
Abstract
Enterohemorrhagic Escherichia coli (EHEC) causes a wide spectrum of food- and waterborne infectious diseases, including diarrhea, hemorrhagic colitis, and even hemolytic–uremic syndrome. Porcine attaching and effacing–associated protein (Paa) was first identified in a porcine enteropathogenic E. coli strain. It has been proven essential in the attaching and effacing mechanism of EHEC. However, the immunologic function of the Paa protein has yet to be established. In the present study, recombinant Paa protein was overexpressed successfully in engineered E. coli and effectively purified to homogeneity. Comparative experiments were carried out in mice with a known adhesion factor (intimin) as reference to investigate the immunogenicity of Paa. Intraperitoneal immunization of Paa protein in mice elicited significantly high levels of serum immunoglobulin G antibodies via Th2-mediated humoral immune response. In mice challenged with E. coli O157:H7, Paa protein exhibited immunological effectiveness against pathogenic bacteria colonization and excretion in vivo. Compared with the intimin, Paa showed better protective effect against E. coli O157:H7 infection in mice, particularly those challenged with high lethal doses of the pathogen. Seventy percent of the mice challenged with 50 minimal lethal dose (MLD) in the Paa group survived, whereas only 50% survived in the intimin group. This finding is the first description of the immunologic function of the Paa protein. These attributes provide support for the development of Paa-based vaccine, which can be beneficial in treating infectious diseases caused by E. coli O157:H7.
Introduction
The pathogenic mechanism of the E. coli O157:H7 infection begins with the attachment or adhesion of the bacteria to the host. The adhesion is mediated by a type III secretion system and a number of bacteria-secreted protein interactions (Elliott et al., 1998), with intimin serving as a key protein in the system (DeVinney et al., 1999). Numerous studies have proven that intimin is an effective vaccine candidate because of its good immunogenicity and efficient protection (Gansheroff et al., 1999; Judge et al., 2004). Studies on different intimins from enteropathogenic E. coli (EPEC) and EHEC have shown that receptor-binding activity is localized in the C-terminal 280 amino acids (Int280), which mediate the interaction with Tir. In our previous work, recombinant protein Int281 (a fragment of intimin) showed excellent immunogenicity, and its specific immunoglobulin G (IgG) antibody titers reached 106 (Gao et al., 2008).
Porcine attaching and effacing associated–protein (Paa) was first identified in a porcine EPEC (PEPEC) strain (An et al., 1999; Batisson et al., 2003). The 753-bp Paa gene encoded a 27.6-kDa protein and was localized in the chromosome. In vivo, Paa-negative mutants induced less severe or no attaching and effacing (A/E) lesions in piglets, which did not develop diarrhea or delayed-onset diarrhea. Moreover, the adherent phenotype was restored after the complementation of the Paa mutant. Further tests revealed that Paa plays a role similar to that of the adherence and/or colonization factor in enterotoxigenic E. coli (ETEC) or EHEC infection. In addition, a study on the distribution of Paa in various groups of diarrheagenic E. coli revealed the presence of Paa in all tested O157:H7 strains. The sequences of the Paa protein in the O157:H7 strains EDL933 and Sakai were found to be identical to that from the original pig EPEC strain (Wadolkowski et al., 1990). These findings suggest that Paa could play a specific role in the pathogenesis of EHEC. Vaccination with bacterial adherence and/or colonization factors, such as vaccination with intimin as described previously, has been suggested as a strategy to prevent EHEC infection (Gao et al., 2008). Therefore, the capability of Paa to provide protection against EHEC infection is worth exploring. The present study aims to express the Paa protein and to detect its immunogenicity and immunoprotective properties compared with the intimin fragment, both in vitro and in vivo. The results of the study are directed toward producing a promising new vaccine candidate that can provide enhanced protection against EHEC infection.
Materials and Methods
Bacterial strains, plasmids, and media
The bacterial strains and plasmids used in this study are listed in Table 1. Bacteria were routinely grown in Luria–Bertani (LB) broth supplemented with 100 μg/mL ampicillin as needed to select and maintain the plasmid. LB broth without ampicillin was used to culture O157:H7. Human HeLa cells for the adherence assay were grown in Dulbecco's modified Eagle's medium (RPMI-1640, Gibco) at 37°C.
Expression and purification
The Paa gene was obtained from the genome of E. coli O157:H7 EDL933, ligated into pET-22b(+), and transformed into the E. coli BL21(DE3). Overnight culture of E. coli BL21(DE3) containing the Paa was diluted in 1 L LB broth at a ratio of 1:100. When cells were grown to OD600=0.6, 1 mM isopropyl β-
The supernatant was used for purification following the instructions for HisTrap™ FF Resin Chromatography (GE Healthcare). The purified protein was validated by 15% SDS-PAGE.
