
Abstract
The antimicrobial activity and model of action of polyphenolic compounds extracted from lowbush wild blueberries (LWB) were studied against Escherichia coli O157:H7. Polyphenols in LWB were extracted using 80% vol/vol methanol and designated as total blueberry phenolics (TBP). The fraction was further separated by a C-18 Sep-Pak cartridge into monomeric phenolics acids (MPA) and anthocyanins plus proanthocyanidins (A&P). The A&P fraction was further separated into anthocyanins and proanthocyanidins using a LH-20 Sephadex column. Each fraction was diluted in 0.85% wt/vol NaCl, inoculated with E. coli O157:H7 to achieve 8 log colony-forming units (CFU)/mL, and incubated at 25°C for 1 h. The survival populations of E. coli O157:H7 in the phenolic fractions were determined by a viable cell counts method. The permeability of the cell membrane of E. coli O157:H7 was determined using LIVE/DEAD viability assay, and the damage was visualized by using transmission electron microscopy (TEM). Significant (p<0.05) reductions of 5 log CFU/mL of E. coli O157:H7 were observed for MPA at 0.4 g/L gallic acid equivalents (GAE), A&P at 0.9 g/L GAE, and anthocyanins at 0.65 g/L GAE. Reductions of 6–7 CFU/mL were observed for MPA at 0.8 g/L GAE, A&P at 1.8 g/L GAE, and anthocyanins at 1.3 g/L GAE compared to the control. The cell membrane of E. coli O157:H7 exhibited a significantly increased permeability when treated with proanthocyanidins (0.15 g/L GAE), A&P (0.45 g/L GAE), anthocyanins (0.65 g/L GAE), and TBP (0.14 g/L GAE). TEM confirmed the inactivation and increased membrane permeability of E. coli O157:H7. This study demonstrated the antimicrobial effect of polyphenols from LWB against E. coli O157:H7 and the probable mode of action.
Introduction
Multiple studies have demonstrated the antimicrobial properties of phytochemicals isolated from Vaccinium berries against both Gram-positive and Gram-negative foodborne pathogens (Puupponen-Pimiä et al., 2001; Viskelis et al., 2009; Biswas et al., 2012). Recent studies indicated that 100% highbush blueberry (V. corymbosum) juice inhibited the growth of Salmonella Typhimurium, Escherichia coli O157:H7, and Campylobacter jejuni without affecting the growth of probiotic species such as Bifidobacteria bifidum and Lactobacillus bulgaris (Biswas et al., 2012). In addition, Puupponen-Pimiä et al. (2001) demonstrated that bilberries (Vaccinium myrtillus), the European variety of blueberries, inhibited the growth of Salmonella Typhimurium and E. coli CM 871, but had no effect on the growth of Lactobacillus acidophilus and Bifidobacterium lactis. Research examining the phytochemical constituents of LWB demonstrated that E. coli O157:H7, Salmonella Typhimurium, and Listeria monocytogenes were twice as susceptible as Lactobacillus rhamnosus to the growth-inhibitory effect of LWB monomeric phenolic acids, anthocyanins, and proanthocyanidins (Lacombe et al., 2012). These findings indicated an important advantage of using LWB in foods to control the growth of pathogenic microorganisms while maintaining the viability of beneficial microorganisms.
Natural products are being revisited as potential sources of antimicrobials. This burgeoning market for natural antimicrobials requires reliable information regarding their safety, economy, ecological impacts, and efficacy (Roller and Lusengo, 1997). With continued research efforts, antimicrobials derived from Vaccinium berries may become an alternative to synthetic preservatives and antibiotics. Results from previous studies indicated that berries have the potential to be used as functional ingredients as well as natural antimicrobials. The objective of this study was to investigate the effect of phenolic compounds derived from LWB on the growth of E. coli O157:H7 and on the permeability and integrity of the bacterium's cell membrane. Findings from this study would provide insight into the effects of LWB phenolic compounds on the growth behavior and the membrane integrity of E. coli O157:H7 and elucidate the associated mode of action.
