
Abstract
Escherichia coli can contaminate raw milk during the milking process or via environmental contamination in milk-processing facilities. Three bacteriophages, designated EC6, EC9, and EC11, were investigated for their ability to inhibit the growth of three strains of E. coli in ultrahigh-temperature (UHT) treated and raw bovine milk. A cocktail of the three phages completely inhibited E. coli ATCC 25922 and E. coli O127:H6 in UHT milk at 25°C and under refrigeration temperatures (5–9°C). The phage cocktail produced similar results in raw milk; however, E. coli ATCC 25922 and O127:H6 in raw milk controls also declined to below the level of detection at both temperatures. This observation indicated that competition by the raw milk microbiota might have contributed to the decline in viable E. coli cells. A cocktail containing EC6 and EC9 completely inhibited E. coli O5:H-, an enterohemorrhagic strain, in UHT milk at both temperatures. In raw milk, the phage cocktail initially inhibited growth of E. coli O5:H- but regrowth occurred following incubation for 9 h at 25°C and 144 h at 5–9°C. In contrast to the other E. coli strains, O5:H- was not inhibited in the raw milk controls. This study demonstrates that bacteriophages are effective biocontrol agents against E. coli host strains in UHT and raw bovine milk at various storage temperatures.
Introduction
Bacteriophages have long been negatively associated with the dairy industry. Of the approximately 500 million tons of milk produced globally each year, an estimated one third is processed into fermented products (Brussow, 2001). The presence of dairy bacteriophages in milk leads to interruption of the fermentation of lactose by starter cultures, causing delays in production and alteration of product quality. Despite this negative association, there has been increasing interest shown in recent years in using bacteriophages to control bacterial pathogens in dairy products. Given the high host-specificity of phages, the risk to starter cultures is considered remote.
Dairy applications for bacteriophages investigated to date include the control of Listeria monocytogenes in chocolate-flavored milk (Guenther et al., 2009), Cronobacter sakazakii in reconstituted powdered infant formula (Kim et al., 2007), and Staphylococcus aureus in pasteurized and raw milks (O'Flaherty et al., 2005; Garcia et al., 2009). Currently, no studies have investigated bacteriophages for the biocontrol of milkborne Escherichia coli.
E. coli can be isolated from many foods of animal origin. While outbreaks of foodborne illnesses attributed to pathogenic E. coli have been associated primarily with minced beef, many outbreaks have been attributed to consumption of contaminated fresh produce and dairy products, including raw milk (Tuttle et al., 1999; Ferguson et al., 2005; De Schrijver et al., 2008; Guh et al., 2010). Although infection with E. coli typically causes self-limiting illness, the effects of Shiga toxin–producing E. coli (STEC), such as O157, can be more severe, leading to serious complications such as hemolytic uremic syndrome in 10–15% of cases (Pennington, 2010).
Raw milk consumption appears to be increasing in popularity in developed countries, which is likely associated with the desire to consume natural, unprocessed foods. This is despite the well-documented risks associated with this practice (Oliver et al., 2009). The risk to raw milk consumers of developing E. coli–associated illness is heightened, given the low infectious dose (10–100 cells) of some pathogenic strains (Feng, 2001). Detection rates of STEC in raw milk vary. Klie et al. (1997) detected STEC in 3.9% of raw milk samples tested, while Altalhi and Hassan (2009) had a detection rate of 9.1%.
While many studies have investigated phages for controlling E. coli O157 in foods (O'Flynn et al., 2004; Abuladze et al., 2008; Viazis et al., 2011), fewer studies have focused on other serotypes. Australia has a low incidence of E. coli O157 infections compared to other countries including Japan, Argentina, and the United States (Sakuma et al., 2006; Leotta et al., 2008). However, pathogenic E. coli belonging to a range of non-O157 serotypes have been implicated in disease outbreaks in Australia (Combs et al., 2005; Pennington, 2010).
In this study, bacteriophages were investigated for their ability to control three strains of E. coli in ultrahigh-temperature (UHT) treated and raw bovine milk.