Mice immunization and challenge
Thirty female BALB/c mice (Vivarium of Academy of Military Medical Sciences, Beijing, China) weighing 14–16 g were randomly divided into three groups. The three groups were immunized intraperitoneally (ip) with (1) Paa, (2) Int281, and (3) bovine serum albumin (BSA). The final concentration of each protein per dose for primary immunization was 25 μg; a dose of 50 μg was used for boosting. All samples were immersed in PBS and emulsified with the same volume as Freund's complete adjuvant; Freund's incomplete adjuvant was used for boosting every 2 weeks.
The mice used for the challenge test were fed with 5 g/L streptomycin for 3 days and were deprived of food 24 h before the challenge. The mice were boosted either twice or thrice. On day 14 after the last booster, the thrice-immunized mice were challenged intragastrically with 109 colony-forming units (CFU; 10 minimal lethal dose [MLD]) of E. coli O157:H7 88321 strain. The twice-immunized mice were challenged intragastrically with 5×109 CFU (50 MLD) or 1010 CFU (100 MLD) of E. coli O157:H7 88321 strain.
Immune parameters
ELISA titers
The mice were immunized on days 0, 14, and 28; sera were collected on days 0, 10, 24, 46, and 82. Antibodies specific to Paa, BSA, and Int281 were measured by enzyme-linked immunosorbent assay (ELISA). Mouse sera (after the last boost) serially diluted in PBS and goat anti-mouse IgG–horseradish peroxidase (HRP) (Sigma) were used as the primary antibodies and second antibodies, respectively. The reactions were developed with tetramethyl benzidine (TMB) and stopped with 2 M H2SO4. The absorbance at 450 nm was detected finally.
Subtype analysis of sera antibodies
The same ELISA method was employed to determine the titers of the specific IgG of the sera. Purified Paa (0.1 mg/mL) was used as antigen, and 100 μL of sera serially diluted in PBS (pH=7.4) were added, followed by incubation for 4 h at 4°C. The detected antibodies were goat anti-mouse IgA-HRP, goat anti-mouse IgG1-HRP, goat anti-mouse IgG2a-HRP, goat anti-mouse Ig2b-HRP, and goat anti-mouse IgG3-HRP (Sigma).
EHEC colonization model
After the challenge, the animals were individually housed and given access to food and water containing streptomycin (5 g/L). Fecal samples from each mouse were collected following infection over the following 2 weeks at 2-day intervals. E. coli O157:H7 fecal shedding was monitored by adding approximately 0.2 g to 1 mL of LB broth, followed by incubation at 4°C for 2–4 h to allow the fecal pellets to soften. The mixture was then vortexed until the pellets were no longer visible. Serial dilutions of the supernatant were plated onto MacConkey agar plates containing novobiocin, tellurite, kanamycin, streptomycin, and vancomycin (Wadolkowski et al., 1990). Plates were incubated overnight at 37°C, and E. coli O157:H7 colonies were enumerated the following day. The limit of detection for plating was 100 CFUs per 0.1 g feces. Bacterial colonies were tested for O157 antigen by polymerase chain reaction (Gao et al., 2011).
Bacterial adhesion
To analyze the anti-adhesion effect of the immune sera, HeLa cells were added (104/well) and grown overnight in 96-well plates with RPMI-1640 supplemented with 10% (vol/vol) fetal bovine serum at 37°C. E. coli O157:H7 EDL933 cells (105/well) were incubated with serially diluted immune sera of Paa-immunized mice or Int281-immunized mice for 1 h at 37°C. All samples were then added and incubated with monolayer HeLa cells for another 4 h at 37°C. The supernatant was discarded, and the cells were washed five times with PBS. Then, 0.25% pancreatin was used to digest the cells. The suspended bacteria were serially diluted 10-fold, plated onto MacConkey agar, and incubated at 37°C for 16 h. Subsequently, the CFUs of bacteria that adhered to the HeLa cells were counted. The sera of the BSA-immunized mice were used as control.
Statistics
All data were processed and analyzed using SPSS 16.0 Data Editor. Differences between groups were considered significant if p<0.05. Significant differences in mice survival between different immunization groups were determined by one-sided Fisher's exact test.
Results
Preparation and characterization of Paa
Paa was expressed in E. coli BL21 (DE3) and purified by HisTrap™ FF. It was then identified by SDS-PAGE and Western blot (Fig. 1). The final yield of the bioactive Paa (purity >95%) was approximately 35 mg/L.
