
Abstract
Vibrio parahaemolyticus infection has been considered the leading cause of bacterial illnesses mainly associated with seafood consumption in Guangdong province in China. In this study, epidemiological and etiological characteristics of 36 V. parahaemolyticus outbreaks that occurred from 2008 to 2010 in Guangdong province were analyzed; 284 strains involved were characterized by serotyping; virulence genes and 66 strains from four outbreaks therein were subtyped by pulsed-field gel electrophoresis (PFGE). Epidemiological evidence showed that 36% (13/36) of outbreaks were caused by food contamination, of which 84.6 % (11/13) were related to salted food, including viscera of pigs and cattle, meat, and vegetable salad. It was also indicated that 88.9% (32/36) of V. parahaemolyticus outbreaks appeared from June to September, 44.4% (16/36) of which occurred in canteens as well as 41.7% (15/36) in restaurants. As for the etiology, 31% (11/36) of outbreaks were caused by single serovar of strains, while 69% (25/36) were caused by multiserovars; O3:K6, O4:K8, O1:Kut, and O2:K3 were the dominant serovars. Among the 284 strains, 98.8% (254/257) of strains from patients were tdh-present and trh-absent, whereas 37.0% (10/27) from food were tdh-present. Cluster analysis of PFGE patterns demonstrated that strains in the same outbreak with identical serovar seemed to be diversified, whereas strains with various serovars could be closely related genetically. Moreover, cross-contamination between salted food and seafood was first confirmed by molecular subtyping in Guangdong, revealing that salted food might be a vital risk factor associated with V. parahaemolyticus outbreaks.
Introduction
V
Serotyping is a conventional method to distinguish phenotypes of V. parahaemolyticus strains. With specific antisera, V. parahaemolyticus strains can be divided into 11 O groups and 71 K antigens (Nair et al., 2007). Since 1996, the cases of V. parahaemolyticus infection have increased dramatically worldwide due to a pandemic spread of newly emerging clone named new O3:K6 serovar (Chiho et al., 2000). Since then, besides O3:K6, multiple serovars such as O1:K56, O4:K68, and O1:Kut (UT, untypeable) have become the major prevalent serovars that are responsible for most of outbreaks in America, Asia, and Africa (Chiho et al., 2000; Jaime et al., 2004; Oralak et al., 2007; Ansaruzzaman et al., 2008). To prioritize outbreak investigation more accurately, it is necessary to determine the genetic relations among strains with various serovars by molecular subtyping. In recent years, pulsed-field gel electrophoresis (PFGE) had been regarded as a criterion standard of molecular subtyping employed to distinguish strains among V. parahaemolyticus outbreaks via fingerprinting patterns (Phuangthip et al., 2007; Oralak et al., 2007; Ansaruzzaman et al., 2008). By means of PFGE subtyping, it was evidential that certain patterns of pandemic strains were remarkably similar to that of O3:K6 serovars, suggesting that these strains were closely related (Harth et al., 2009; Ansaruzzaman et al., 2008; Kam et al., 2008).
Since 2008, V. parahaemolyticus have become one of the most vital foodborne pathogens in Guangdong province, probably since Guangdong is located in a subtropical coastal region, harvests large quantities of seafood, and enjoys high consumption (Ma et al., 2009). Our previous surveillance results also indicated that seafood had high densities of V. parahaemolyticus contamination (Fang et al., 2009; Tan et al., 2010). Nevertheless, it is worth further noting that local people consumed salted food (salinity exceed to 3%) more commonly than seafood as one type of daily dish in reality. Salted food, which is mainly made from viscera of pigs and cattle, frequently appear in V. parahaemolyticus outbreaks reported by epidemiology investigation (Yan et al., 2006; Ma et al., 2009). Therefore, to clarify the causes of outbreaks, the impact of salted food should be considered as well.
In this study, to determine the epidemiological and etiological characteristics of V. parahaemolyticus emerging in Guangdong province, the causal factor, and the spatial and temporal distribution in 36 outbreaks were studied along with 284 strains from patients or the food vehicles were characterized by serotyping and virulence genes. The genetic relatedness of 66 strains in four outbreaks therein was analyzed via PFGE subtyping.
