Typhi Salmonella enterica isolates collected in the United States from 1996 to 2003 and in recent reports from Europe (Sjölund-Karlsson et al., 2010; Veldman et al., 2011). The aac(6′)-Ib-cr gene was observed in two (1.7%) Enteritidis isolates. This gene has been reported previously in Salmonella isolates as well as in other Enterobacteriaceae of animal or human origins (Veldman et al., 2011). Taken together, it seems at present in Tunisia that dissemination of PMQRs among Enteritidis and Typhimurium isolates from food-animal products and mainly from chicken may remain restricted. The spread of PMQR among Salmonella isolates of animal origin could have serious consequences for fluoroquinolones treatment of nontyphoid Salmonella infection in humans and animals. Furthermore, NAL resistance in the PMQR-free isolates is likely due to mutations in gyrA, parC, or parE as well as efflux pumps or the decreased expression of outer membrane porins that remain the principal mechanisms of quinolone resistance in Salmonella (Kozoderović et al., 2012).
Genetic relatedness of the isolates was investigated by PFGE. Three major genotypes that were found—designated PFGE A, PFGE B, and PFGE D—each contained 31, 45, and 11 isolates, respectively; a minor cluster (genotype C) contained seven isolates. The two Typhimurium isolates had two different profiles; one of them clustered with three Enteritidis isolates (pulsotype G). Similar results of low genetic heterogeneity of quinolone-resistant Salmonella strains have been reported (Campioni et al., 2012). Interestingly, some genotypes (PFGE patterns) contained isolates from chicken, clams or red meat (beef) which means cross dissemination of specific genotypes between different ecological niches. Moreover, isolates of the same pulsotype showed various antibiotic-resistance profiles and genes content. These findings might be explained by clonal dissemination of specific clones with independent steps of acquisition or loss of genetic markers encoding antibiotic resistance or other traits not investigated in our work. From the genetic point of view, it seems that the whole genome (chromosome) of Enteritidis undergoes a low genetic variation in time and that some clones are well established in their ecological niche.
Conclusions
In conclusion, 26.7% of Salmonella isolates, recovered from food-animal products, were resistant to NAL, and this resistance was mainly observed in Salmonella serovar Enteritidis (113/115 isolates). To the best of our knowledge, we report for the first time the concurrent occurrence of qepA and rmtB genes in a Salmonella isolate. Genetic investigations to characterize the genetic background of qepA and rmtB genes and to establish whether they are genetically linked in a mobile genetic element as reported in other Enterobacteriaceae isolates is needed, and is under investigation. Plasmid-mediated quinolone resistance determinants were rare in our collection; therefore, mutations in gyrA and parC genes would be the main molecular mechanism of quinolone resistance in reported isolates. PFGE revealed a high genetic homogeneity of nalidixic-resistant isolates. Furthermore, it revealed the persistence of clonal clusters over 4 years in different regions in Tunisia and from various food-animal sources.
Footnotes
Acknowledgments
We thank Amna Al-Gallas, MD, International Fellow, Massachusetts Eye and Ear Infirmary (hospital affiliated with Harvard Medical School), Boston, Massachusetts, for helping with the English revision and editing of the article. We thank all public health and hygiene laboratories in Tunisia for their continued cooperation and reporting. All authors contributed equally to this article.
Disclosure Statement
No competing financial interests exist.