
Abstract
To investigate the prevalence of nine virulence and toxin genes of Campylobacter, a total of 90 Campylobacter strains including 48 C. jejuni and 42 C. coli were recovered from chicken feces by cultivation methods. The isolates were identified on the basis of polymerase chain reaction (PCR) detection of 16SrRNA and multiplex PCR for determining two species. For confirmed strains, PCR was carried out for the presence of virulence genes using specific primers. Data were analyzed by SPSS software, version 12.0.1. The cadF gene and three genes associated with cytolethal distending toxin were present in 100% of isolates. Plasmid virB11 gene was not found in any of the Campylobacter isolates, and the prevalence of pldA, wlaN, iamA, and cgtB genes were 92.22%, 82.22%, 81.11%, and 22.22%, respectively. The findings revealed that the distribution of the majority of these genes were not dissimilar among Campylobacter species. The results emphasized that many of the pathogenic C. jejuni and C. coli may have these genes, and the Campylobacter strains with poultry origin have pathogenic potential properties for humans.
Introduction
Materials and Methods
Bacterial strains and culture method
A total of 90 Campylobacter strains including 48 C. jejuni and 42 C. coli were recovered from broiler feces during September 2011 to January 2012, in Shiraz slaughterhouse, Iran, according to the method that was previously described (Ansari-Lari et al., 2011). Briefly, fecal samples were collected in tryptic soy broth (TSB) broth tubes using sterile gloves and brought cooled to the laboratory in less than 6 h. To eliminate the other bacteria, a 0.8-μm membrane filter was used and 250 μL of filtered samples were cultured in an enriched TSB broth (containing 2.5 g/L dextrose, 0.5 g/L thioglycolate, and the selective supplement [10 mg/L rifampicin, 10 mg/L vancomycin, 10 mg/L ceftriaxone, 10 mg/L trimethoprim, and 20 mg/L amphotericin B] [Sigma, St. Louis, MO]), incubated in a microaerophilic atmosphere (Anaerocult C, Merck) at 37°C for 4 h, followed by incubation at 42°C for 44 h. Thereafter, 50 μL of enriched samples in TSB broth was cultured on selective agar (brucella agar base [41 g/L], and above antibiotics with identical dose). The growth of thermophilic campylobacters was detected by their typical appearance on culture media (i.e., the presence of flat grayish colonies like droplets of water sprayed on the medium). Preliminary identification of Campylobacter species was based on phenotypic characteristics, such as colony appearance, Gram staining, microscopic morphology, oxidase and catalase reaction, fermentation of glucose, and nitrate reduction. The type strains C. jejuni (ATCC 33291) and C. coli (RTCC 2541) were included as positive controls.
DNA preparation
DNA extraction was carried out using the phenol–chloroform extraction techniques. Briefly, a loopful of colonies of each isolates on an agar plate was picked and suspended in 200 μL of distilled water. After vortexing, the samples were centrifuged at 10,000×g; the supernatants were discarded before adding 250 μL of buffer 1 (resuspension solution contained 100 μg/mL RNase) and 250 μL of buffer 2 (Lysis buffer), a 550-μL saturated phenol was then added, mixed thoroughly, and centrifuged at 8000×g. The supernatant was collected into a new Eppendorf tube; the same volume of the phenol was then added and centrifuged as described above. The clear phase was collected into a new tube, before adding sodium acetate (2 M, pH 5.2, 0.1×volume of each aliquot). The aliquots were mixed with 1.5 mL 100% ethanol, kept at −20°C for 1 h, and centrifuged at 12,000×g. The supernatant was then discarded and a DNA pellet was washed with 80% ethanol, before being dried and suspended in 30 μL of 1x TE buffer (10 mM Tris-HCl pH 8, 1 mM EDTA) until further use.