Materials and Methods
Preparation and quantification of LWB phenolic fractions
Freeze-dried LWB powder (Durham Research, Londonderry, NH), 40 g, in 100 mL 80% vol/vol methanol was stirred overnight at 25°C according to the procedure described by Lacombe et al. (2012). From the solution, 4 mL was removed and designated as total blueberry phenolics (TBP). TBP was separated into monomeric phenolic acids (MPA) and anthocyanins plus proanthocyanidins (A&P) using a 35-cc SePak C-18 cartridge (Waters, Milford, MA) according to the procedures of Lacombe et al. (2012). A LH-20 Sephadex column (GE Healthcare, Schenectady, NY) was used to further separate A&P into anthocyanins and proanthocyanidins according to the procedure of Kalt et al. (2008). After rotorevaporation and overnight drying in a desiccator, all fractions were resuspended in distilled water and stored at 5°C in the dark for use in the experiments.
The quantification of all fractions was conducted after drying and reconstitution. The Folin-Ciocalteu method (Slinkard and Singleton, 1977) was used to analyze the total phenolics in each fraction. Samples were read at 725 nm in a 96-well microplate (Biotech, Winoskii, VT), and the total phenolics concentrations were reported as the equivalents of gallic acid (GAE). A pH differential method was used to determine the total anthocyanins concentration, based on structural changes of the anthocyanins chromophore between pH 1.0 and 4.5 (Lee et al., 2008). The difference in absorbance was used as indication of relative anthocyanins content, and concentrations were reported as the equivalents of cyanidin 3-galactoside (Lee et al., 2008). The 4-dimethylaminocinnamaldehyde (DMAC) protocol (Payne et al., 2010) was used to quantify proanthocyanidins. The absorbance of the reaction product between the proanthocyanidins in the sample and DMAC reagent (Sigma Aldrich, St. Louis, MO) was measured at 640 nm in a 96-well microplate. Results were compared to a standard curve of procyanidin-A2/DMAC (Indofine, Hillsborough, NJ) and reported as the equivalents of procyanidin-A2 (PA2).
Culture preparation
E. coli O157:H7 ATCC 35150 and 12900 (The American Type Culture Collection, Manassas, VA) were grown in brain heart infusion (BHI) broth (Neogen, Lansing, MI) at 37°C for 24 h. To ensure purity of the cultures, strains were grown on MacConkey Sorbitol agar (MSA; Neogen) and sorbitol fermenting colonies were verified with the LATEX agglutination test (Biomeriuex, France).
Effects of phenolic fractions on the viability of E. coli O157:H7
E. coli O157:H7 was grown to late stationary phase in BHI broth, and cells were washed by pelleting via centrifugation at 10,000×g for 10 min twice and resuspended in sterile 0.85% wt/vol NaCl. Treatments of 2.5, 5, and 10% (vol/vol in 0.85% NaCl) of each of the LWB phenolic fractions were created to a final volume of 9 mL and inoculated with 1 mL E. coli O157:H7 to achieve an inoculation level of 108 CFU/mL. The positive control was 70% (vol/vol) isopropanol and the negative control was 0.85% wt/vol NaCl. After inoculation, treatments were incubated at 25°C for 1 h with vortexing every 15 min. Treatments were stopped by centrifuging cells at 10,000×g for 10 min and the pellets were resuspended in 0.85% wt/vol NaCl. This procedure was repeated three times to remove phenolic fractions, and the final cell pellets were resuspended in 0.85% wt/vol NaCl. Viable cell counts were enumerated by spread-plating the cell suspension on MSA plates in duplicate, and the plates were incubated at 37°C before colonies were counted.