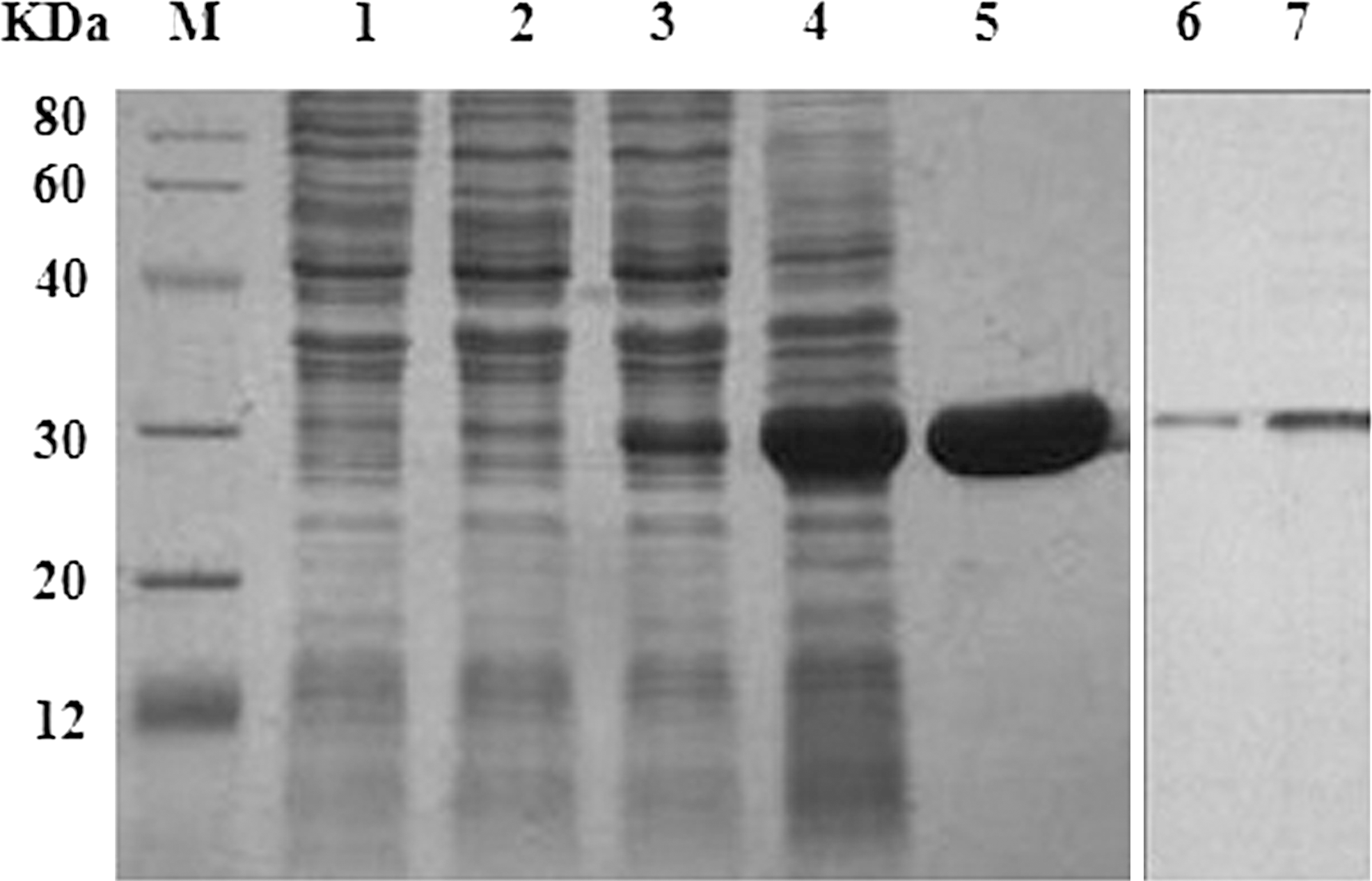
Preparation and characterization analysis of porcine attaching and effacing associated–protein (Paa). M, protein marker; Lane 1, total bacterial proteins of Escherichia coli O157:H7 EDL933; Lane 2, the proteins of E. coli BL21 (DE3); Lane 3, the proteins of E. coli BL21 (DE3) containing Paa un-induced by isopropyl β-D-1-thiogalactopyranoside (IPTG); Lane 4, the proteins of E. coli BL21 (DE3) containing Paa induced by IPTG; Lane 5, the purified recombinant Paa protein; Lane 6, Paa recognized by anti-His antibody; Lane 7, Paa recognized by anti-O157:H7 sera.
Time course of the antibody titers during immunization
To evaluate immunization response, specific IgG antibody titers changing during immunization were monitored by ELISA as previously described. Then, the generation of antibodies in mice with Int281 was compared with that in the BSA group as control. The results are shown in Figure 2.