Materials and Methods
Case definition and classification
According to a foodborne illnesses notification network based on the “National Public Health Emergency Events Surveillance System” established by China CDC (Li et al., 2009), we conducted a retrospective analysis of data on 36 outbreaks emerged in nine cities and reported by epidemiologists in Guangdong Province during 2008–2010. Outbreak was defined as an incident in which two or more persons experience a similar illness occurring within 4 to 30 hours after dining and resulting from consumption of a certain food vehicle. Suspected cases were defined as patients with abdominal pain, diarrhea (≥3 times per day) or symptoms of nausea, vomiting, and fever. The food vehicle consumed by suspected cases was defined as a sort of food contaminated by V. parahaemolyticus, which was confirmed by microbiology testing of remnants or the same lot of product. The causes of outbreaks, and spatial and temporal distribution of incidents were analyzed in this study. The spatial distribution of outbreak was classified to four types of sites, including “in the restaurant,” “in the canteen,” “at home,” and “on travel.” It was noted that “restaurant” served seafood as a daily supply, whereas “canteen” seldom offered seafood dishes at the local level. SPSS 13.0 (IBM, U.S.A.) and Excel (Microsoft, U.S.A.) software were used for statistical analysis by Mann–Whitney mean for nonparametric test and Pearson mean for chi-square test.
Microbiological analysis
Specimens including stool, rectal swab, and vomit from suspected cases and various suspected food vehicles within these outbreaks were collected and within 4 hours enriched by alkaline peptone water with 3% sodium chloride for 24 h at 36°C. Enriched culture was plated onto Thiosulfate Citrate Bile Sucrose Agar (TCBS, HuanKai, China) for 18–24 h incubation. Green colonies were selected and identified based on biochemical profiles by API 20E (Bio Merieux, France) biochemical test strips (USFDA, 2005). A total of 284 strains (257 strains from patients and 27 from food vehicles) were preserved and subsequently delivered to Guangdong CDC for further identification.
Polymerase chain reaction (PCR) amplification
V. parahaemolyticus strains were grown in tryptic soy agar medium with 3.5% NaCl at 37°C for 24 h. The culture (a loop, 10 μL) was transferred into 2-mL tubes containing 500 μL sterile water, the concentration of which was approximately 50 ng/μL. The suspension was heated for about 10 min at 100°C and then cooled on ice for 5 min. The DNA in suspension was separated from cell debris by centrifugation at 10,000 rpm for 3 min and followed by 5 min in ice. The collected supernatant was used as the source of DNA templates for PCR. PCR reaction was performed by the Premix Ex Taq™ kit (Takara, Japan). Sense and anti-sense primers were commissioned to synthesize (Sangon, China). tdh gene was amplified using primer TDH1: 5'-agcttccatctgtccctttt-3' and TDH2: 5'-attaccactaccactctcata-3'; trh gene was amplified using R2:5'-ggctcaaaatggttaagcg-3' and R6:5'-catttccgctctcatatgc-3' (USFDA and CFSAN, 2005). Composition of the reaction system (25 μL) was as follows: 12.5 μL 2×Premix Taq contained 1.25 U/μL of Taq enzyme, 2×dNTP mixture with 0.4 mM each, 1×Ex Taq buffer with 4 mM Mg2+, 0.5 μL of each primers (0.4 μmol/L), 1 μL of DNA template (<500 ng) and 9.5 μL sterile ddH2O. PCR amplification was performed with a single cycle at 94°C for 5 min, followed by 35 cycles of denaturation at 94°C, annealing at 58°C for 30 s, and extension at 72°C for 60 s, with a additional extension at 72°C for 4 min. Five microliters of PCR products was electrophoresed through 1.5% agarose and stained by Goldeview® dye (Parkson, China) and photographed with the Gel Doc 2000 system (Bio-Rad, USA). V. parahaemolyticus ATCC 33847 (tdh+, trh−; amplified fragment size is 434 bp) and ATCC 17802 (tdh+, trh+; amplified fragment size is 250 bp) were the positive controls along with PCR amplifications (ATCC, USA).
Serotyping
Serotyping was conducted according to the method described in the National standards for detection of V. parahaemolyticus using the commercial antisera (Denka, Seiken Corp., Japan) based on the lipopolysaccharide (O) and capsular (K) antigens (Ministry of Health, China, 2008).
Molecular subtyping
PFGE was performed in accordance with the standard laboratory protocol for molecular subtyping of V. parahaemolyticus published by the PulseNet, CDC, Atlanta, GA (Kam et al., 2008). Chromosomal DNA of the tested strains was digested with 20 U of NotI (Promega, United Kingdom) at 37°C for 4 h in 0.5X TBE buffer (44.5 mM Tris-HCl, 44.5 mM boric acid, 1.0 mM EDTA, pH 8.0) for an 18.5-h run maintained at 14°C by Auto-algorithm mode of CHEF DRIII system (Bio-Rad, .USA) using a 78-kb–396-kb size range. DNA from Salmonella Braenderup H9812 was used as marker of molecular size. DNA bands were visualized and photographed with the Gel Doc 2000 system (Bio-Rad, USA) and the NotI-PFGE restricted patterns were compared by Bio Numerics software (Applied Maths, Belgium).
