
Abstract
Salmonella enterica serovar Enteritidis is a leading cause of salmonellosis throughout the world and is most commonly associated with the consumption of contaminated poultry and egg products. Salmonella Enteritidis has enhanced ability to colonize and persist in extraintestinal sites within chickens. In this study, 54 Salmonella Enteritidis isolates from human patients (n=28), retail chicken (n=9), broiler farms (n=9), and egg production facilities (n=8) were characterized by antimicrobial susceptibility testing, plasmid analysis, genetic relatedness using XbaI and AvrII pulsed-field gel electrophoresis (PFGE), and the presence of putative virulence genes. Nine isolates were evaluated for their abilities to invade and survive in intestinal epithelial and macrophage cell lines. Overall, 56% (n=30) of isolates were resistant to at least one antimicrobial agent tested, yet no isolates showed resistance to more than three antimicrobials. All isolates carried a common ∼55-kb plasmid, with some strains containing additional plasmids ranging from 3 to 50 kb. PFGE analysis revealed five XbaI and AvrII clusters. There were significant overlaps in the PFGE patterns of the isolates from human, chicken, and egg houses. All isolates tested PCR positive for iacP, purR, ttrB, spi4H, rmbA, sopE, invA, sopB, spvB, pagC, msgA, spaN, orgA, tolC, and sifA, and negative for iss, virB4, and sipB. Of the isolates selected for virulence testing, those containing the iron acquisition genes, iutA, sitA, and iucA, and ∼50-kb plasmids demonstrated among the highest levels of macrophage and epithelial cell invasion, which may indicate their importance in pathogenesis.
Introduction
Salmonella Enteritidis is a facultative intracellular pathogen and can cause invasive infections in poultry and humans (Vugia et al., 2004). In contrast to most other serotypes, Salmonella Enteritidis present in laying hens has the unique ability to contaminate shell-egg contents through the transovarian route following colonization of the intestinal tract (Thiagarajan et al., 1994; Humphrey, 2004; Gantois et al., 2009). Salmonella Enteritidis appears to have an enhanced ability to colonize and reach extraintestinal sites within chickens. Some Salmonella Enteritidis virulence is likely associated with their conserved virulence plasmid, Salmonella pathogenicity islands, and presence of multiple fimbriae (Thomson et al., 2008; Gantois et al., 2009; Betancor et al., 2012). Due to public health importance, the serotype has been the target of multiple recent whole genome sequencing projects aimed at learning more about the genetics of the pathogen (Timme et al., 2012; Allard et al., 2013). However, the exact mechanisms involved in Salmonella Enteritidis pathogenicity in humans and food-producing animals remains poorly understood.
In humans, Salmonella Enteritidis generally causes a relatively mild, self-limited gastroenteritis (Silva et al., 2012); however, some cases may involve invasive infections, for which antimicrobial therapy is usually warranted. While Salmonella Enteritidis often displays less widespread antimicrobial resistance than some other serotypes, antimicrobial-resistant strains have been reported from several countries around the world (Cardinale et al., 2005; Dias de Oliveira et al., 2005; Kang et al., 2009; Vaz et al., 2010; Zou et al., 2012).
The objectives of this study were to characterize antimicrobial resistance and virulence in a subset of Salmonella Enteritidis from humans, chicken, and egg production environments, and compare the diversity or similarity among them and identify potential factors that may play a role in increased virulence in in vitro cell culture models.
Materials and Methods
Bacterial strains
Fifty-four Salmonella enterica serovar Enteritidis strains isolated from humans (n=28), retail poultry (n=9), broiler farms (n=9), and egg production facilities (n=8) were included in the study (see Fig. 1 for more detailed information on the isolates). The isolates were stored in brain heart infusion broth with 20% glycerol at −80°C and streaked on tryptic soy agar with or without 5% sheep's blood (Remel, Lenexa, KS) and incubated at 37°C for 18 to 24 h for the individual analyses.