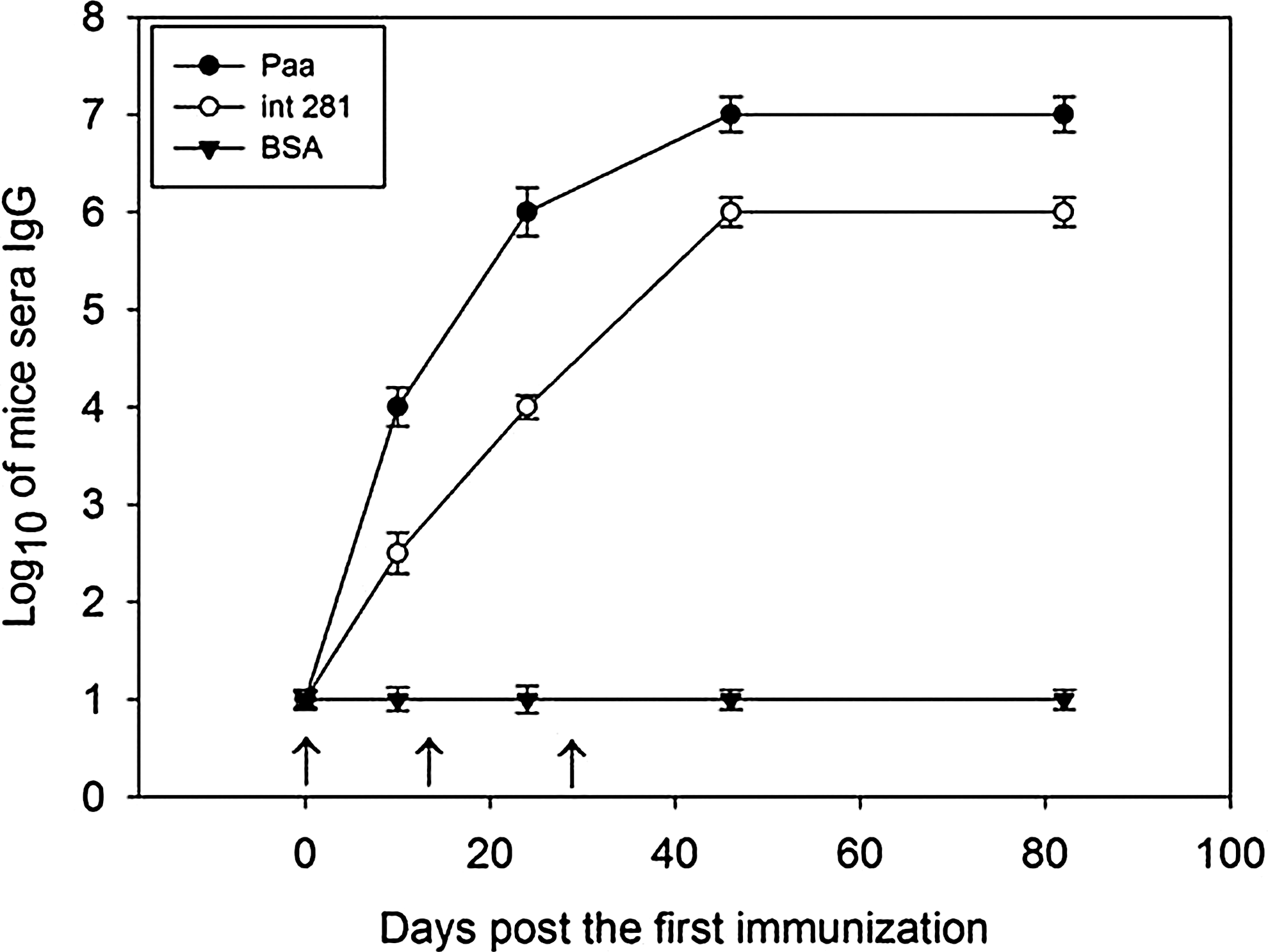
Time course analysis of the porcine attaching and effacing associated–protein (Paa)-specific immunoglobulin (IgG) titers. A total of 30 mice were randomly divided into three groups. Detection was performed by enzyme-linked immunosorbent assay. Purified (Paa), Int281 (fragment of intimin), and bovine serum albumin (BSA) (100 μg/mL) were coated on 96-well plates. Goat anti-mouse immunoglobulin G (IgG)–horseradish peroxidase (1:5000) was used as the antibody. The horizontal bars represent the geometric mean of the log of the IgG titers to Paa, fragment of intimin (Int281), and BSA; the error bars indicate±1 S.E, and the arrows represent the immunized points.
The ELISA titers of Paa peaked on day 14 after the third vaccination and did not decrease until day 82. During immunization, the IgG titers specific to Paa and Int281 were higher than those specific to BSA (p<0.01). Meanwhile, the IgG antibody titer of the Paa immunization-induced serum was significantly higher than that of the Int281 immunization-induced serum (p<0.01) during monitoring.
Subtyping of IgG induced by Paa
The titers of the IgG subtypes (IgG1, IgG2a, IgG2b, and IgG3) in the final vaccinated mice sera were examined by ELISA to determine which type dominates the response (Fig. 3).