Results
Causes of outbreaks
In this study, only 36% (13/36) of outbreaks were confirmed by microbiology identification to result from certain food vehicles, while the remaining 64%(23/36) were still unknown (Table 1). Salted food seemed to be more common in causing V. parahaemolyticus outbreaks than seafood: 84.6% (11/13) of confirmed outbreaks were associated with salted food, whereas seafood was only 38.46% (5/13) (of these, four of five occurred simultaneously in outbreaks with seafood and salted food). The incidence of infections by salted food consumption was significantly higher than that by seafood (chi-square value was 4.248, p=0.039). In addition, it was suggested that the types of salted food were diversified, mainly including viscera of pigs and cattle, meat, and vegetable salad (Table 1).
The food vehicle was confirmed by microbiology isolation and identification.
Spatial distribution
The breakdown of spatial distribution was as follows: 44.4% (16/36) of outbreaks occurred in canteens and 41.7% (15/36) in restaurants, whereas only 8.3% (3/36) and 5.5% (2/36) of outbreaks emerged during traveling or dining at home, respectively. While comparing to the sites of outbreaks that taken place (Fig. 1), it was demonstrated that the majority occurred in the Pear River Delta and coastal cities of Guangdong province. Dongguan city, which located in the Pear River Delta, was one of the locations where the most outbreaks occurred (13/36, 36.1%), and outbreaks in the seaboard city of Zhanjiang also accounted for 25% (9/36) of V. parahaemolyticus outbreaks.
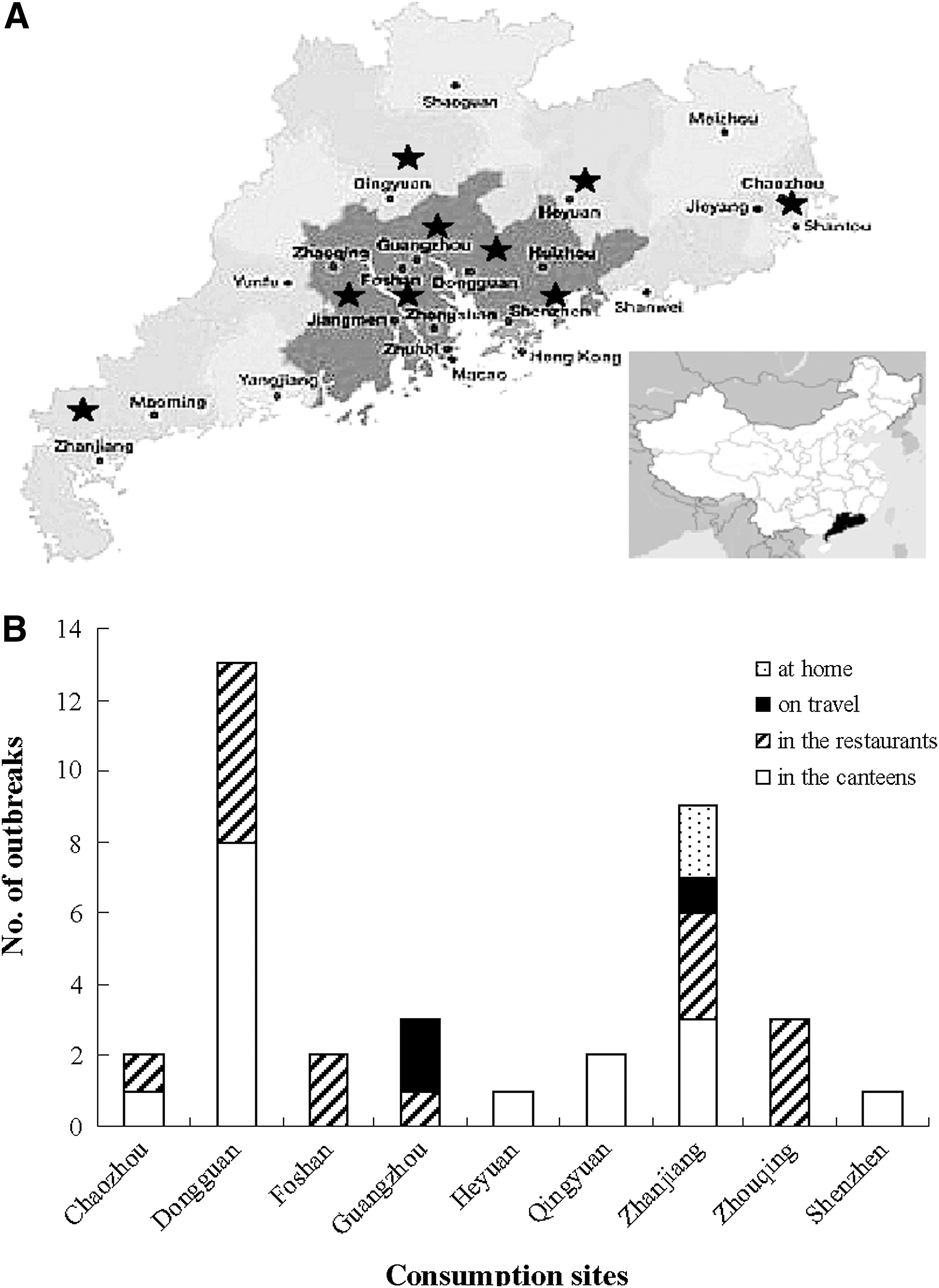
Spatial distribution of 36 Vibrio parahaemolyticus outbreaks during 2008–2010 in Guangdong Province.