Effects of phenolic fractions on the cell membrane permeability of E. coli O157:H7
After fractional treatments, the cell membrane permeability of E. coli O157:H7 was assessed by using the LIVE/DEAD BacLight Bacterial Viability Kit (Invitrogen, Grand Island, NY). When used alone, SYTO 9 dye (excitation/emission at 480/500 nm) generally labels all bacterial cells in a population. In contrast, propidium iodine (PI) dye (excitation/emission at 490/635 nm) penetrates only bacterial cells with damaged membranes, causing a reduction in the fluorescence of SYTO 9 when both dyes are present. Treatment conditions were the same as mentioned in the previous section. After treatment, aliquots of 100 μL of cell suspension were placed into a 96-well plate (Biotech, Winoskii, VT). A 3 mM solution of each dye was prepared in dimethylsulfoxide (Fisher Scientific, Pittsburgh, PA) and diluted to 0.3 mM with distilled water. A 100-μL aliquot of the dye solution was added to each cell suspension and incubated for 15 min in the dark. After incubation, fluorescence intensity was measured by a BMG Labtech spectrophotometer (Cary, NC). The ratio of the integrated intensity between cells stained with SYTO 9 to cells stained with PI was calculated for each treatment to give the relative value of permeabilized cells.
Effects of phenolic fractions on the cell membrane integrity of E. coli O157:H7
Treatment conditions for TEM visualization were the same as mentioned in the previous section. Fractional treatments were screened for growth inhibition using a negative staining of phosphotungstic acid and viewed with a Philips CM10 TEM at an accelerating potential of 80 kV. The lowest concentration that produced visual morphological damage (5% vol/vol) was then selected for higher-resolution ultrathin sectioning. Fixed pellets of 5% vol/vol treatments were then treated with 1% wt/vol OsO4, dehydrated with graded alcohols, and embedded into Epon-Araldite resin. Ultrathin sections cut from these blocks were stained with uranyl acetate and lead citrate (Carson et al., 2002) and examined with a Philips CM10 TEM at an accelerating potential of 80 kV.
Statistical analysis
Log reductions and membrane integrity of E. coli O157:H7 were analyzed by comparing the viable cell counts and the relative fluorescence unit, respectively, between the treatment and the control. Three replicates of each experiment were performed. The numbers of cells were translated into log CFU/mL and treatments were assigned for comparison. Analysis of variance was performed on cell counts using the SAS General Linear Models procedure with SAS software 8.0 (SAS Institute, Cary, NC). Mean separations were examined by least significant difference with significance level at α=0.05.
Results
Characteristics of phenolic fractions
Multiple assays were performed to assess the completeness of the extraction process and the relative concentration of the components of each fractional treatment (Table 1). Phenolic compounds derived from LWB have been extensively characterized (Prior et al., 1998; Schmidt et al., 2005; Kalt et al., 2008). The concentration of each fraction obtained from the extraction process was in agreement with previous research using cranberries and LWB (Wu et al., 2004; Wu et al., 2006; Lacombe et al., 2010; Lacombe et al., 2012). In commercially available LWB, the concentrations of total phenolics and total anthocyanins were approximately 398.0 and 148.2 mg/100 g, respectively (Wu et al., 2006). The pH for each LWB phenolic fraction at 2.5, 5, and 10% in 0.85% wt/vol NaCl are listed in Table 2. The A&P fraction had the lowest pH followed by anthocyanins, MPA, TBP, and proanthocyanidins.
Fractions were analyzed as undiluted blueberry fractions. Results represent the average of three repeats.
Readings were taken before inoculation with bacteria cocktail and after being diluted in 0.85% NaCl. The specific concentration is indicated in parentheses where GAE=gallic acid equivalents.
Log-reducing action of LWB fractional treatment against E. coli O157:H7
Log reductions of E. coli O157:H7 treated by LWB fractions are shown in Figure 1. A significant (p<0.05) reduction of 7 log CFU/mL was observed for MPA at 0.8 g/L GAE and a 5-log CFU/mL reduction at 0.4 g/L GAE. For A&P at 1.8 g/L GAE and 0.9 g/L GAE, a 7- and 5-log CFU/mL reduction was observed, respectively. For anthocyanins, a significant reduction of 6- and 5-log CFU/mL was observed at 1.3 g/L GAE and 0.65 g/L GAE, respectively. At the lowest concentrations of MPA at 0.2 g/L GAE, A&P at 0.45 g/L GAE, and anthocyanins at 0.32 g/L GAE, a 3-, 4-, and 2-log CFU/mL reduction was observed, respectively, whereas there was no significant reduction for proanthocyanidins and TBP. These results are comparable to previous experiments that demonstrated the minimum inhibitory concentrations (MICs) in nutrient-rich conditions for MPA, A&P, and anthocyanins were 2.0 g/L GAE, 1.45 g/L GAE, and 2.0 g/L GAE, respectively (Lacombe et al., 2012). In addition, the MICs for TBP and proanthocyanidins were 1.11 g/L GAE and 2.0 g/L GAE, respectively (Lacombe et al., 2012), which were higher than the concentrations obtained in this experiment.