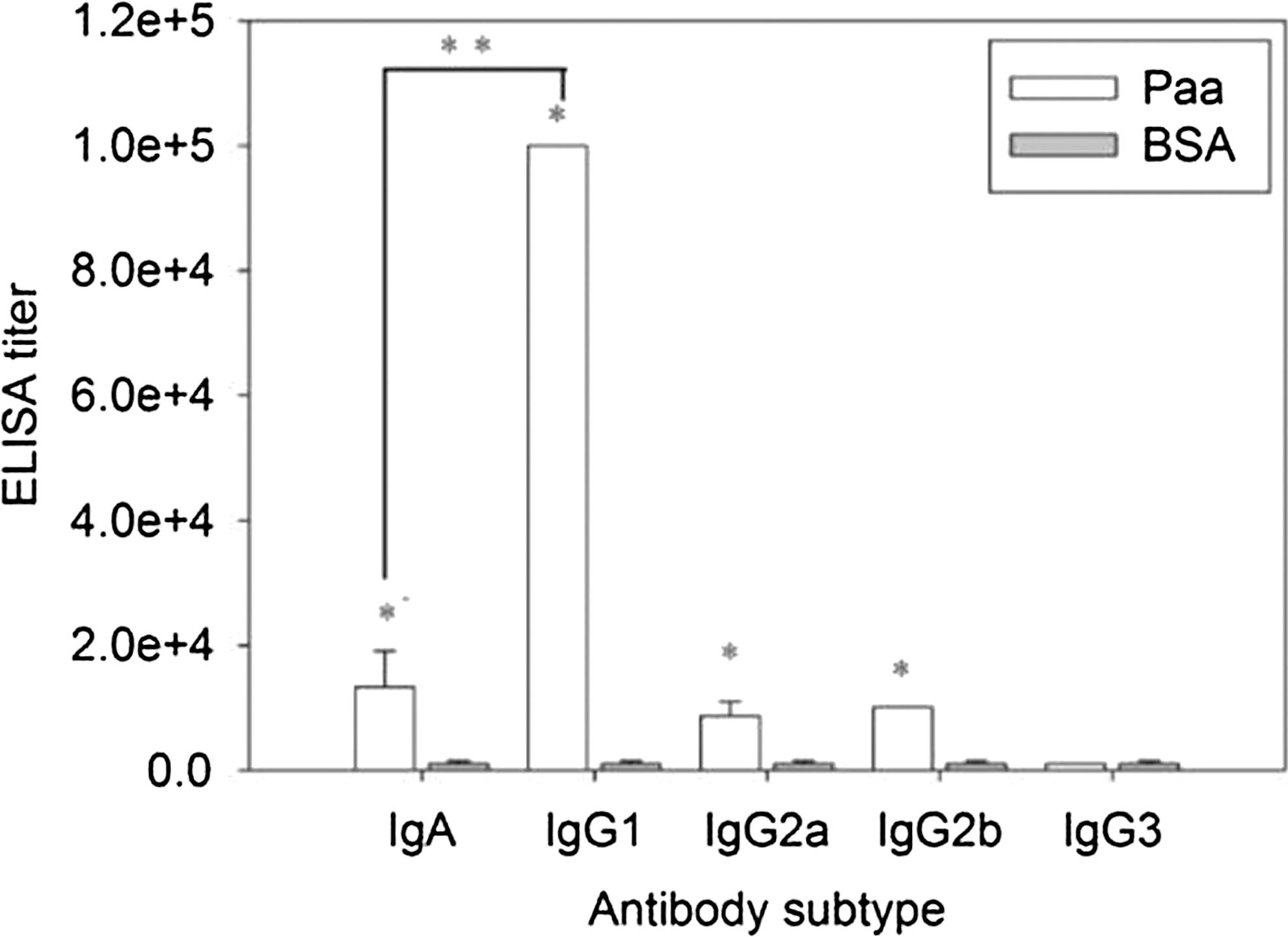
Serum-specific immunoglobulin G (IgG) subtype titers and IgA titers from porcine attaching and effacing associated–protein (Paa)-immunized mice. Enzyme-linked immunosorbent assay (ELISA) was used to perform the detection. Purified Paa (50 μg/mL) was used as antigen. Goat anti-mouse immunoglobulin A–horseradish peroxidase (IgA-HRP), goat anti-mouse IgG1-HRP, goat anti-mouse IgG2a-HRP, goat anti-mouse IgG2b-HRP, and goat anti-mouse IgG3-HRP (1:5000) were used as the antibodies. *p<0.05 versus bovine serum albumin (BSA) group in each subtype; **p<0.01.
IgA, IgG1, IgG2a, and IgG2b of the Paa group were significantly higher than those of the BSA group (p<0.01); IgG3 did not exhibit any further increase. IgG1 was significantly higher than the other subtypes in the Paa group (p<0.01). IgG1 is known to be associated with Th2-mediated humoral immune response. Thus, the immune response to Paa in the study was assumed to be biased to Th2-mediated humoral immune response.
Bacterial adhesion to epithelial cells
The CFUs of adherent E. coli O157:H7 cells were counted to analyze the anti-adhesion effect of the immune sera. The Paa immune sera could inhibit 90% bacterial adhesion onto target cells. This value was significantly higher than that of the Int281 sera (79.79%, p<0.05), as shown in Figure 4. The protection rate decreased following the dilution of sera. When the sera were diluted 128 times, the Paa immune sera could protect 9% of the cells (p<0.01), whereas the Int281 immune sera failed to inhibit adhesion. The Paa sera failed to inhibit adhesion when diluted 256 times. The protection rates of the Paa were higher than that of Int281 for all cases.