PCR assay
Simple and multiplex PCR reactions were done for identification of Campylobacter genus and C. jejuni and C. coli species, respectively, and then PCR was done for detection of nine virulence factors listed in Table 1 for each confirmed strain. PCR amplifications were performed in a final volume of 25 μL. The reaction mixtures consisted of 2 μL of the DNA template, 2.5 μL 10x PCR buffer (75 mM Tris-HCl, pH 9.0, 2 mM MgCl2, 50 mM KCl, 20 mM [NH4]2SO4), (CinnaGen, Iran), 1 μL dNTPs (50 μM), (CinnaGen), 1 μL (1 U Ampli Taq DNA polymerase), (CinnaGen), 1 μL (25 pmol) from the forward and reverse primers (CinnaGen), shown in Tables 1 and 2, and the volumes of the reaction mixtures were reached to 25 μL using distilled deionized water. The thermal cycler (MJ mini, BioRad, USA) was adjusted under the following conditions: initial denaturation at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C for 1 min, and annealing as shown in Tables 1 and 2 for 1 min and extension at 72°C for 1 min. Final extension was carried out at 72°C for 10 min and the PCR products remained in the thermal cycler at 4°C until they were collected. Amplified products were separated by electrophoresis in 1.5% agarose gel stained with ethidium bromide. Visualization was undertaken using an ultraviolet transilluminator (BTS-20, Japan), and the 100-bp DNA ladders were used as molecular size markers (Figs. 1 and 2).
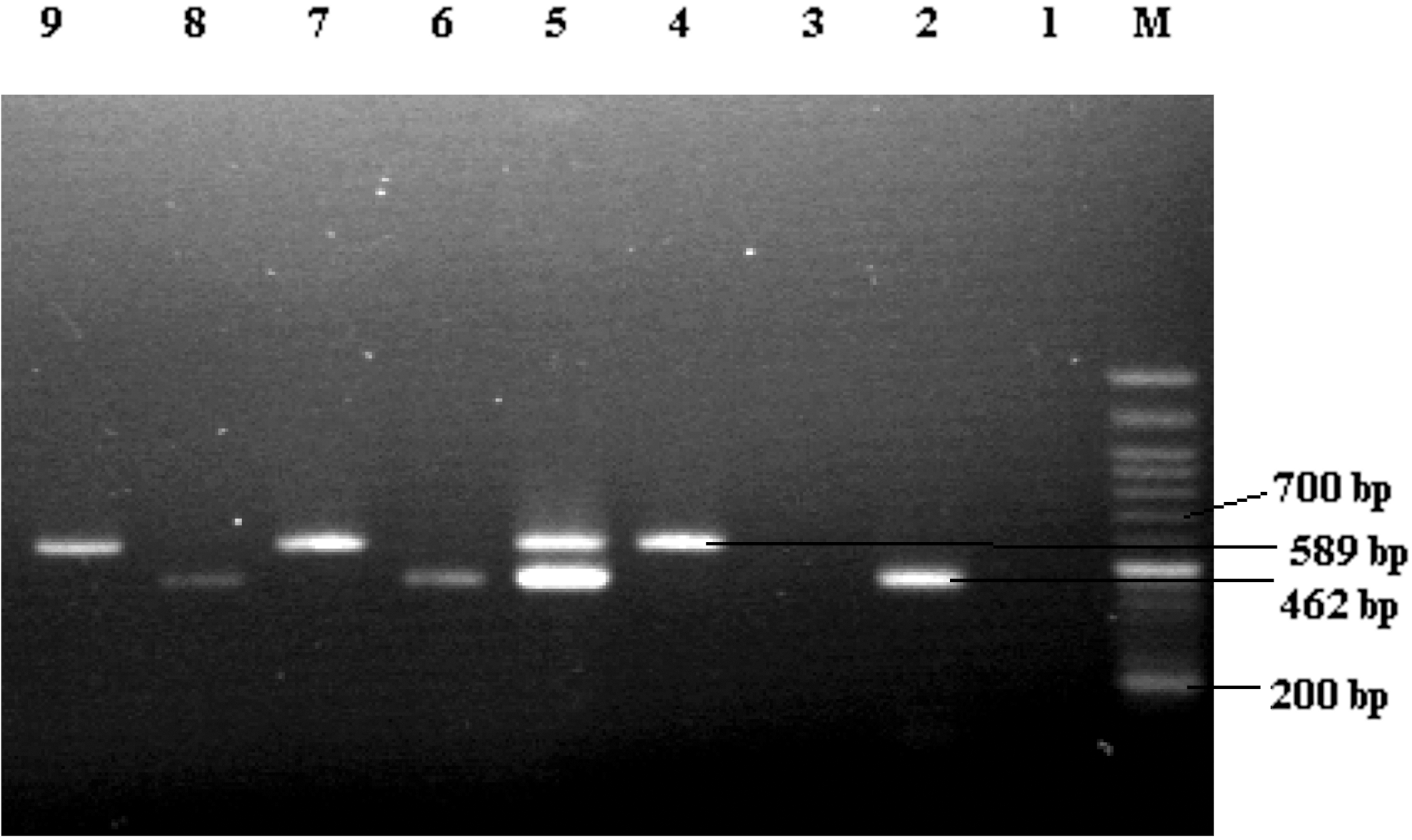
Agarose gel electrophoresis of mapA and ceuE gene multiplex polymerase chain reaction (PCR) products. Lane 1: negative control; Lane 3: negative sample; Lanes 2, 6, 8: ceuE gene PCR product; Lanes 4, 7, 9: mapA gene PCR product; Lane 6: Mix sample; Lane M: the 100-bp DNA marker.
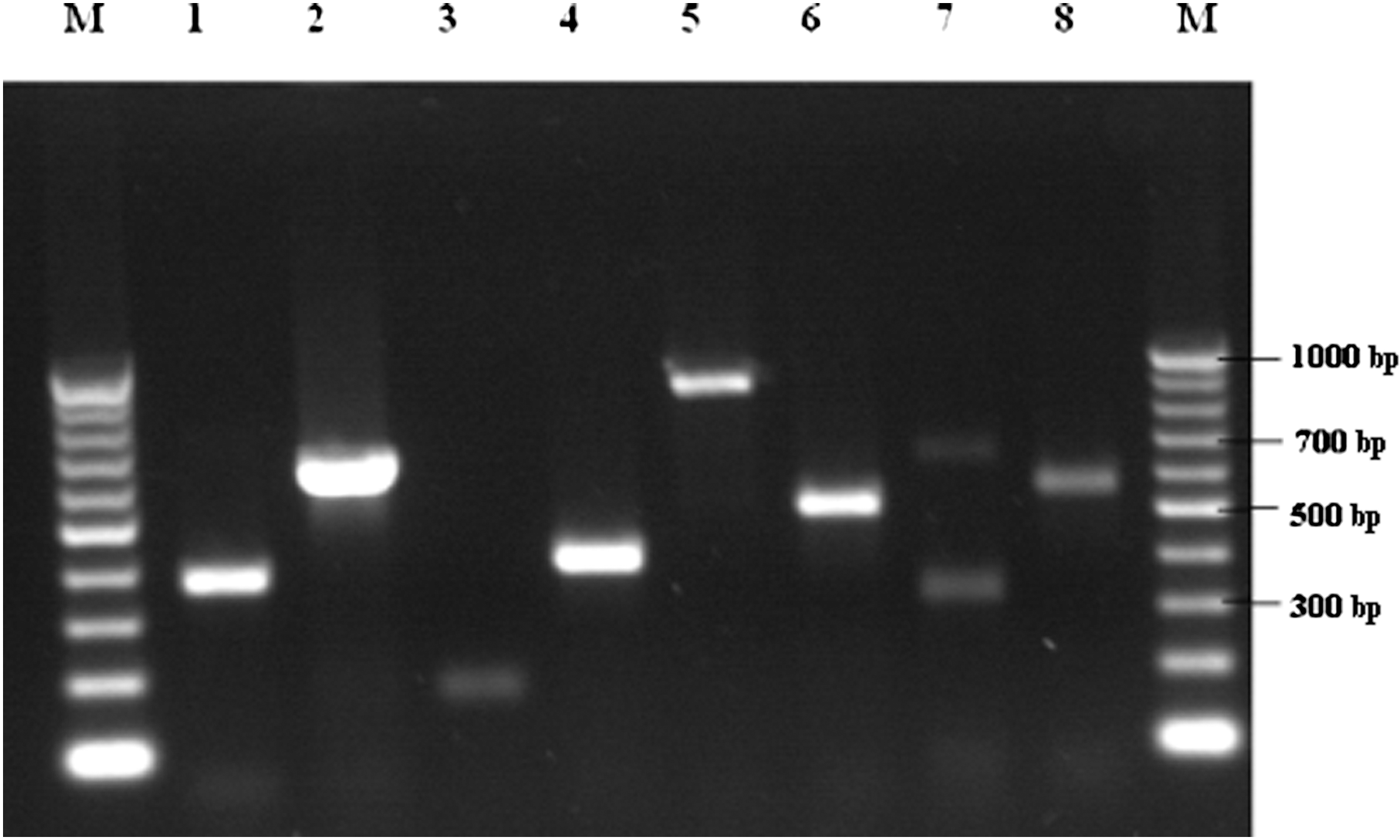
Agarose gel electrophoresis of polymerase chain reaction products of eight pathogenic genes of Campylobacter spp. Lanes: M, the 100-bp marker; 1, cdtA; 2, cdtB; 3, cdtC; 4, cadF; 5, pldA; 6, iamA; 7, wlaN; 8, cgtB.
Statistical analysis
Statistical analysis of the occurrence of the marker genes in the C. jejuni and C. coli isolates was performed using SPSS version 12.0.1. Discrete variables were expressed as percentages and proportions were compared using the chi-square test with the significance level defined at p<0.05.
Results
Among 90 Campylobacter strains including 48 C. jejuni and 42 C. coli, prevalence of cdtA, cdtB, cdtC, and cadF virulence genes was 100% (90/90). The plasmid-associated virulence marker, virB11 gene, was not present in any of the isolates. The frequency of the pldA gene among C. jejuni and C. coli strains was 48/48 (100%) and 35/42 (83.33%), respectively. Furthermore, in a total of 90 Campylobacter isolates, the presence of iamA, wlaN, and cgtB genes was 81.11%, 82.22%, and 22.22% respectively. Statistical analyses showed that the occurrence of the cgtB gene was significantly higher in C. jejuni isolates than in C. coli. The occurrence of the other virulence markers did not differ significantly between species. Detailed results of PCR detection of nine virulence genes in two thermophilic Campylobacter species obtained from poultry feces are shown in Table 3.