Temporal Distribution
In terms of temporal distribution, 88.9% (32/36) of outbreaks appeared in the months from June to September, which were always the hottest months in Guangdong with annual average temperature ranged between 27.0°C and 28.8°C (Fig. 2). Outbreaks increased during these days as temperatures rose, suggesting that occurrence of V. parahaemolyticus outbreaks peak during summer.
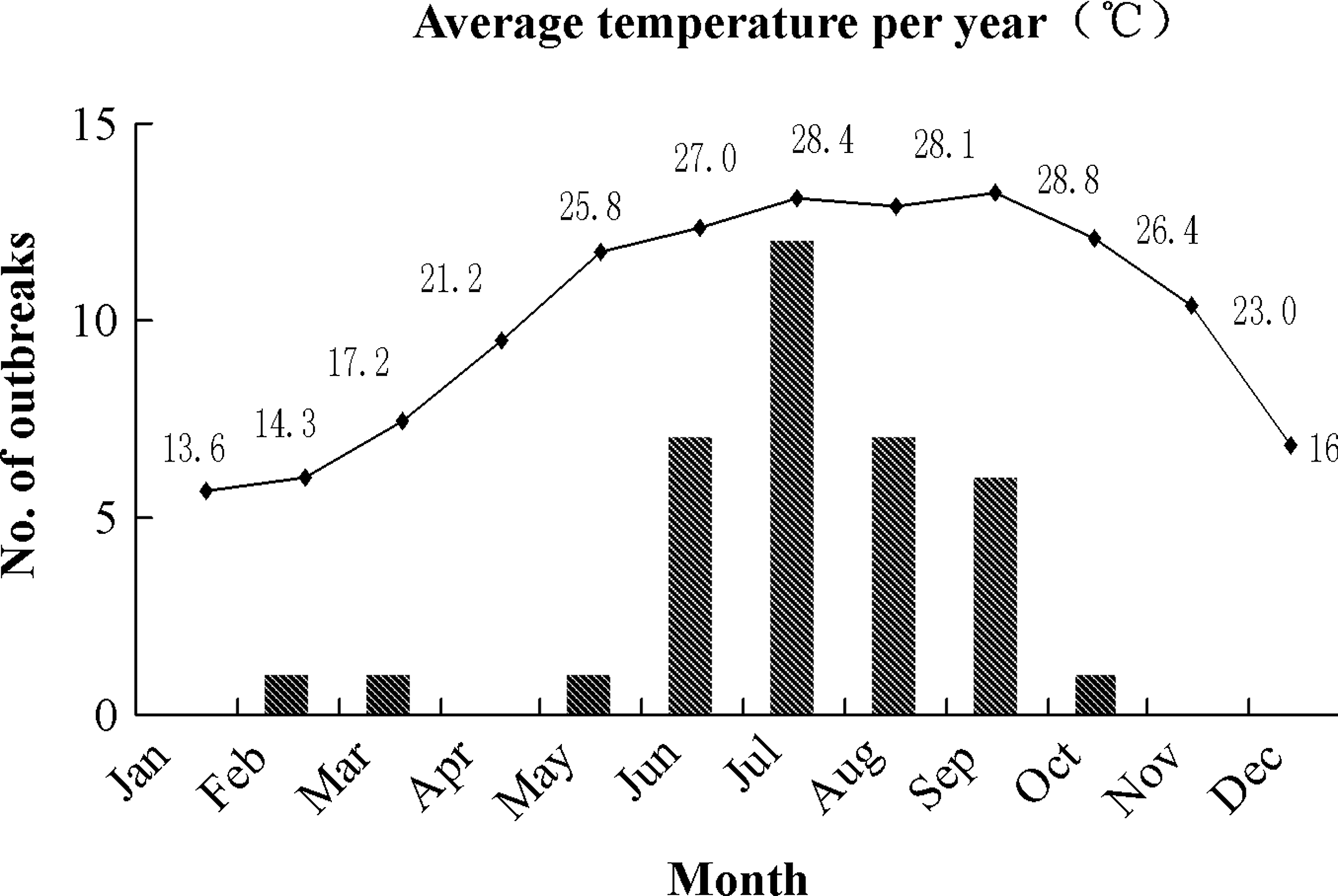
Temporal distribution of 36 Vibrio parahaemolyticus outbreaks during 2008–2010 in Guangdong Province. Statistical analysis of the number of outbreaks that occurred in different months was conducted by histogram. The average temperature during these months is shown in parallel by line chart (Data source: website of Guangdong Meteorological,
Distribution of serovars
It was demonstrated that 31% (11/36) of V. parahaemolyticus outbreaks were caused by a single serovar of strains, whereas 69% (25/36) were caused by two or more serovars (Table 2). A total of 284 strains were characterized into 22 serovars, and the majority of serovars included O3:K6 (51.4%, 146/284), O4:K8 (16.9%, 48/284), O1:Kut (8.5%, 24/284), and O2:K3 (4.6%, 13/284). O3:K6 and O4:K8 seemed to be the most dominant serovars in Guangdong (Fig. 3 and Table 2). It was suggested that 68.1% (175/257) of strains from patients and 37.0% (10/27) from food vehicles belonged to these serovars.
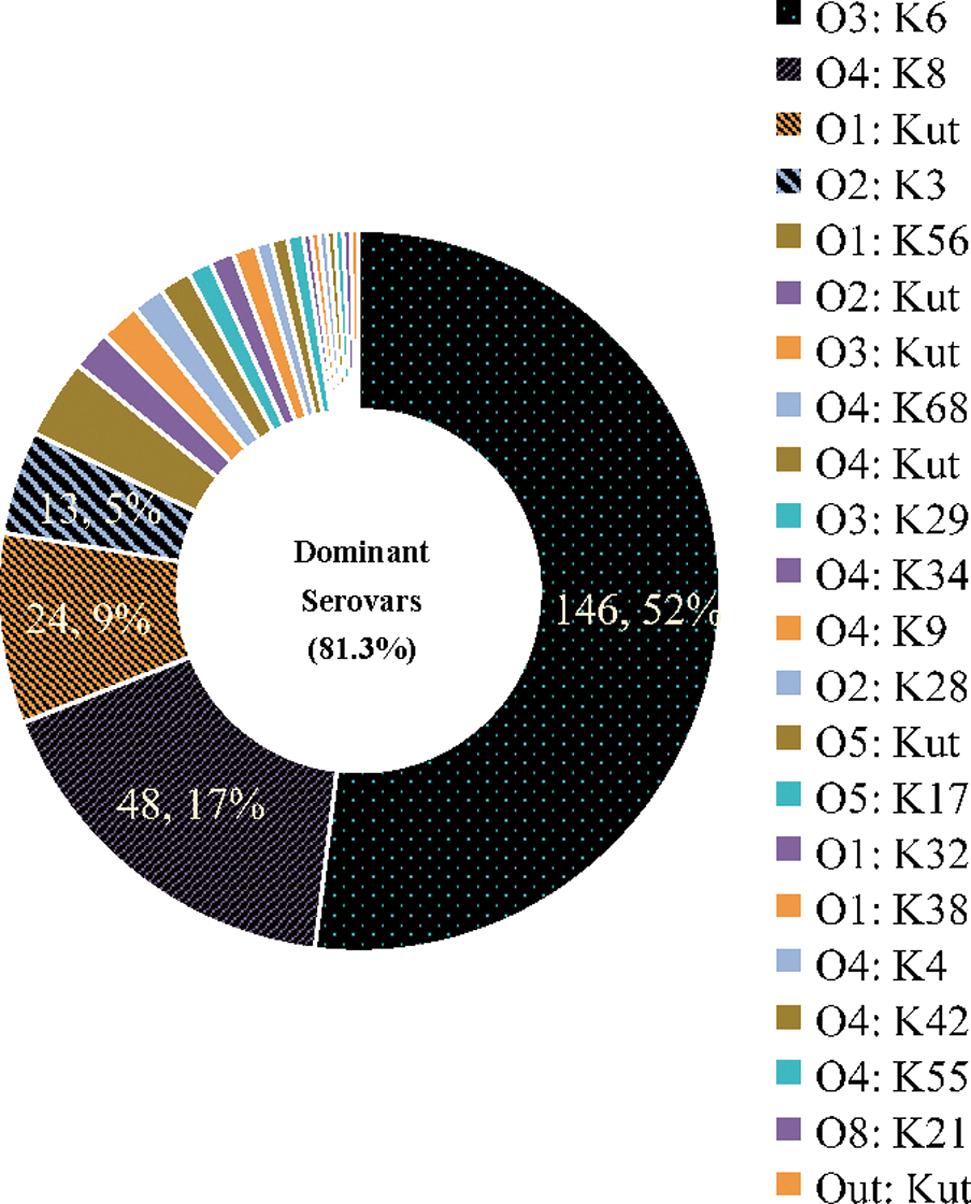