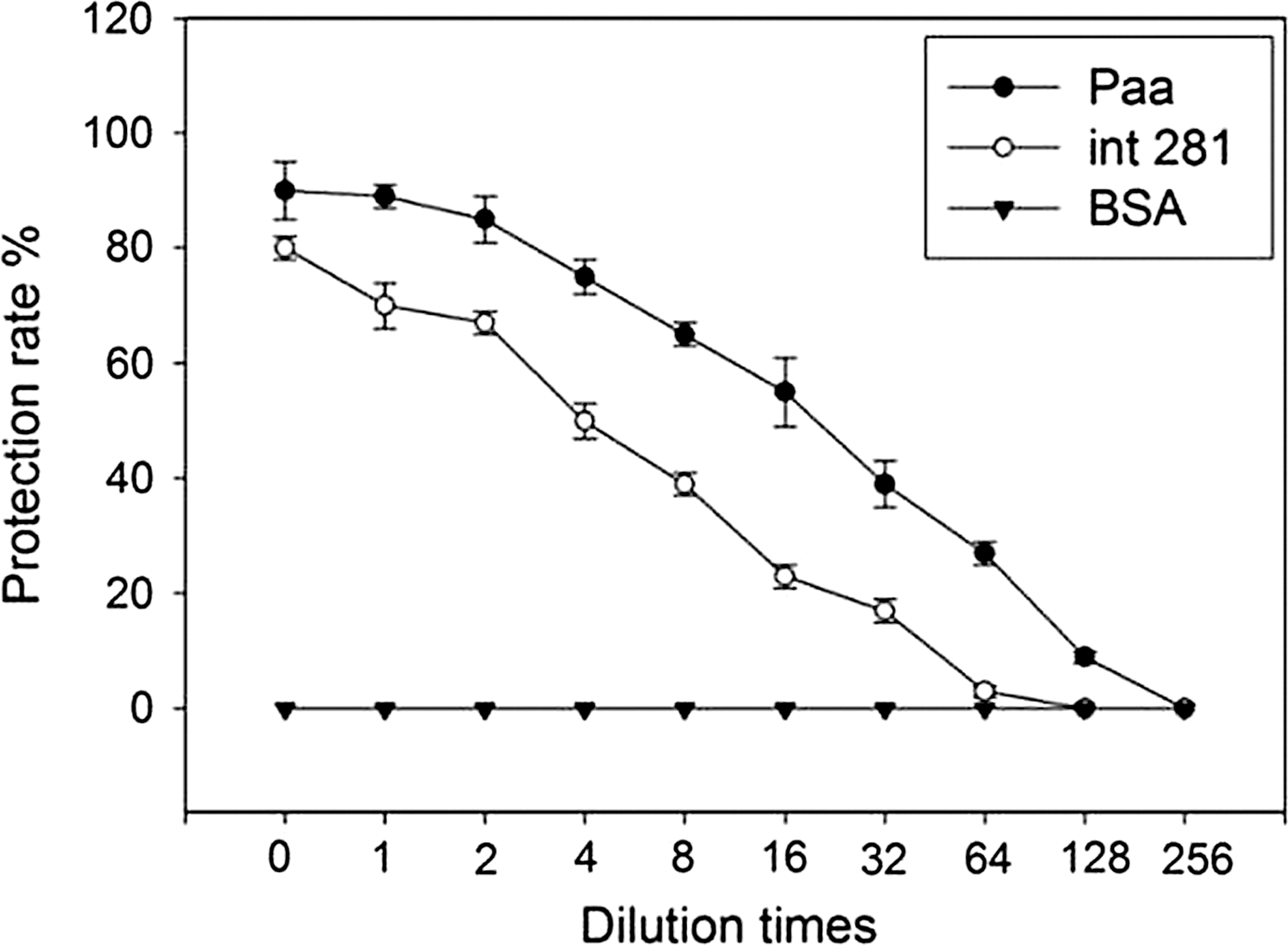
The protection of anti-adhesion effect provided by porcine attaching and effacing associated–protein (Paa). Escherichia coli O157:H7 EDL933 cells (105/well) were incubated with serially diluted immune sera of Paa-immunized mice or Int-immunized mice for 1 h at 37°C. All samples were then added and incubated with monolayer HeLa cells for another 4 h at 37°C. After digestion, the suspended bacteria were serially diluted 10-fold, plated on MacConkey agar, and incubated at 37°C for 16 h. After that, the colony-forming units of bacteria were counted. BSA, bovine serum albumin; int281, (fragment of intimin).
In vivo colonization
Two weeks following immunization, the CFU of E. coli O157:H7 shedding in feces was counted. The results of the fecal shedding are given in Figure 5. During the entire sampling period, the BSA-immunized control mice shed higher levels (p<0.01) of E. coli O157:H7 than the Paa- or Int281-immunized mice. On day 2 after infection, the Int281-immunized mice shed significantly higher (p<0.01) E. coli O157:H7 than the Paa-immunized mice. None of the mice immunized with either Paa or Int281 shed the organism after day 10 of the challenge, whereas all BSA-immunized control mice shed high levels of E. coli O157:H7 over the entire sampling period. Immunization with Paa or Int281 recombinant proteins obviously reduced EHEC colonization in mice intestines, particularly those immunized with Paa.