Materials and Methods
Media and bacterial cultures
Escherichia coli ATCC 25922, E. coli O127:H6, an enteropathogenic clinical isolate, and E. coli O5:H-, an enterohemorrhagic clinical isolate, were used as primary bacteriophage hosts. Eighteen strains of E. coli, including laboratory strains and pathogenic strains, were used in host range studies (Table 1). The pathogenic strains were clinical isolates provided by the University of Melbourne from their available collection. The E. coli strains used in the study were routinely subcultured on tryptone soya agar (Difco, New Jersey, USA) and tryptone soya broth (Difco), and biocontrol studies were performed using either tryptone soya agar or Chromocult Coliform Agar (Merck, New Jersey, USA). All cultures were incubated at 37°C for 24 h.
Nonpathogenic strain.
Pathogenic type strain ATCC 43895.
Pathogenic strains (clinical isolate).
+, clear plaque; −, no plaque; (+), turbid plaque.
Bacteriophage isolation and characterization
Three lytic bacteriophages—EC6, EC9, and EC11—were isolated from sewage and purified as described by Carey-Smith et al. (2006). Preliminary characterization of the phages was performed by transmission electron microscopy, DNA isolation, and restriction fragment length polymorphism analysis (Shivu et al., 2007). The host range of the phages was determined against a panel of 18 pathogenic and nonpathogenic E. coli using the overlay method (Adams, 1959). Clear plaques indicated lysis of the host strain, and turbid plaques were considered indicative of incomplete lysis.
Preparation of phage cocktails
High-titer stocks were prepared for biocontrol studies as described by Carey-Smith et al. (2006) and titers were determined by plaque assay. Phage cocktails were prepared by combining equal concentrations of phage stocks to a working concentration of approximately 1×1010 plaque-forming units (PFU)/mL. A cocktail containing EC6, EC9, and EC11 was used in studies investigating inhibition of E. coli ATCC 25922 and E. coli O127:H6. The cocktail used in studies investigating the inhibition of E. coli O5:H- contained phages EC6 and EC9 only due to limited host range.
Preparation of milk samples
UHT milk was purchased in 100-mL tetra packs. Raw milk samples were collected from a local milk-processing plant and transported on ice to the laboratory. Raw milk samples with aerobic plate counts (APC) of less than 10,000 colony-forming units (CFU)/mL were screened for the presence of E. coli by plating onto Chromocult coliform agar following enrichment in buffered peptone water. Samples found to be negative were used in biocontrol studies and were concurrently tested for APC and E. coli at defined intervals during the course of the studies.
Phage biocontrol of E. coli in UHT and raw milk
Overnight broth cultures of E. coli were centrifuged at 5000×g for 2 min and the pellet was washed and resuspended in PBS. Suspensions were adjusted with PBS to approximately 1×108 CFU/mL by colorimetry and then diluted 100-fold to achieve an approximate concentration of 1×106 CFU/mL. UHT and raw milk samples were inoculated with approximately 1.0×105 CFU/mL of E. coli host and 1.0×109 PFU/mL of phage cocktail or an equivalent volume of PBS (control). Triplicate samples were incubated either at 25°C for 24 h, or in a domestic refrigerator for 168 h. The refrigerator thermostat was set to simulate mild temperature abuse and fluctuated between 5°C and 9°C. Aliquots were removed from each sample at defined intervals and serially diluted in 0.1% buffered peptone water. Dilutions were plated onto tryptone soya agar (for UHT milk) or Chromocult Coliform Agar (for raw milk) and incubated at 37°C for 24 h for viable count determination. Chloroform (1% [vol/vol]) was added to dilutions and phage titers were determined using the overlay method (Adams, 1959).
Statistical analysis
Comparison between means was carried out using paired-samples t-tests. All statistical analyses were performed using Statistical Package for Social Sciences (SPSS) for Windows Version 18.0. Statistical significance was defined at a p value≤0.05.
Results
Bacteriophages
Morphological analysis of phages revealed them to belong to different families within the order Caudovirales. EC6 was classified into the Siphoviridae due to the presence of a long, flexible tail and icosahedral capsid. The morphology of EC9 was similar to T4, with an elongated head and contractile tail, indicating it belonged to the Myoviridae. EC11 appeared to possess a very small tail and an icosahedral capsid and was therefore grouped into the Podoviridae. Restriction endonuclease digestion with SspI indicated that all phages possessed dsDNA genomes.