Distribution of serovars of Vibrio parahaemolyticus strains isolated in 36 outbreaks during 2008–2010 in Guangdong Province. A total of 284 strains were distributed in 22 serovars and demonstrated by pie chart (numbers of strains, proportion), the streak parts of which represent the strains belonging to the four dominant serovars that are displayed as numbers and percentage.
Virulence genes detection
PCR amplification results showed that 93.0% (264/284) of strains were tdh-present and trh-absent, whereas only one strain (0.3%) was trh-present and tdh-absent. Also, 98.8% (254/257) of clinical strains were tdh-present and trh-absent, whereas only two lacked both markers and one was trh-present and tdh-absent. As opposed to clinical strains, 37.0% (10/27) of strains from food vehicles were tdh-present and trh-absent, whereas the remaining strains lacked both virulence genes.
PFGE subtyping
The homology of a total of 66 strains from four typical outbreaks represented by a diversity of serovars, spatial, and temporal distribution were conducted by means of cluster analysis (Fig. 4), including outbreak of No. 31 caused by a single serovar of strains (Fig. 4A), No. 16 caused by multiserovars (Fig. 4B), and No. 8 and No. 22 included strains from patients and a food vehicle (Fig. 4C and D). Generally, the PFGE patterns of strains from the same outbreak were distinguishable. Serovars of strains from outbreaks No. 31 were identical (O1: Kut), but the PFGE patterns could be divided into two distinct clusters, the similarity of which was only 59%, indicating that these O1:Kut strains were genetically diversified (Fig. 4A). Analysis of No. 16 (Fig. 4B) demonstrated that an O4:K8 (strain number: F09-72) and two O4:K68 were closely related to all the O3:K6 strains (the similarity within these strains was 80.9%), which were distinctly different from the major cluster that was composed of other O4:K8 and O3:K29 strains (the similarity was 56.5%). These O4:K8, O3:K6, and O4:K68 strains possessed indistinguishable patterns that were closely genetically related to each other. The patterns of strains from patients belonging to O1:Kut in No. 8 (Fig. 4C) demonstrated 100% similarity with the O3:Kut from stewed colon of pigs. Another O4:K8 strains from fish was also 100% similarity with strains from another two patients. These results might indicate that several strains from food and patients with different serovars seemed to be closely correlated and those dishes were food vehicles that probably were associated with infections. In addition, we first confirmed cross-contamination between vehicles of salted food and seafood by molecular subtyping. The PFGE patterns of O2:Kut strains from stewed colon shared 100% similarity with the strains from mussels along with several O2:Kut from patients in outbreak No. 22 (Fig. 4D).