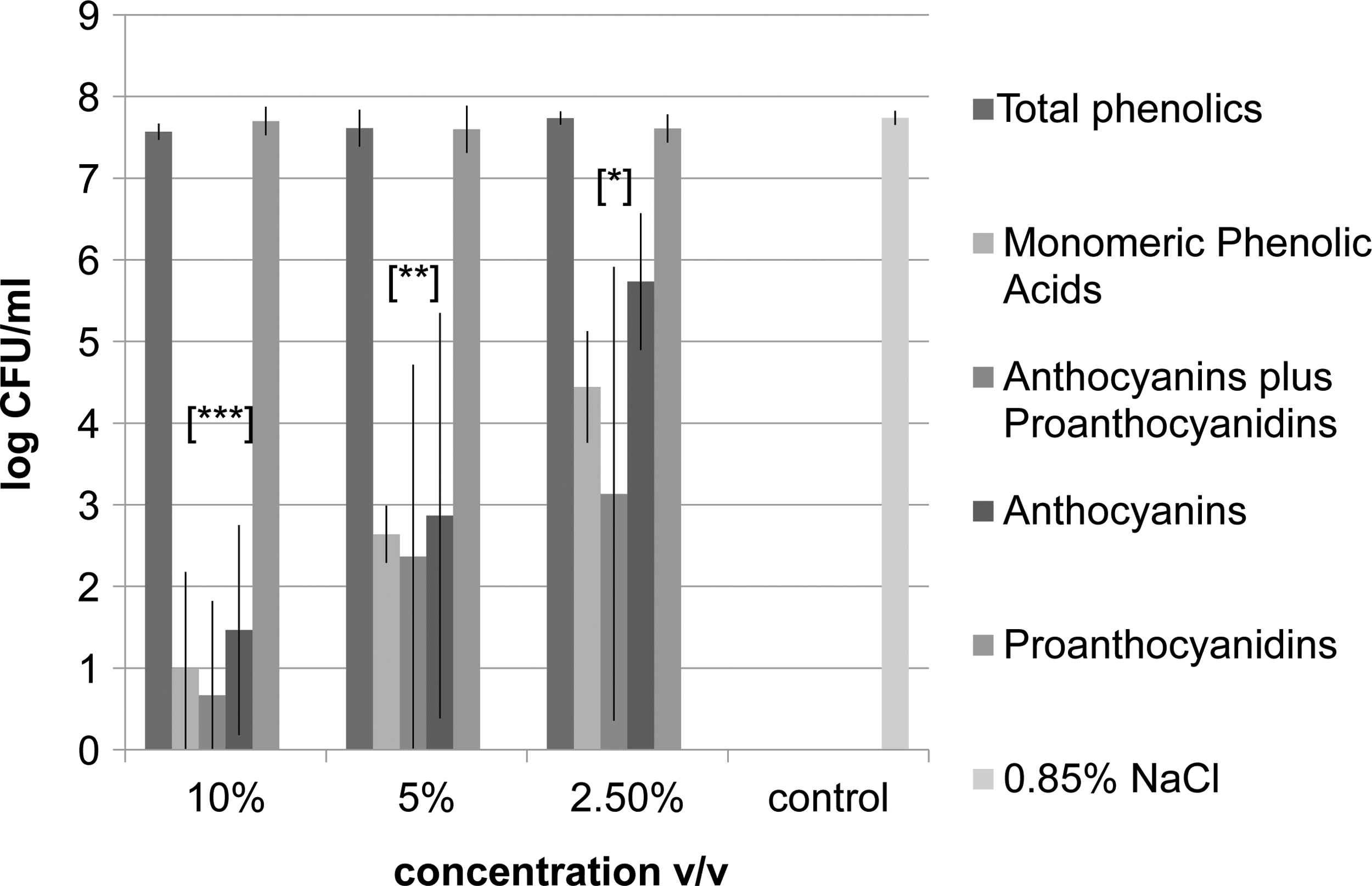
Inactivation of Escherichia coli O157:H7 by lowbush wild blueberry phenolic fractions at 2.5, 5, and 10% in 0.85% wt/vol NaCl. Significant differences (p<0.05) from the control (0.85% wt/vol NaCl) are represented by *, **, and ***, which denote p<0.05, p<0.01, and p<0.001, respectively. CFU, colony-forming units.