EC9 possessed the broadest host range of the three phages, infecting pathogenic strains belonging to different serotypes. EC6 and EC9 were able to infect all five nonpathogenic E. coli strains and one pathogen (Table 1).
Phage inhibition of E. coli in UHT milk
UHT milk was first investigated as a model for raw milk that was free from native microbiota. In UHT milk, the phage cocktails reduced E. coli ATCC 25922 to below the level of detection within 3 h of incubation at 25°C (Fig. 1B) The same result was achieved in 3 h for E. coli O127:H6 at 25°C (Fig. 2B). Furthermore, E. coli O5:H- was also reduced to below the level of detection within 3 h at 25°C (Fig. 3B). No regrowth of E. coli was observed in any of the phage-treated milk samples. In contrast, the three E. coli strains increased by up to 4.3 log10 CFU/mL in the control milk samples after 24 h.
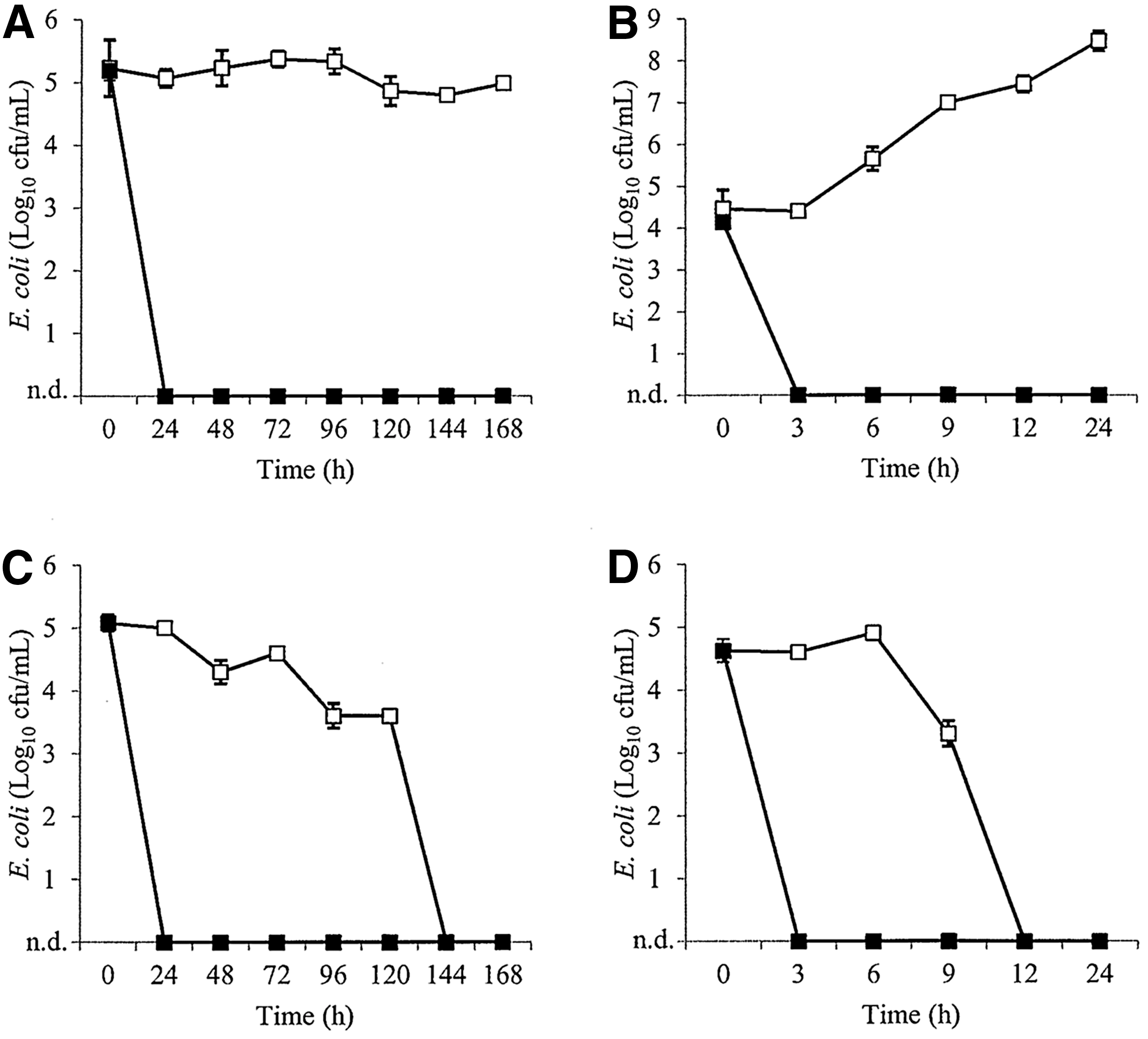
Inhibition of Escherichia coli ATCC 25922 in ultrahigh-temperature-treated milk
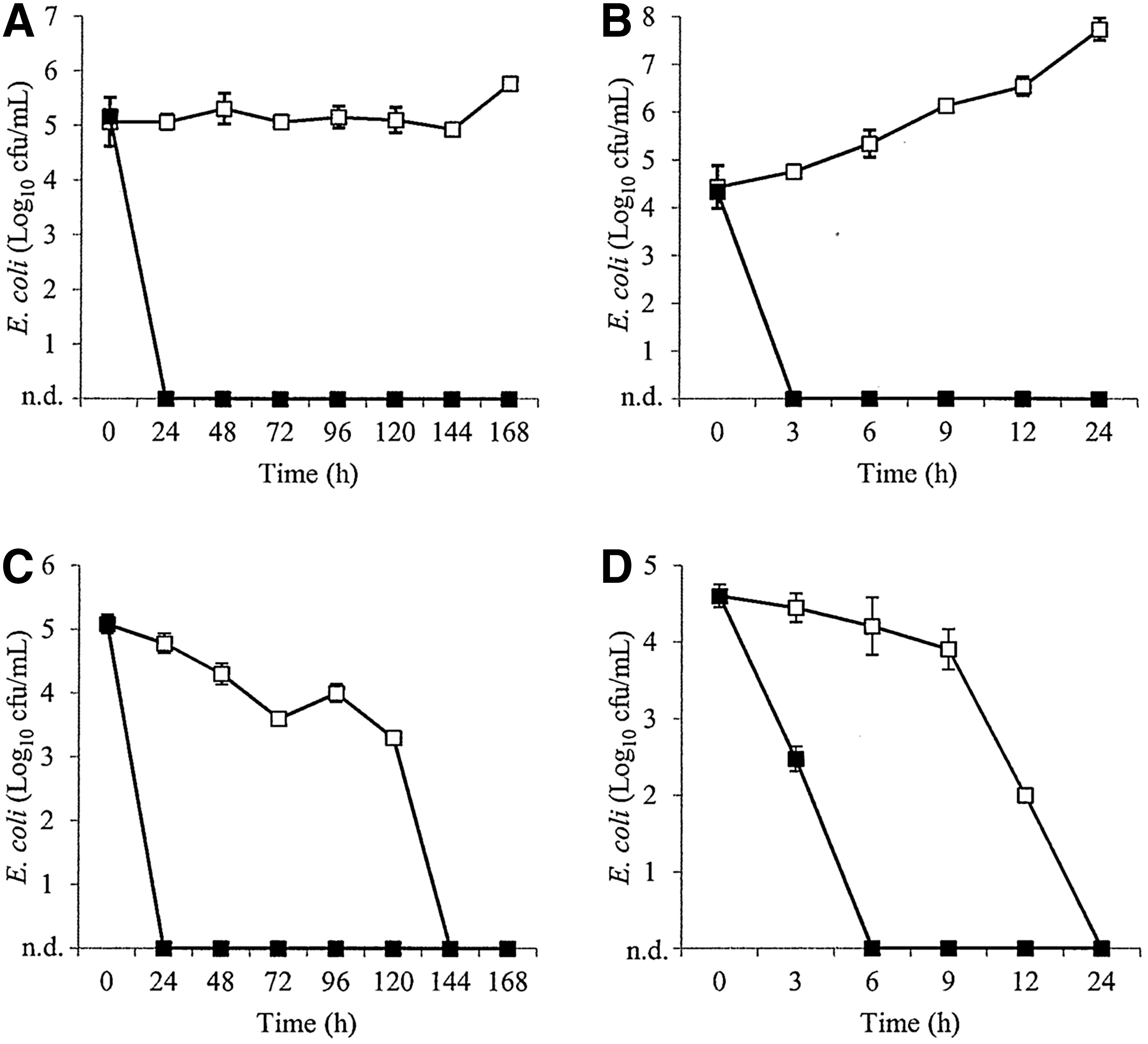