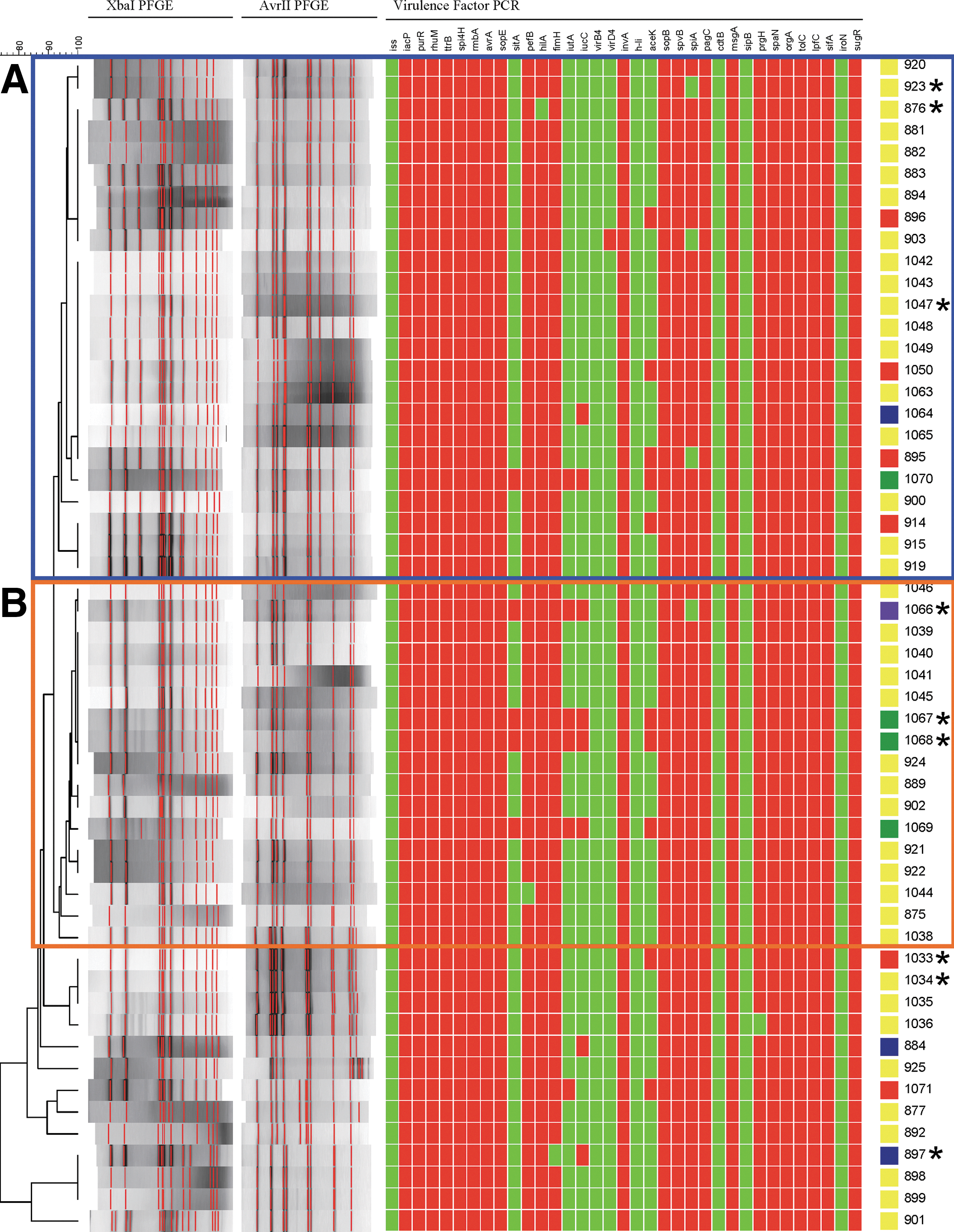
Results of pulsed-field gel electrophoresis (PFGE) analysis and plasmid analysis and virulence factor polymerase chain reaction (PCR). The dendrogram is based on the composite XbaI/AvrII PFGE restriction digestion profiles analyzed using Dice coefficients. Two major PFGE clusters
Pulsed-field gel electrophoresis (PFGE)
PFGE was performed following restriction digestion with either 50 U of XbaI or 8 U of AvrII (New England Biolabs, Ipswich, MA) according to PulseNet methods (Ribot et al., 2006). The digested DNA was separated using a Chef Mapper (Bio-Rad, Hercules, CA), and the gels were stained with Lonza GelStar® Nucleic Acid Gel Stain (Cambrex Bio Science, Walkersville, MD) and analyzed using BioNumerics software (Applied Maths, Kortrijk, Belgium). Salmonella Braenderup H9812 (ATCC BAA-664) digested with XbaI was used as the size standard for normalization of the band patterns. PFGE patterns were identified and named based on comparison of the XbaI profiles to defined PFGE restriction patterns. The relatedness of the gel band patterns was calculated using Dice coefficients with a 1% band tolerance and 1.5% optimization. A dendrogram was generated using the unweighted pair group method using arithmetic averages by averaging the similarities of the XbaI and AvrII experiments.