Cell membrane permeability of E. coli O157:H7
The cell membrane permeability of E. coli O157:H7 was quantified using the LIVE/DEAD permeability assay. The exclusion of PI was considered an indication of the relative intactness of the cell membrane, and the results are reported as the proportion of fluorescent signal at 500/635 nm, live indicator SYTO9/dead indicator PI (Fig. 2). E. coli O157:H7 treated with TBP at 0.14, 0.27, and 0.55 g/L GAE demonstrated a significant uptake of PI, indicating a significant increase in cell membrane permeability. Comparably, a significant uptake of PI by E. coli O157:H7 was observed for A&P at concentrations of 0.45, 1.8, and 0.9 g/L GAE, for anthocyanins at 0.32, 1.3, and 0.65 g/L GAE, and for proanthocyanidins at 0.15, 0.3, and 0.6 g/L GAE.
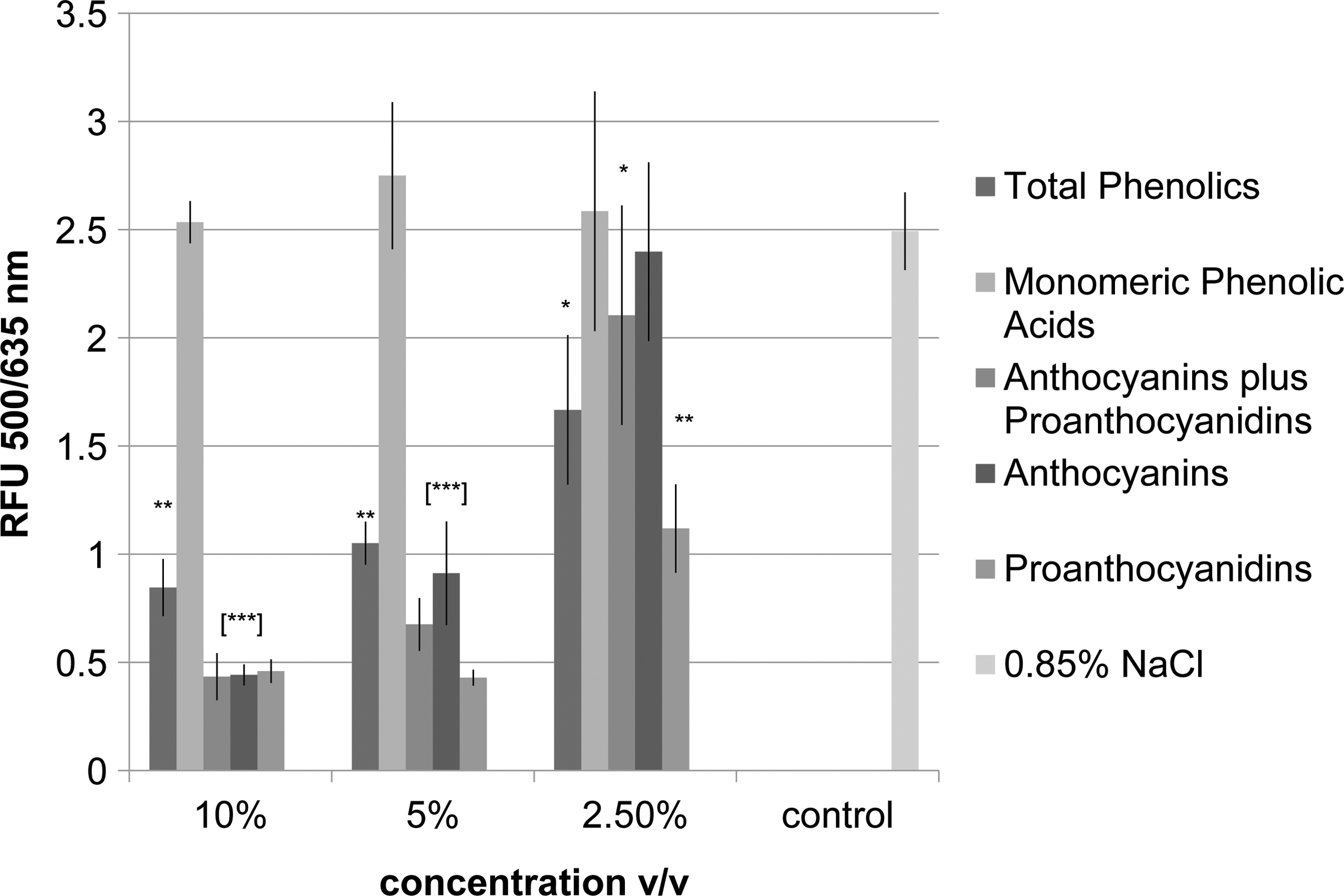
The effect of lowbush wild blueberry phenolic fractions on the membrane permeability of Escherichia coli O157:H7. Significant differences from the control (0.85% wt/vol NaCl) are represented by *, **, and ***, which denote p<0.05, p<0.01, and p<0.001, respectively. RFU, relative fluorescence units.
Morphological damage of E. coli O157:H7 inflicted by LWB
For evaluating the cellular damage induced by each phenolic fraction, cells of E. coli O157:H7 were graded based on the continuity of the cell membrane, consistency of the cytoplasm, and cell shape with respect to the control. Relative intactness was determined