Discussion
Several factors have been suggested to clarify the varied clinical form associated with Campylobacter infection, and the phenotypic traits associated with different Campylobacter strains may be related to their genetic diversity (Wassenaar et al., 1998; Boer et al., 2002). Some studies emphasize the importance of poultry as a reservoir and source of Campylobacter infection (Corry and Atabay, 2001; Herman et al., 2003). Normally, Campylobacter colonize the intestines of chickens in the first week after hatching. Broiler flocks were contaminated with C. jejuni and C. coli during the growing cycle on the farms (Chuma et al., 1997). The research on the virulence properties of potentially pathogenic bacteria in domestic animals and in food with animal origin is essential to consumers' safety. The detection and occurrence of Campylobacter strains possessing pathogenicity markers in food have not yet been a subject of investigation in southern Iran, although it is an important factor of campylobacteriosis risk assessment. Investigation about the virulence marker properties of Campylobacter strains isolated from broilers, as the main reservoir in human campylobacteriosis, is significant. For this purpose, in this study, we investigate the distribution of nine virulence-associated genes of Campylobacter strains isolated from broiler feces. Results of the present study showed the high prevalence of seven virulence genes including three cdt genes (cadF, pldA, wlaN), and an invasion-associated gene, iamA; moreover, the virB11 gene with plasmid origin was absent among 90 Campylobacter isolates and an LOS-associated gene, cgtB, showed lower prevalence than the other seven virulence genes. The virulence of Campylobacter species is associated with the production of cytotoxins. The three cdt genes, which form a polycistronic cdt operon, all are necessary for the expression of cytotoxins that are known to be lethal for host enterocytes by arresting the G2 stage prior to mitosis (Smith and Bayles, 2006). Another investigation suggests that cytolethal distending toxin may also play a role in the development of the inflammatory process in humans (Hickey et al., 2000). Although according to the study of Pickett et al. (1996), C. jejuni shows higher cytotoxic activity in comparison to C. coli, in the current study both thermophilic species of Campylobacter showed 100% prevalence of cdt genes. More studies are necessary to determine the production of CDT toxin in these isolates. The rate of presence of three CDT-associated genes is higher than in some studies (Wieczorek and Osek, 2008; Rizal et al., 2010). However, other investigations showed 90%–100% prevalence of these genes among isolates from various sources (Müller et al., 2006; Datta et al., 2009; Ripabelli et al., 2010). Microarray testing of the cdt cluster showed that these genes were present in all C. jejuni isolates from human samples (Dorrell et al., 2001; Volokhov et al., 2003). Furthermore, Fouts et al. (2005) showed that cdt genes are conserved among different Campylobacter strains. The product of the cadF gene is an adhesin- and fibronectin-binding protein involved in the process of invasion, influencing microfilament organization in host cells (Monteville et al., 2003). It was shown that cadF-negative strains were not able to colonize chicken guts (Ziprin et al., 1999). Thus, the cadF gene, which appears to be essential for chicken gut colonization, may presumably have a similar role in the pathogenesis of human infection (Rozynek et al., 2005). Analysis of the prevalence of the cadF gene revealed that all of the Campylobacter isolates derived from poultry feces carried this marker, despite of their species. A study conducted by Rozynek et al. (2005) on Polish C. jejuni and C. coli isolates showed that all 92 analyzed strains obtained from poultry carcasses possessed the cadF gene, and 79 of 80 Campylobacter strains, which were recovered from children with diarrhea, also had this virulence marker. In a similar study, all isolates with human origin had the cadF gene (Rizal et al., 2010). The high prevalence (100%) of the cadF gene in the present study shows that many strains originating from poultry feces have pathogenic potential properties for humans. We assume that cadF genes are probably conserved among Campylobacter isolates. The distribution of the pldA gene, another important factor for colonization of C. jejuni in the intestine of chickens (Ziprin et al., 2001), was dissimilar among two species, as C. jejuni showed a higher presence of this gene. The results of our study are in accordance with the results of Datta et al. (2009), which showed enhancement in the presence of the pldA gene in chicken C. jejuni isolates from 88% to 100% with age, because our samples were recovered from broilers at the end of the breeding period in slaughterhouse. This phenomenon increases the chances of poultry serovars to have better colonization, making them more virulent for their other hosts. Carvalho et al. (2001) showed the presence of a 1.6-kb genetic marker of Campylobacter strains, isolated from children with