Antimicrobial susceptibility testing
Antimicrobial minimum inhibitory concentrations for Salmonella isolates were determined using a manual broth microdilution method following the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2008). The following antimicrobials were tested: ampicillin, chloramphenicol, ciprofloxacin, gentamicin, kanamycin, nalidixic acid, streptomycin, sulfisoxazole, and tetracycline. Escherichia coli ATCC 25922 was used as a quality-control organism for the testing, and the results were interpreted according to the CLSI guidelines (CLSI, 2008), with the exception of streptomycin, for which the National Antimicrobial Resistance Monitoring System breakpoint (32 μg/mL) was used. Polymerase chain reaction (PCR) was used to screen for the presence of antimicrobial resistance genes associated with sulfonamide resistance. The primers (Supplementary Table S1; Supplementary Data are available online at
Plasmid profile analysis
Plasmid DNA was isolated using the Qiagen® Plasmid Mini Kit (Qiagen, Valencia, CA) following the manufacturer's protocols. The isolated plasmids were separated in 0.7% SeaKem® LE agarose (Cambrex Bio Science) gels prepared with 1X Tris-borate-EDTA buffer (Fisher Biochemicals, Baltimore, MD) at 4 V/cm for 6 h at room temperature, and stained with Lonza GelStar® Nucleic Acid Gel Stain. The gels were visualized and plasmid sizes were determined based on the relative migration to the BAC-Tracker™ Supercoiled DNA Ladder (Epicentre, Madison, WI).
Virulence gene detection
PCR was used to amplify the following targets using the conditions described in the referenced papers: spvB, spiA (also named yscC), pagC, cdtB, msgA, sipB, prgH, spaN, orgA, tolC, iroN, lpfC, and sifA (Skyberg et al., 2006), sopB, invA, h-li and aceK (Nayak et al., 2004), sitA, fimH, iss, iutA, and iucC (Rodriguez-Siek et al., 2005), pefB and hilA (Foley et al., 2006), rmbA, avrA, rhuM, ttrB, spi4H, iacP, purR, sopE, and sugR (Zou et al., 2011), and virB4 and virD4 (this study). Primers sequences are provided in Supplementary Table S1; virB4 and virD4 were amplified as described by Foley et al. (2006), with an annealing temperature of 58°C. Sequence-confirmed positive controls were included with all sets of reactions. PCR products were analyzed using 96-well 2% E-gels (Invitrogen, Carlsbad, CA). The results were imported as binary (positive/negative) data into BioNumerics, and the cumulative virulence profiles were analyzed using principal components analysis to separate the isolates (David et al., 2010).