by the degree of perforation of the outer membrane and its ability to retain the cytoplasmic constituents. The extent of damage was also graded based on comparisons among the treated cells. A treatment of 5% vol/vol was chosen for thin-section TEM based upon previous experiments that visualized cells using a negative stain technique (data not shown). The control cells displayed a continuous and intact outer membrane, and the cytoplasm was evenly distributed throughout the cells and displayed no aggregation (Fig. 3f). Cells of E. coli O157:H7 treated with TBP at 0.27 g/L GAE showed an indistinguishable outer membrane along with a collapsed shape and irregular form, and the cytoplasmic constituents displayed aggregation (Fig. 3a). The observed morphological damages were consistent among treated cells and different from control cells. Cells treated with MPA at 0.4 g/L GAE (Fig. 3b) displayed localized disintegration of the outer membrane with leeching of cellular constituents into the surrounding environment, irregularity in shape, and slight aggregation in the cytoplasm. Cells treated with A&P at 0.9 g/L GAE displayed localized disintegration and irregularity in the outer membrane with leeching of cellular constituents (Fig. 3c). Cells exposed to 13.9 g/L GAE of anthocyanins and 0.3 g/L GAE proanthocyanidins (Figs. 3d–e) displayed localized disintegration, irregularity in the outer membrane, and leaking of cytoplasm.