diarrhea and symptom-free children, associated with adherence and invasion and named it as invasion-associated marker, but Al-Mahmeed et al. (2006) discussed the absence of this marker in the isolates from clinical cases in the children but not in older patients and suggested more research on the prevalence of this marker. In the present study, C. coli isolates showed higher prevalence of the iamA gene than C. jejuni strains. Rozynek et al. (2005) showed that only 1.6% of C. jejuni strains and 83.3% of the C. coli isolates from children possessed this sequence. Moreover, they showed that 50% of C. jejuni and 100% of C. coli isolates from chicken carcasses were iam positive. It seems that more investigations are necessary about the iam sequence product of different Campylobacter species, which probably is connected with the diarrheal form of the disease and their effects on human intestinal cells. Bacon et al. (2000) investigated the possible involvement of a plasmid in virulence of C. jejuni. A plasmid (pVir) contains four open reading frames and mutation in any of the four genes—the virB11 homologue gene—results in reduction in adherence and invasion. We could not detect a PCR product of the virB11 gene in any Campylobacter strains as in other similar studies (Müller et al., 2006; Feng et al., 2009). In some other studies, this gene was identified in 7%–15% of the tested isolates (Bang et al., 2003; Datta et al., 2003). These discrepancies may be due to variation in the isolates from a different geographic area and the nature of the virB11 plasmid. Some authors suggested that products of the pVir plasmid genes may have a significant role in serious cases of disease caused by C. jejuni (Tracz et al., 2005). We suggest that continuing investigation in different origins in this area needs to be performed in order to get more reliable data about the presence of the gene. The next virulence markers examined in the present study were the wlaN and the cgtB genes. The gene products of wlaN and cgtB both were characterized as ß-1, 3 galactosyltransferases. WlaN product is responsible for specific lipo-oligosaccharides (LOS) connected with long-term immunoreactive effects after C. jejuni infection such as Guillain-Barré and Miller–Fischer syndromes (Gilbert et al., 2000; Linton et al., 2000). In the present study, unlike other studies, the wlaN gene showed high prevalence among Campylobacter isolates (82.22%). Datta et al. (2003) showed the gene was present in 4.7% of broiler feces isolates and in less than 25% in other animal and human sources. Moreover, the wlaN gene was detected in 7.5% of C. jejuni strains isolated in Bangladesh from diarrheal patients (Talukder et al., 2008). Already, intragenic homopolymeric tract was detected within the wlaN gene (Parkhill et al., 2000). In this polymorphic region, variations in sequence were found. These variations resulted in changes of the open reading frame and in expression of intact or truncated proteins. Finally, the phase-variable expression of wlaN results in alternate ganglioside-mimicking LOS structures (Linton et al., 2000). Such differences as gene variations and presence or absence of the gene in isolates with different origins or various geographic areas can alter Campylobacter antigenic properties and change the ability of strains to infect their hosts. The presence of the cgtB gene in C. jejuni was about two times more than in C. coli. Results indicated that presumably the cgtB gene has a more significant role in C. jejuni LOS structure than C. coli strains. Altogether, 15 (16.66%) isolates showed the concurrent presence of the wlaN and cgtB genes, including 10 C. jejuni and five C. coli strains. A total of 90 Campylobacter strains isolated from broiler feces yielded only nine (10%) isolates, including six C. jejuni and three C. coli, showed the presence of all virulence genes except the virB11 gene.
Conclusions
In conclusion, the high prevalence of some virulence and toxin genes demonstrated that these putative pathogenic determinants are common among the Campylobacter isolates from poultry feces. The present study showed the high prevalence of the cadF and cdt genes among C. jejuni and C. coli isolated from poultry feces. Furthermore, the isolates showed a wide variation for the presence of some other pathogenic genes; however, the identification of a specific nucleotide sequence encoding virulence factor does not mean that the genes are expressed and their products are present. However, detection and better understanding of these factors allow us to recognize chromosome or plasmid-born virulence factors associated with diseases. These data also indicate that Campylobacter strains recovered from broiler feces may have a potential virulence for humans. Finally, additional studies of the prevalence of virulence genes in Campylobacter isolates from various sources are essential to get more information on Campylobacter spp. hazards circulating in developing countries like Iran.
Footnotes
Acknowledgments
This work was supported by a Grant from Shiraz University.
Disclosure Statement
No competing financial interests exist.