Macrophage and intestinal epithelial cell invasion assays
Based on the PCA analysis, nine Salmonella Enteritidis isolates and E. coli OneShot Top10 (Invitrogen) were chosen for further virulence studies using J774 mouse macrophage cells (TIB-67; American Type Culture Collection [ATCC], Manassas, VA) and rat epithelial cells (CRL-1592; ATCC). For the macrophage experiments, cells were grown in 75-mm culture flasks in Dulbecco's modified Eagle's medium (DMEM) supplemented with 2 mM glutamine, 10% fetal bovine serum (FBS), penicillin (100 U/mL), and streptomycin (100 μg/mL) at 37°C with 5% CO2 until they reached complete monolayer. The cells were detached by scraping, washed, suspended in DMEM, and plated in 24-well culture plates at a density of 4×105 cells per well. The plates were incubated at 37°C with 5% CO2 for 48 h. The epithelial cells were cultured in DMEM supplemented with 4 mM glutamine, 1.5 g/L sodium bicarbonate, 4.5 g/L glucose, 0.1 U/mL bovine insulin, and 10% FBS at 37°C with 5% CO2 until they reached a confluent monolayer in 75-mm culture flasks. After this, the cells were detached by incubating in 25% trypsin, 0.53 mM EDTA solution at 37°C for 15 min. The detached cells were washed, suspended in DMEM, and plated in a 24-well culture plate at a density of 5×104 cells per well and incubated at 37°C with 5% CO2 for 48 h.
For invasion studies, bacteria were grown to mid log-phase in LB broth, their cell density determined using a SmartSpec (Bio-Rad), centrifuged, and suspended in DMEM to a density of 8×108 cells/mL for macrophage assays and 1×108 cells/mL for epithelial cell assays. A 100-μL aliquot of each bacterial suspension was added to two wells of the culture plate in each experiment. In each well there was a 200:1 bacterial-to-host-cell ratio. The plates were centrifuged at 500 rpm for 5 min and incubated for 1 h at 37°C in 5% CO2. After incubation, the cells were washed with PBS three times and incubated with 50 μg/mL of gentamicin for 1 h at 37°C to kill the extracellular bacteria. After incubation, the cells were washed to remove the antibiotic and treated with 1% Triton X-100 for 5 min at 37°C to lyse the host cells. The suspensions were serially diluted in phosphate buffered saline (PBS) buffer, plated onto LB agar plates, and incubated overnight at 37°C. The next day the cells were counted to determine the number of bacterial cells that survived the host cells. Each set of experiments was repeated at least three times and the results were averaged and standard deviations were determined. Bacterial survival rates were calculated by dividing the average number of bacterial cells that survived by the number of cells used to infect the host cells. Noninfected host cells served as negative controls for the invasion assays.
Results and Discussion
This study examined antimicrobial resistance and virulence in 54 Salmonella Enteritidis isolates from human patients and egg- and poultry-related sources. To determine the potential overlap of genotypes among isolates from different sources, PFGE analyses following digestion of bacterial DNA with XbaI and AvrII were conducted. Two large clusters were identified based on an average 93% combined similarity from both PFGE methods (Fig. 1). Cluster A contained 24 isolates with the PFGE pattern JEGX01.0005 from chicken farm, meat, and human sources, and Cluster B contained 17 isolates with pattern JEGX01.0004 from egg house, chicken, and human sources (Fig. 1 and Table 1). The detection of common PFGE profiles indicates the potential for transmission of Salmonella Enteritidis from poultry and egg sources to human patients; however, this study did not focus on the causality of the human infections or examine the specific temporal relationships of the isolates.
Shaded isolates were those characterized in the macrophage and intestinal epithelial cell invasion assays. Resistance was detected to the following antimicrobials: ampicillin (Amp), nalidixic acid (Nal), streptomycin (Str), sulfisoxazole (Sul) and tetracycline (Tet).
PFGE, pulsed-field gel electrophoresis; PCA, principal components analysis; AR, Arkansas; WV, West Virginia.