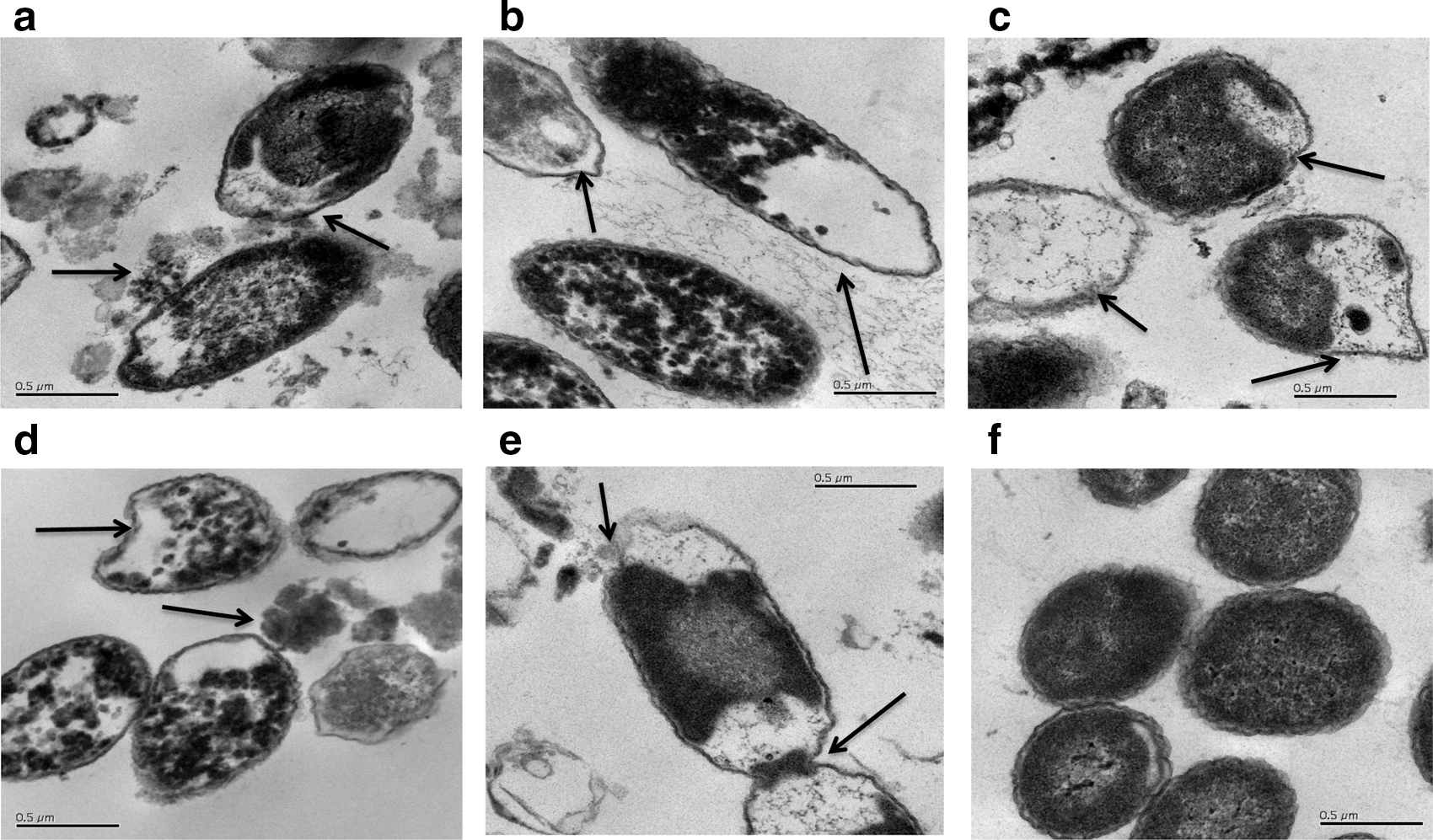
Micrographs of Escherichia coli O157:H7 at 46,000× treated with 5% vol/vol of blueberry fractions:
Discussion
Phytochemicals derived from berries have demonstrated inhibitory effects at the membrane interface and on cytosolic enzymes (Kwon et al., 2007). This study identified the LWB phenolic fractions and concentrations that were capable of inactivating the E. coli O157:H7 and identified the possible mechanism of inactivation. The results are in agreement with previous experiments using similar fractional components of LWB and cranberries that exhibited inactivation and membrane damage against E. coli O157:H7 (Lacombe et al., 2010; Lacombe et al., 2012).