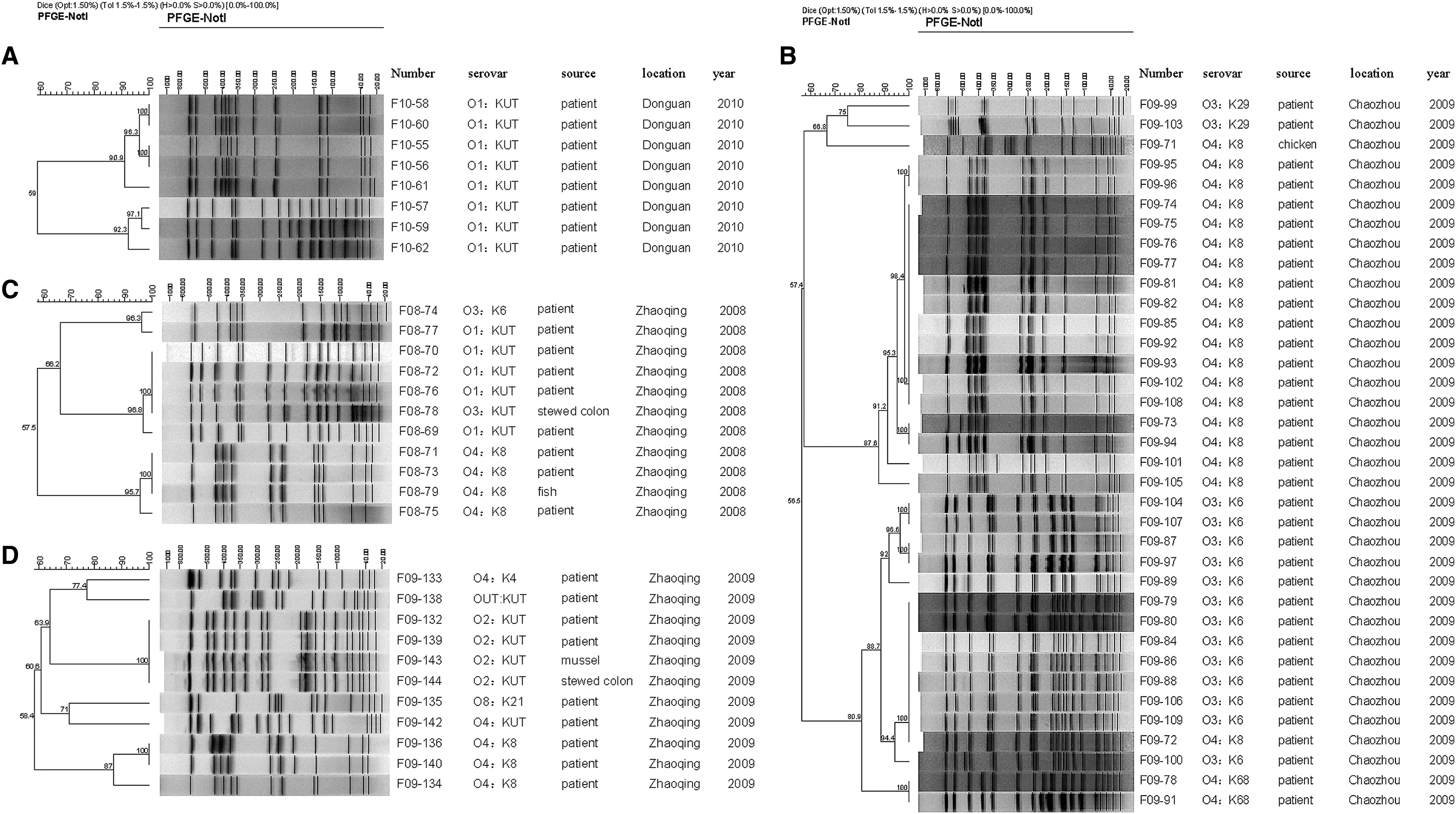
Cluster analysis of pulsed-field gel electrophoresis (PFGE) patterns of Vibrio parahaemolyticus strains from four typical outbreaks that occurred during 2008–2010. Dendrograms were constructed by the unweighted-pair group method with arithmetic mean clustering based on the genetic similarity coefficient (Dice). PFGE patterns and relative information on 66 strains isolated in outbreaks No. 31, No. 16, No. 8, and No. 22 are shown in panels A–D, respectively.
Discussion and Conclusions
There had been many reports about raw products cross-contaminated with cooked seafood in V. parahaemolyticus outbreaks (WHO, FOS, SDE, 2009). In this study, we noticed that the majority of cause-confirmed outbreaks were associated with various types of seafood and also salted food. The salted food (including viscera of pigs and cattle, meat, and vegetable salad) seemed to be a vital risk factor for resulting in V. parahaemolyticus infections as determined from our findings; these factors were rarely reported previously. The molecular evidence of strains within food vehicles indicated that cross-contamination between salted food and seafood might exist in these outbreaks. Since salted food is a common part of the diet in Guangdong Province, the findings were not too surprising but the risk was alarming. In addition, V. parahaemolyticus outbreaks were seasonally distributed with a high incidence in summer, and usually emerged in canteens and restaurants, especially in cities with a large population. Therefore, continuous supervision of food processing, including preparation and storage and risk assessment of V. parahaemolyticus in salted food, is urgently needed.