Inhibition of Escherichia coli O127:H6 in ultrahigh-temperature-treated milk
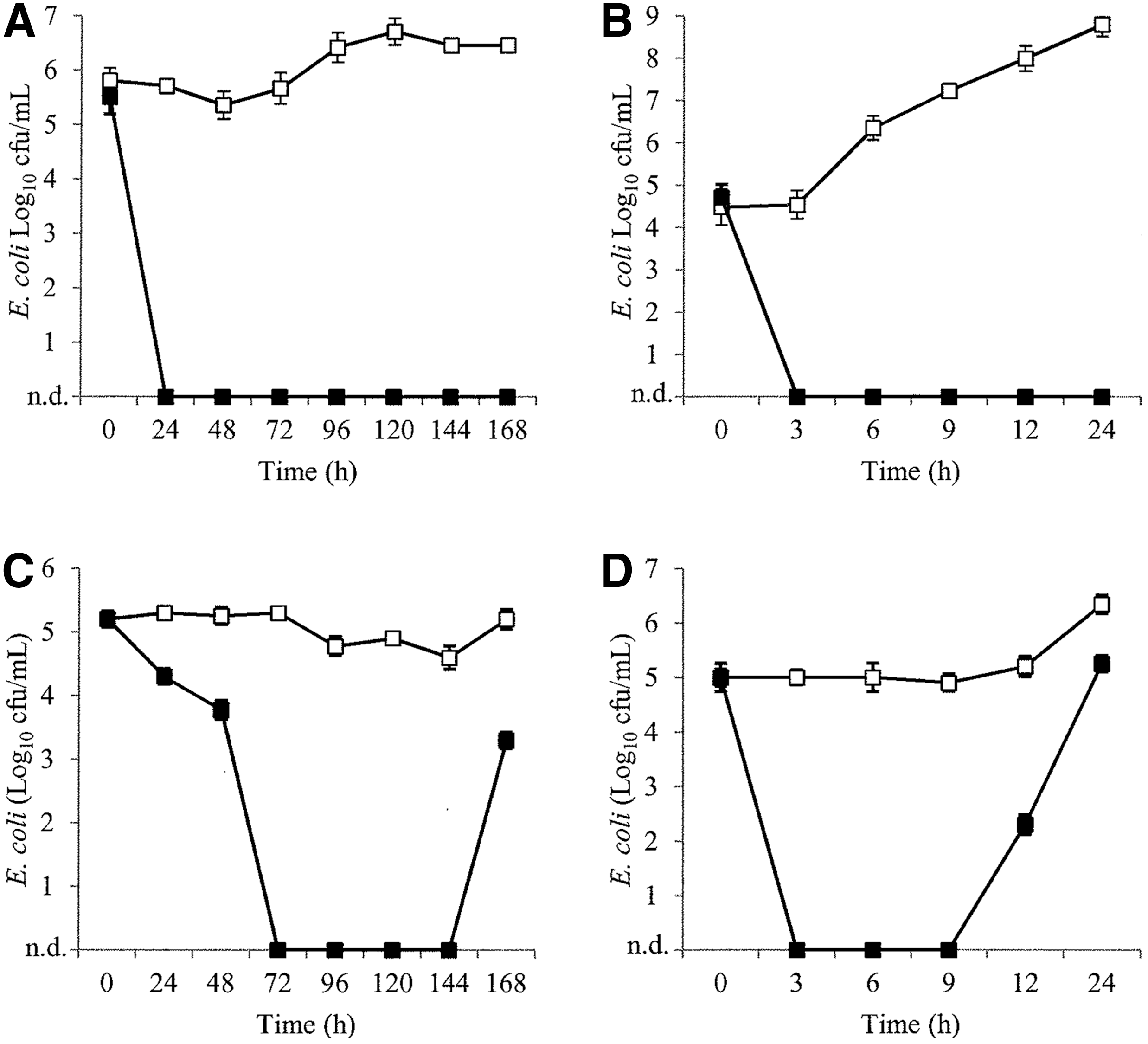
Inhibition of Escherichia coli O5:H- in ultrahigh-temperature-treated milk
In refrigerated samples, the phage cocktails inhibited all three strains of E. coli to below the level of detection within 24 h of incubation, while E. coli in the control milk samples did not significantly change over the 7-day studies (p>0.5) (Figs. 1A, 2A, and 3A). Furthermore, phage titers did not fluctuate by more than 1 log10 PFU/mL over the course of the studies.