Thirty isolates (56%) were resistant to at least one of the antimicrobial agents tested (Table 1). Resistance was detected to sulfisoxazole (n=29, 54%), followed by nalidixic acid (n=4, 7.4%), streptomycin, ampicillin, and tetracycline (n=3, 5.6%). Eight (15%) isolates were resistant to more than one antimicrobial tested, with two isolates each resistant to ampicillin, sulfisoxazole and nalidixic acid, or tetracycline and sulfisoxazole. The rate of sulfisoxazole resistance in this study was higher than those reported in the other studies (Crump et al., 2011; FDA, 2011; Zou et al., 2012) but similar to those reported by Melendez et al., 2010, whose isolates from retail chicken also displayed PFGE profiles that were similar to those in the present study. To determine the genetic factors contributing to sulfisoxazole resistance, different sets of PCR reactions were used to identify resistance genes sul1, sul2, and sul3; however, no products were amplified in the sulfisoxazole-resistant isolates, even though the positive controls were amplified. Interestingly, while others have reported these genes in sulfisoxazole-resistant isolates, the present findings are in agreement with those reported by Perugini et al. (2010), who were unable to detect the genes in their sulfisoxazole-resistant Salmonella Enteritidis isolates. Combined, these results suggest that other, unidentified resistance mechanisms play a role in sulfisoxazole resistance in certain Salmonella Enteritidis isolates.
In order to evaluate the potential factors that may contribute to the ability of Salmonella Enteritidis to cause an infection, 35 putative virulence genes were investigated by PCR. All of the isolates were PCR positive for iacP, purR, ttrB, spi4H, rmbA, sopE, invA, sopB, spvB, pagC, msgA, spaN, orgA, tolC, sifA, lpfC, rhuM, sugR, and avrA, indicating that these virulence genes are widespread in Salmonella Enteritidis. In contrast, all isolates were negative for iss, h-li, virB4, sipB, cdtB, and iroN (Fig. 1). Of the other virulence genes found in more than one isolate, pefB, hilA, fimH, prgH, avrA, and tolC were each found in 53 (98%) isolates, while spiA was detected in 50 (93%), aceK in nine (17%), iucC was found in eight (14%), and iutA and sitA in five (9.3%) isolates (Fig. 1). The five isolates positive for iutA and sitA were also positive for iucC. These three genes are associated with iron acquisition systems and are typically found on common plasmids detected in avian pathogenic E. coli and certain Salmonella serotypes that are prominent in poultry, including Kentucky, Heidelberg, and Typhimurium (Johnson et al., 2010).
When the results of the virulence PCR were compared to Salmonella Enteritidis whole genome sequences available through the PATRIC database (which includes the GenBank cataloged genomes) (Gillespie et al., 2011), none of the genomes (as of April 17, 2013) were positive for iutA, iucC, or the plasmid-associated sitA genes. The majority of the sequenced genomes contained aceK (100/110) and spiA (109/110), which is a higher percentage, especially for aceK, than detected in the current study. These differences may be due in part to the characteristic that the vast majority of available genome sequences are from isolates associated with a 2010 Salmonella Enteritidis outbreak associated with eggs (Timme et al., 2012; Allard et al., 2013).
A number of antimicrobial resistance and virulence genes can be carried on plasmids. Most Salmonella Enteritidis strains contain a virulence plasmid of approximately 55–60 kb in size with genes that carry the Salmonella plasmid virulence (spv) and plasmid encoded fimbriae (pef) genes (White et al., 2000). In our study, a ∼55-kb plasmid was detected in all of the isolates (Table 1) and tested positive for spvB and pefB (with one exception each). Many of the isolates carry other plasmids as well, including 13 that have three additional plasmids of approximately 10, 5, and 3 kb. Other isolates were found to have 50-kb (n=5) and 40-kb (n=1) plasmids.
When the results of the virulence gene PCR were analyzed using PCA, there seemed to be five groupings of isolates that ranged in size from a single isolate (purple group) to 40 isolates (yellow group, Fig. 2). The PCA analysis utilizes algorithms to identify the maximum variance in the data and group isolates based on these parameters. The most discriminatory virulence factors that impacted the PCA were spiA, aceK, iutA, sitA, and iucC (results not shown), due to their relative variability in frequency among the isolates in the population. From these groups, a subset of nine isolates were semirandomly selected (a minimum of one isolate was chosen from each group) for further in vitro virulence testing in macrophage and intestinal epithelial cell cultures.

Results of principal components analysis of virulence factor polymerase chain reaction data shown in Figure 1. Isolates that clustered near one another were joined in groups, which are displayed as different colors on the coordinate plane. The cluster and subgroups to which each of the isolates belong are displayed in the PCA ID column of Table 1.