Escherichia coli O157:H7 88321 fecal shedding following intragastric administration in mice. The mice were immunized with porcine attaching and effacing associated–protein (Paa), Int281 (fragment of intimin), and bovine serum albumin
Protection of immunized mice against O157:H7 88321
The three-times-immunized mice
Two weeks after the last booster, all immunized mice were challenged intragastrically with 109 CFU of E. coli O157:H7 88321 (10 MLD). Among all mice immunized with BSA/adjuvant, three from the Int281group and one from the Paa group displayed slight diarrhea characterized by soft and sodden feces followed by an infection on the second day. Ten days postchallenge, all mice from the Paa and Int281 groups survived, whereas all BSA-immunized mice died (0/10) (Fig. 6A).
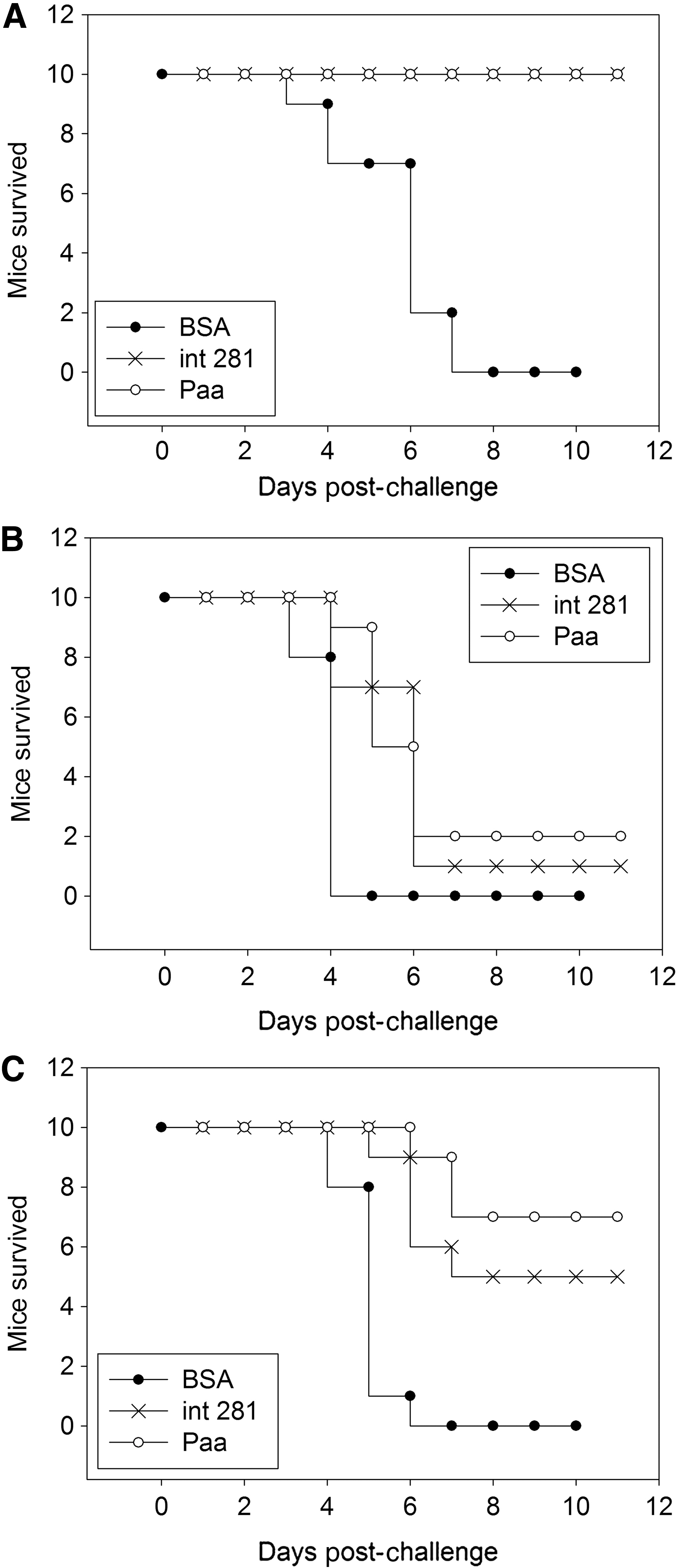
Protection of porcine attaching and effacing associated–protein (Paa)-immunized mice against a high lethal dose of Escherichia coli O157:H7. Each experimental group comprised 10 mice. The experiments were repeated three times with different doses of enterohemorrhagic Escherichia coli O157:H7.
The twice-immunized mice
The twice-immunized mice were challenged with two lethal doses of the pathogen: 5×109 CFU (50 MLD) and 1010 CFU (100 MLD). Seventy percent of the mice (7/10) challenged with 50 MLD in the Paa group survived, whereas only 50% of the mice (5/10) survived in the Int281 group (Fig. 6B). Meanwhile, only 20% of the mice (2/10) challenged with 100 MLD in the Paa group survived, whereas 10% of the mice (1/10) survived in the Int281 group (Fig. 6C). All BSA-immunized (negative control) mice died (0/0).