While current reviews and investigations pertaining to microbial inactivation by plant compounds do not provide one definitive mechanism of action, they suggest that the mechanism is due to a concerted activity involving multiple pathways based upon the environmental conditions and the particular microorganisms involved (Friedman et al., 2003; Puupponen-Pimiä et al., 2005; Alakomi et al., 2007; Kwon et al., 2007; Apostolidis et al., 2008). Polyphenols are capable of hydrophobic interactions and hydrogen bonding with membrane proteins (Apostolidis et al., 2008). They can sequester ions required for protein stability (Guo et al., 2007) and donate or accept electrons along the membrane interface (Kwon et al., 2007), allowing them to be versatile antimicrobial agents. In the present study, fractions containing anthocyanins produced the highest degree of permeability and lowest recovery of E. coli O157:H7 cells. In addition, TBP and proanthocyanidins demonstrated a high recovery of E. coli O157:H7 after treatment. This may indicate that the membrane damage caused by these fractions may be reversible if cells are subsequently transferred to favorable growth conditions or that the bacterial inactivation might involve more than one cellular target. MPA did not demonstrate significant uptake of PI; however, it did cause significant reductions in viable E. coli O157:H7 cells and membrane damage. In a previous work, flow cytometric analysis showed that dye exclusion might have affected the direct fluorescence measurements (Torres et al., 2007). Nonetheless, the TEM experiments confirmed that all LWB phenolic fractions at 5% vol/vol increased cell membrane permeability, as indicated by the leaching of cytoplasmic components into the surrounding environment.
Berries are acidic by nature, with a pH ranging from 2 to 3.5. Wu et al. (2008) examined the effect of low pH and its contribution to the growth inhibition of E. coli O157:H7 using organic acids commonly found in cranberry and blueberry. The acid solutions were buffered at pH 4.7 and 3.5 and compared to cranberry concentrates (25 μL/mL and 100 μL/mL) at the same pH. The results indicated that low pH plays a role in inhibition. However, cranberry extract exhibited additional antimicrobial properties with an increased reduction of approximately 1 log CFU/mL (Wu et al., 2008). In another previous study, neutralized cranberry organic acids, monomeric phenolics, and anthocyanins were compared to their native pH fractions to determine their antimicrobial activity against E. coli O157:H7. Cranberry organic acids relied solely on low pH, as they exhibited growth inhibition at pH 3.5 and no inhibition at pH 7 (Lacombe et al., 2012). In contrast, MPA demonstrated the same inhibition at pH 3.7 and pH 7, suggesting a pH-independent mechanism of inhibition. Neutralization of anthocyanins doubled the MIC against E. coli O157:H7, suggesting its reliance on low pH (2–3) for structural stability as a part of the antimicrobial mechanism. These findings have major implication toward the utilization of cranberry extracts as food-grade antimicrobials and may be extended to LWB.
Food scientists are challenged to develop new preservation systems that will improve the quality and extend the shelf life of food products without compromising safety or sensory qualities. Berries have been used as an alternative to nitrates in meat products because of their ability to reduce microbial contamination and their perceived health benefits (Wu et al., 2009; Khanal et al., 2010; Eells et al., 2011). Recent studies have demonstrated that cranberry extract at 7.5% wt/wt was capable of inactivating E. coli O157:H7 in ground beef without adversely affecting the product's taste and other sensory qualities (Wu et al., 2009). The potential applications of LWB extracts include adding the extracts into ready-to-eat foods such as dressings, deli meats, salads, and in juices or juice-cocktail combination for pathogen reduction and shelf-life extension. Fruit juices contaminated with enteric bacterial pathogens such as E. coli O157:H7 and Salmonella species have caused outbreaks of foodborne illness; therefore, the U.S. Food and Drug Administration (FDA) requires a process performance standard of a 5-log CFU/mL reduction for the pathogens in these products (Mosqueda-Melgar et al., 2008). The present study demonstrated a >5-log CFU/mL reduction of E. coli O157:H7 by MPA, A&P, and anthocyanins from LWB, which meets the FDA's performance standard. Therefore, LWB may be applied as a part of hurdle technology, possibly in conjunction with other processing technologies.
Footnotes
Acknowledgments
The authors of this article would like to thank Electron Microscopy facility, Mr. Kelly Edwards, and Dr. Seth Tyler at the University of Maine for their assistance in this research. This research was supported by USDA NNF under agreement No. 2007-38420-17764 and Maine Agricultural and Forest Experiment Station at the University of Maine with external publication number 3325. Work is based upon research supported in part by Hatch Grant number ME08562-10 from the USDA National Institute of Food and Agriculture.
Disclosure Statement
No competing financial interests exist.