The isolates selected were incubated with rodent macrophages and intestinal epithelial cells, which were selected for the invasion studies due to their roles in Salmonella pathogenesis (Foley and Lynne, 2008). An initial stage of infection is the ability to enter and survive in intestinal cells, which is associated with the signs and symptoms of gastroenteritis. For more invasive infection, a key feature is the ability to survive in immune cells, such as macrophages; thus, these two cell types were chosen to evaluate virulence. The rodent cell lines are likely appropriate surrogate models to evaluate for Salmonella Enteritidis, since the serotype is known to effectively colonize rodents and serve as a potential vector for the spread of Salmonella Enteritidis in poultry (Callaway et al., 2008). The results of the invasion assays are shown in Figure 3. For the intestinal epithelial cells, cells from isolate 923 had the highest percentage of bacteria invading and surviving within the cell line after 1-h invasion, followed by isolates 1067, 1068, and 1066, respectively (Fig. 3A). Isolates 1067, 1068, and 1066 also displayed the highest ability to survive within macrophages. Isolates 1067 and 1068 displayed on average a greater than 10-fold higher survival than isolates 923, 897, 1033, and 1047 (Fig. 3B). These results indicate marked differences among the isolates in their ability to survive in macrophages. The E. coli Top10 cells, which lack type 3 secretion system genes present in Salmonella, were not able to enter epithelial cells and were minimally able to survive in macrophages after 1 h (0.003±0.001% survival; Fig. 3). Interestingly, three iron acquisition genes, iucC, iutA, and sitA were detected in isolates 1066, 1067, and 1068 and all three isolates harbored 50-kb plasmids (Fig. 1 and Table 1). These iron acquisition genes are typically plasmid-associated, and plasmids carrying these iron acquisition systems can play a role in avian pathogenicity in Salmonella and E. coli (Johnson et al., 2010). Work is ongoing to characterize the genes present on the 50-kb plasmids. Isolate 923, which displayed the greatest intestinal epithelial cell invasion, did not have any major virulence profile differences from the other isolates in its PCA/virulence grouping that would explain its increased invasiveness. Interestingly, spiA (yscC), an important effector associated with Salmonella pathogenicity island 2 (SPI-2) (Foley and Lynne, 2008), was not detected in isolate 923. This lack of spiA may explain why isolate 923 was poorly able to survive in macrophages, since SPI-2 is important for survival in macrophages. aceK encodes the isocitrate dehydrogenase kinase/phosphatase enzyme that is important for microorganisms to utilize acetate as a sole carbon source (Zheng and Jia, 2010). There did not seem to be a correlation between the presence of aceK and virulence; while aceK-positive isolates 1067 and 1068 displayed relatively high levels of invasion, PCR-positive isolate 1033 did not (Fig. 3).
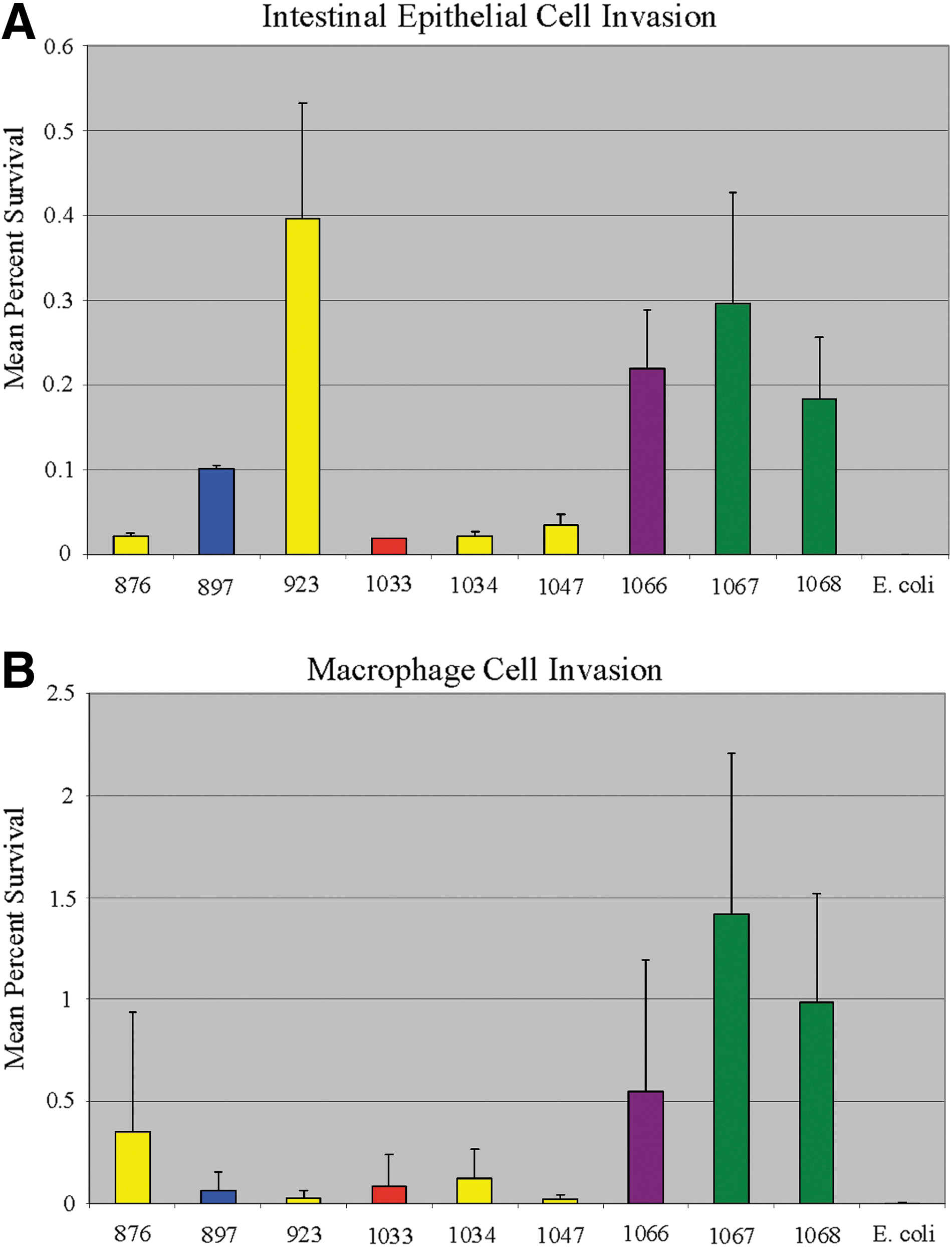
Results of the intestinal epithelial cell
Conclusions
In many instances, Salmonella Enteritidis isolated from humans shared high similarity with those isolated from chicken and egg-associated sources when evaluated using PFGE, antimicrobial susceptibility, plasmid analysis and virulence gene profiles. The resistance rate of the isolates in this study to sulfisoxazole was higher compared to many studies reported in literature. However, the common sulfisoxazole-associated resistance genes—sul1, sul2, sul3—were not detected, indicating that other genes likely contributed to resistance in these isolates. Three Salmonella Enteritidis isolates from retail chicken containing iron acquisition-associated genes (iucC, iutA, and sitA) and a 50-kb plasmid demonstrated the greatest ability to invade and survive in murine macrophages and were among the best at invading the intestinal epithelial cells. The factors that impacted the ability of isolate 923 to invade epithelial cells were unclear; therefore, the mechanisms by which the putative virulence factors impact survival of Salmonella Enteritidis in macrophages and epithelial cells warrant further research.
Footnotes
Acknowledgments
The authors would like to thank Drs. Carl Cerniglia, Huizhong Chen, and Mohamed Nawaz for their critical reviewing of the manuscript. Dr. Jing Han, Dustyn Barnette, and Anthony Rooney are supported through the Oak Ridge Institute for Science and Education. Dr. Kuppan Gokulan is supported by an FDA Commissioner Fellowship Program and by Oak Ridge Institute for Science and Education.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.