Discussion
EHEC infection is highly pathogenic and fatal (Mukherjee et al., 2002). Currently, the most promising approach to the prevention of EHEC infection is the development of a genetic vaccine based on candidate antigens.
In this study, the recombinant Paa protein of EHEC O157 was successfully constructed and expressed in soluble form with high yield. The expressed Paa demonstrated strong immunogenicity. The mice immunized three times with Paa were challenged with 10 MLD E. coli O157:H7. All mice in the Paa group survived, whereas all mice in the BSA group died. Thus, in the next experiment, we decreased the immunization frequency and added the doses of bacteria used in the challenge assays to determine the exact protective effect of the Paa protein. The twice-immunized mice were challenged with two lethal doses of E. coli O157:H7 88321. Paa provided enhanced protection during the challenge with 50 MLD or 100 MLD. Paa also induced an antibody response with high titers and maintained higher IgG titers than the BSA during the challenge. The dominant immunoglobulin subclass in mice sera was analyzed to understand further the response mechanism of Paa immunization. The level of IgA was found to be lower than that of IgG after immunization. This result indicates that mucosal immunity is not the main immune response. Among the IgG subclasses of mice sera, IgG1 was found to be dominant. Similar to a number of subunit vaccines, such as Int281 mentioned in our previous study (Cai et al., 2010), Paa significantly induced the Th2-mediated humoral immune response.
The capability of the Paa protein to protect against E. coli O157:H7 colonization and excretion in vivo and in vitro was also evaluated. In vivo, E. coli O157:H7 fecal shedding was detected after intragastric administration of Paa. The limit of detection for plating was 100 CFUs per 0.1 g feces. Interestingly, the vaccine formulation in the study restricted fecal shedding in all vaccinated animals to less than 102 CFU/0.1 g feces at day 10 after the infection. Meanwhile, the BSA-vaccinated animals still exhibited fecal shedding that ranged between 105 and 106 CFU/0.1 g feces. The results suggest that the candidate antigen vaccine may have a significant impact on the spread of bacteria in a group of animals. Along with the decreased excretion was the low colonization of E. coli O157:H7 in the intestinal tract of the vaccinated animals, excluding those immunized with BSA. In vitro, Paa could inhibit bacterial adhesion onto HeLa cells, with an inhibition rate (90%) higher than that of BSA (0%) and even higher than that of some fusion proteins, such as a fusion protein with stx2B-stx1B-int281 (SSI) (83.33%) reported in our previous study (Gao et al., 2011). The cytotoxicity of Paa was also analyzed in this study. Analysis results reveal that a super high concentration (1 mg/mL) of Paa protein had no toxicity effect on HeLa cells (data not shown).
The intimin fragment (Int281) was compared with Paa in the study. Int281 showed excellent immunogenicity and high IgG antibody titers (106). However, Paa demonstrated higher specific IgG titers (107) compared with Int281 when mice were challenged with 50 MLD and 100 MLD E. coli O157:H7 88321. Paa also presented better protective immunity than Int281. Furthermore, Paa could provide a more effective protection against E. coli O157:H7 colonization and excretion in vivo and in vitro than Int281.
The Paa gene is often identified in the genome of pathogenic E. coli, such as ETEC, EPEC, and EHEC. The Paa gene has been occasionally detected in ETEC strains that have been isolated as early as 1978 (3% of the isolates), and it has been observed in an increasing proportion of strains isolated in recent years (75% of E. coli isolated) (Maynard et al., 2003; Boerlin et al., 2005). The presence of Paa was associated with the eae gene and its A/E phenotype in PEPEC O45 strains (Batisson et al., 2003). In the examination of ETEC isolates from humans and various animal species, a strong correlation was observed between the presence of Paa and that of eae in EHEC O157:H7 (An et al., 1999). In the current study, the nucleotide sequences of Paa from typical pathogenic serotypes of EHEC, including O157, O55, O26, O111, and O103, were blasted and analyzed. Paa was found to have the highest homology (above 99%) among the different EHEC strains. As previously described, the sequence of the Paa protein in EHEC O157:H7 was similar to that of the original PEPEC strain (100%) and to that of the eight ETEC strains (above 92%) (Batisson et al., 2003). These findings indicate high conservation of protein sequences of Paa in virulent E. coli strains. Therefore, the Paa-based candidates reported in this study may theoretically provide an effective protection against EPEC and ETEC infection.
This study is the first to describe an efficient preparation of Paa protein by genetic engineering technology. The results demonstrate that Paa can significantly reduce bacterial shedding following a challenge with E. coli O157:H7 and provide better protection against E. coli O157:H7 infection in mice compared with the intimin fragment. The data from this study prove that Paa is an effective and attractive novel vaccine candidate.
Footnotes
Acknowledgments
This work is supported by National Natural Science Foundation of China (No. 81072677) and Beijing Natural Science Foundation of China (No. 7122134).
Disclosure Statement
No competing financial interests exist.