Serotyping results demonstrated that most V. parahaemolyticus outbreaks were caused by multiserovars of strains, mainly including O3:K6, O4:K8, O1:Kut, and O2:K3 serovars, which was basically consistent with the previous studies besides O4:K8 (Chiho et al., 2000; Jaime et al., 2004; Oralak et al., 2007; Ansaruzzaman et al., 2008; Harth et al., 2009). O3:K6 was also the dominant serovar of V. parahaemolyticus, resulting in outbreaks in Guangdong. It is noteworthy that O4:K8 was also a significant serovar that resulted in several outbreaks locally.
Pathogenic V. parahaemolyticus strains that had tdh and/or trh virulence genes present were considered to mainly appear in clinical strains, whereas these strains seldom existed in those from the environment (Yeung et al., 2004; Yang et al., 2008). In this study, the percentage of pathogenic strains from patients (98.5%) was similar to that in previous reports (USFDA, 2005; Ma et al., 2009). Surprisingly, we found that the percentage of pathogenic strains from food vehicles (37.0%, 10/27) were significantly higher than that of strains from seafood (3.78%, 2/53), which we reported before (Ma et al., 2009), suggesting that occurrence of pathogenic strains from food vehicles in outbreaks seemed to be more frequent than those isolated from marine products. There were studies reporting that tdh-absent isolates in the environment may transfer to the tdh-present strain through lateral gene transfer phenomenon (LGT) for acquiring and integrating fragments of tdh gene by functional insertion (Kamruzzaman et al., 2008a, b). Whether there were factors leading to the differences between pathogenic strains from food vehicles and environment through LGT were still unknown.
PFGE is a sensible means of molecular typing for distinguishing strains from V. parahaemolyticus outbreaks with identical serovars. In this study, 66 isolates from four typical outbreaks were subtyped. Considering a more detailed discussion, other similar characteristics of outbreaks have not been demonstrated in this article. The subtyping results indicated that strains with an identical serovar in the same outbreak showed different PFGE patterns that were genetically diversified. We also noticed that different serovars of strains possessed the same pattern, which might belong to the pandemic colonies that possess the characteristics of positive results of group-specific PCR (GS-PCR) amplification and share a high degree of similarity of molecular patterns within various serotypes (Chiho et al., 2000; Khamphouth et al., 2008; Hatsumtsuyai et al., 2000). For further analysis, strains with highly similar PFGE patterns could be confirmed by GS-PCR. In addition, we only conducted the NotI-PFGE method instead of double-enzyme digestion to analyze the genetic relatedness of strains, because they were obtained from a certain outbreak with clear epidemiology correlation instead of from several sporadic cases with unknown relevance. As a preferred method for subtyping, we were convinced that NotI-PFGE possessed a sufficient capacity for differentiating the strains with the same serovar, such as O3:K6 and O4:K8 in this study.
Above all, due to scarcity of systematic surveillance to determine risk factors and sources, V. parahaemolyticus infections are still a severe public health issue globally and locally (Yeung et al., 2004; Ma et al., 2009). CDCs of Guangdong province should pay more attention to food surveillance, especially for salted food including viscera of pigs and cattle, meat, and vegetable salad. PFGE typing for homogeneity analysis of isolates from sporadic infections and outbreaks as well as epidemiology investigation of food vehicles should also be carried out promptly to explore the contamination sources and control the incidence trend.
Footnotes
Acknowledgments
We thank the CDCs of Chaozhou, Dongguan, Foshan, Guangzhou, Heyuan, Qingyuan, Shenzhen, Zhanjiang, and Zhaoqing in Guangdong province for supporting the strains isolation and for providing epidemiology information, and the Institute of Nutrition and Food Safety of GDCDC for providing a statistics assistant. This study is funded by the Research Center for Health Emergency Responding Technology of Food Safety in Guangdong Province (YueKeHanSheZi[2011] No. 733); Preservation and utilization on germ-plasm resources of pathogenic microorganisms in Guangdong province (2012B060400012); the major projects of the National Science and Technology: Study on epidemiology characteristics of pathogens on infectious disease in Guangdong Province (2012ZX10004213-004); Medical Science and Technology Research Fund (A2010061), Guangdong Province; and Guangdong Provincial Natural Science Foundation Team Project (06201654).
Disclosure Statement
No competing financial interests exist.