Phage inhibition of E. coli in raw milk
The phage cocktail containing EC6, EC9, and EC11 eliminated E. coli ATCC 25922 and O127:H6 in raw milk within 3 h and 6 h of incubation, respectively, at 25°C (Figs. 1D and 2D, respectively). However, elimination of E. coli in the untreated milk samples was also observed within 24 h. This was also observed in refrigerated milk samples, in which E. coli in phage-treated samples was eliminated within 24 h compared to 144 h in the untreated controls (Figs. 1C and 2C, respectively). Although E. coli ATCC 25922 and O127:H6 did not survive in the raw milk controls, the phage cocktail was able to eliminate these strains more rapidly than observed in the untreated milk samples, suggesting that this type of treatment may still be useful where raw milk is to be consumed within a few days of production. Furthermore, phage titers did not fluctuate by more than 0.5 log10 PFU/mL over the course of the studies.
In contrast, E. coli O5:H- remained stable in control raw milk at low temperature and increased by 1.3 log10 CFU/mL at 25°C (Fig. 3D). Despite an initial reduction in E. coli O5:H- in phage-treated raw milk incubated at 25°C, regrowth was observed after 9 h. At low temperature, it took 72 h for E. coli O5:H- to be reduced to below the level of detection, and regrowth was observed after 144 h (Fig. 3C). After 168 h, E. coli O5:H- was 2.3 log10 CFU/mL lower in the phage-treated samples compared to the control. Phage titers remained within 1 log10 PFU/mL of the starting titer after 24 h of incubation at 25°C and after 7 days of incubation under refrigeration. The APC of the raw milk negative control increased by an average of 6 log10 CFU/mL over 168 h of incubation at low temperature, and no E. coli was observed.
Discussion
Phages of the Myoviridae have been reported to possess a wider host range than phages belonging to the Siphoviridae among a range of bacterial hosts including L. monocytogenes (Loessner et al., 1997), Staphylococcus (Pantůček et al., 1998), Vibrio cholerae (Miller et al., 2003), and E. coli (Chibani-Chennoufi et al., 2004). This is supported by results from the current study, with EC9 exhibiting the broadest host range of the three phages isolated. All three bacteriophages were able to infect nonpathogenic strains of E. coli in addition to at least one pathogenic strain. The ability of the phage isolates to be propagated on nonpathogenic hosts is a desirable feature for therapeutic or biocontrol candidates to avoid the potential for transfer of virulence or toxin genes to recipient cells via generalized transduction (Hagens and Loessner, 2010). Furthermore, it would simplify production and increase the safety of phage preparations (Santos et al., 2010). However, the narrow host range of the phages isolated in this study makes them useful for this proof of concept study but unsuitable as “real world” biocontrol candidates.
Phage biocontrol has been shown to be more effective in liquid foods than in solid foods. Guenther et al. (2009) found that a bacteriophage cocktail was able to eliminate L. monocytogenes populations in chocolate milk and mozzarella cheese brine incubated at 6°C over 6 days. On solid foods, diffusion is limited and a greater concentration of phages may be necessary to achieve the same result as in liquid foods. However, liquid foods such as milk may also present unique barriers to phage biocontrol. For example, milk proteins or fat globules may obstruct contact between phages and their target cells, resulting in inactivation of the phages (O'Flaherty et al., 2005; Garcia et al., 2009). The phages used in the current study did not appear to be inhibited by milk components.
It is unknown whether the reductions in E. coli observed in the current study were the result of infection or lysis from without (Callaway et al., 2008). However, many studies have used comparable concentrations of phages and reported findings similar to those of the current study (Abuladze et al., 2008; Guenther et al., 2009). In addition to multiplicity-of-infection, time and temperature also contribute to efficacy of phages in biocontrol applications (Viazis et al., 2011). The phages investigated in the current study did not appear to be affected by suboptimal temperatures as they were effective in controlling their target hosts in UHT milk at both 25°C and under refrigeration.
While the phage cocktail comparably eliminated E. coli ATCC 25922 and O27:H6 in raw milk regardless of incubation temperature, regrowth of E. coli O5:H- in phage-treated milk occurred following 9 h of incubation at 25°C and after 144 h of incubation at 5–9°C. It is likely that E. coli O5:H- was reduced to very low numbers by the phage cocktail that were not detectable by the recovery method used. The phage cocktail used in that study comprised only two phages, compared to the three phages used for the other E. coli strains. In addition, in raw milk studies using other E. coli strains, the host in the control also died out during the study, making the probability of E. coli recurring in the corresponding phage-treated milk unlikely. A limitation of the current study is that the raw milk was not tested for native phages. However, it is unlikely that native phages would be present in sufficient quantity to affect the results significantly.
The results from the current raw milk study are promising given the mixed results reported by others. O'Flaherty et al. (2005) found that Bacteriophage K was able to eliminate S. aureus in heat-treated milk but not raw milk. In a similar study, Garcia et al. (2009) used phages φA5 and φA72 to reduce S. aureus in full cream and semiskimmed raw milk incubated at 37°C. S. aureus in phage-treated milks increased during the study but to significantly lower levels (p<0.05) than in the untreated milks. The concentration of the host bacteria and phage were lower and more realistic than the concentrations used in the current study. The authors acknowledged that lower host cell densities require greater concentrations of phage for sufficient infection to occur and that the antimicrobial activity of the phages may have been reduced due to the slower growth of S. aureus in raw milk compared to heat-treated milk as a result of the presence of competitive microbiota. Furthermore, φA5 and φA72 were temperate phages, which may also have contributed to the lower efficacy in inhibiting S. aureus. In another study by the same group, lytic mutants of φA5 and φA72 were found to be effective in eliminating S. aureus in UHT milk and during curd production using pasteurized milk (Garcia et al., 2007). However, the efficacy of these lytic mutants in raw milk was not investigated.
Over the course of the current studies, phage titers remained within±1 log10 PFU/mL of the starting titer. O'Flaherty et al. (2005) found that Bacteriophage K was inhibited in raw milk due to agglutination of bacterial cells with fat globules, which prevented phage attachment. Similarly, Garcia et al. (2009) reported that the titer of φA5 and φA72 decreased by 1 log10 unit after 10 h of incubation in raw milk, suggesting possible partial inactivation. However, phages of other bacteria have been successful in inhibiting their hosts in raw milk products without significant loss in titer. For example, Modi et al. (2001) were able to recover Salmonella-specific phages from raw milk and raw milk cheese with no loss in titer.
The composition and microbiota of raw milk varies greatly, depending on the breed of cattle, geographic location, season, and milking and handling procedures (Oliver et al., 2005). These factors may contribute to the efficacy of phage biocontrol, and a more extensive study should be undertaken to determine their effect on the reproducibility of the results of this study. The concentration of microbiota in the current study was relatively low, and therefore the effect of a high microbial load should also be assessed. Wilkinson (2001) suggested that phage efficiency in the gastrointestinal system was impeded by the high density of nontarget species (decoys). Only when the host was present in higher numbers than the decoys would the phages be predicted to reach the same levels as in the absence of competitive organisms. The results from the current study are in agreement with this model. The concentration of E. coli initially added to the milk samples was approximately 500-fold higher than the APC calculated at the beginning of the study. All phage-treated milk samples showed a reduction in E. coli within 24 h of inoculation at 5–9°C.
Overall, this preliminary proof of concept study demonstrated that bacteriophages can be effective in controlling E. coli hosts in UHT and raw bovine milk at various storage temperatures. Further investigations should be carried out in order to determine the feasibility of this approach in real settings. Broader host range phages able to infect E. coli pathotypes commonly associated with raw milk should be isolated. Furthermore, the effect of differences in milk composition and microbiota and lower target cell and phage concentrations on the efficacy of the phages must also be determined.
Footnotes
Disclosure Statement
No competing financial interests exist.