
Abstract
The detection of pathogenic Vibrio in seafood from Senegal has generated five food alerts in the European Union. To investigate the presence and abundance Vibrio cholerae and Vibrio parahaemolyticus in seafood and coastal and estuarine waters, 123 seafood samples and 52 water samples were collected during 2007–2009 from two large seafood markets in Dakar, and from different oceanic and estuarine areas of the country. V. parahaemolyticus was detected in 30.1% of seafood samples, whereas presence of V. cholerae was only found in 1.6%. In water samples, V. parahaemolyticus and V. cholerae were detected in 28.8% and 5.7% of the samples, respectively. Abundance of V. parahaemolyticus in seafood from the fishing areas ranged from <0.3 to 7.5 most probable number (MPN) per gram. In samples from markets, densities of V. parahaemolyticus showed higher values ranging from 0.61 to >110 MPN/g. Densities of V. cholerae in the two positive seafood samples reached values of 0.36 and 0.61 MPN/g, repectively. V. parahaemolyticus strains were found to possess tlh, but not tdh and trh by polymerase chain reaction, and all the strains of V. cholerae were non-O1 or non-O139. These results suggest that the prevalence of high salinities in coastal and estuarine environments of Senegal limits the occurrence of V. parahaemolyticus and V. cholerae, despite warmer temperatures prevailing in seawater environments throughout the year. Furthermore, temperature abuse driven by a deficient cold chain over the distribution and retail sales may represent a major risk due to the postharvest multiplication of these Vibrio pathogens.
Introduction
Pathogenic bacteria belonging to the Vibrio genus are present or transitorily present (Vibrio cholerae) in seawater or in estuarine water worldwide and are implicated in numerous outbreaks of foodborne disease (David-Prince et al., 1980; Hlady et al., 1993; Kaper et al., 1995; DePaola et al., 2003). The most well-known and studied species are V. cholerae, V. parahaemolyticus, and V. vulnificus. Throughout the last 10 years, five food alerts have been declared that describe the association of pathogenic Vibrio with seafood harvested from coastal Senegalese waters and exported to the European Union (Bilan des alertes sanitaires/Europe de 1999 à 2010, Sénégal). In addition, from 1971 to the present time, the country has recorded several cholera epidemics. The last four affected some 50,000 people and at least 1000 deaths were reported. Epidemiological evidence suggested that contamination was due to poorly prepared foods and contaminated water (Diop and Coll-Seck, 1991; Sow et al., 1997; Aidara et al., 1998; Epidémie de choléra au Sénégal, 2006; Ndour et al., 2006). Bacterial isolates obtained during these epidemics often showed the coexistence of V. cholerae strains with pathogenic strains of V. parahaemolyticus (Miyamoto et al., 1969; Barker et al., 1974; Sow et al., 1997). Then, the question is: Do we have all the relevant data on the extent of this pathogen (V. parahaemolyticus) in the aqueous environment between epidemics?
Despite the growing emphasis on these pathogens in Senegal and the strategic importance of the fisheries sector, this country lacks of a regular surveillance program for pathogenic Vibrio to elucidate the relevance of the organisms in the marine environments.
To assess the incidence of Vibrio spp. in the environment and Senegalese seafood products, we carried out a surveillance study to investigate the presence, abundance, and pathogenicity of V. cholerae and V. parahaemolyticus in seafood, seawater, and estuarine waters. This was done in order to help the authorities make the best decisions in terms of management strategies of risks related to these pathogenic strains in relation to the exportation of Senegalese seafood products.
Materials and Methods
Sample collection
The study was carried out from May 2007 to December 2009. A total of 175 samples originated from the environment and seafood were investigated.
Environmental collection
Environmental samples were collected from estuarine and sea areas:
Estuarine water collection
Thirty-six (36) estuarine water samples were collected in Pointe Saint Georges (medium salinity), Ziguinchor (high salinity), and Goudomp (very high salinity) as shown in Figure 1.
The figure shows the location of the different sampling points along the Senegalese coast. Senegal: With 700 km of coast, Senegal is the westernmost region of the African continent as part of the Guinea Gulf. Saint-Louis, Dakar, Mbour, Joal, Djiffer, Kafountine, and Ziguinchor are the sampling points selected in the study. They are among the most important fishing areas in the country. Ziguinchor is the estuarine area with more than 200 km of length. In this area, we have three sampling points: Point Saint Georges, Ziguinchor, and Goudomp.
Seawater collection
Sixteen (16) seawater samples were collected along the coast from North to South (Fig. 1) during 4 months: November 2008, February 2009, March 2009, and April 2009. About 1 L per sample was collected using a sterilized bottle and placed in an icebox, and all samples were cooled with ice during transport (16 h by boat) and analyzed within 48 h of collection at the Amerger Casamance laboratory in Dakar.
Estuarine shrimps collection
Seventy-two (72) shrimp samples were collected in the estuarine area of Pointe Saint Georges, Ziguinchor, and Goudomp as shown in Fig. 1. Approximately 500 g per sample were collected from two different shrimpers in each site, from May 2007 to April 2008.
Seafood collection
Fifty one (51) seafood samples were randomly purchased from retailers in two markets of Dakar, every 2 months from June 2008 to December 2009: shrimps (27 samples), mussels (8), oysters (2), cephalopods (9), and flat fish (5). Samples were placed in an icebox and cooled with ice during transport (between 5 and 16 h by car or boat) from the harvest sites to the markets.
Temperature and salinity determination
Bottom water temperature and salinity were determined monthly at each site by using a Cond 330i/340i combination thermometer and salinometer (WTW 82362 Wellheim, Germany).
Microbiological analysis
Sample preparation
Two hundred milliliter aliquots of each water sample were concentrated by filtration through a 0.45-μm-pore-size filter (Sartorius, Germany) using a vacuum pump, and filters were subsequently placed into 225 mL of Alkaline Peptone water (APW). Filters in APW were homogenized for 60 s using a stomacher (Humeau Laboratoires, France) and then the samples were incubated at 37°C for 16–18 h.
Samples of shrimp, shellfish, cephalopod, and flat fish were analyzed using the most probable number (MPN) technique according to the procedure described by Blanco-Abad et al. (2009), except that APW and salt polymyxin broth were not analyzed by polymerase chain reaction (PCR); in addition, respective presumptive isolates were gram stained, tested for oxidase activity, and identified using the API 20E biochemical testing kit (Biomerieux, France) (Fig. 2).
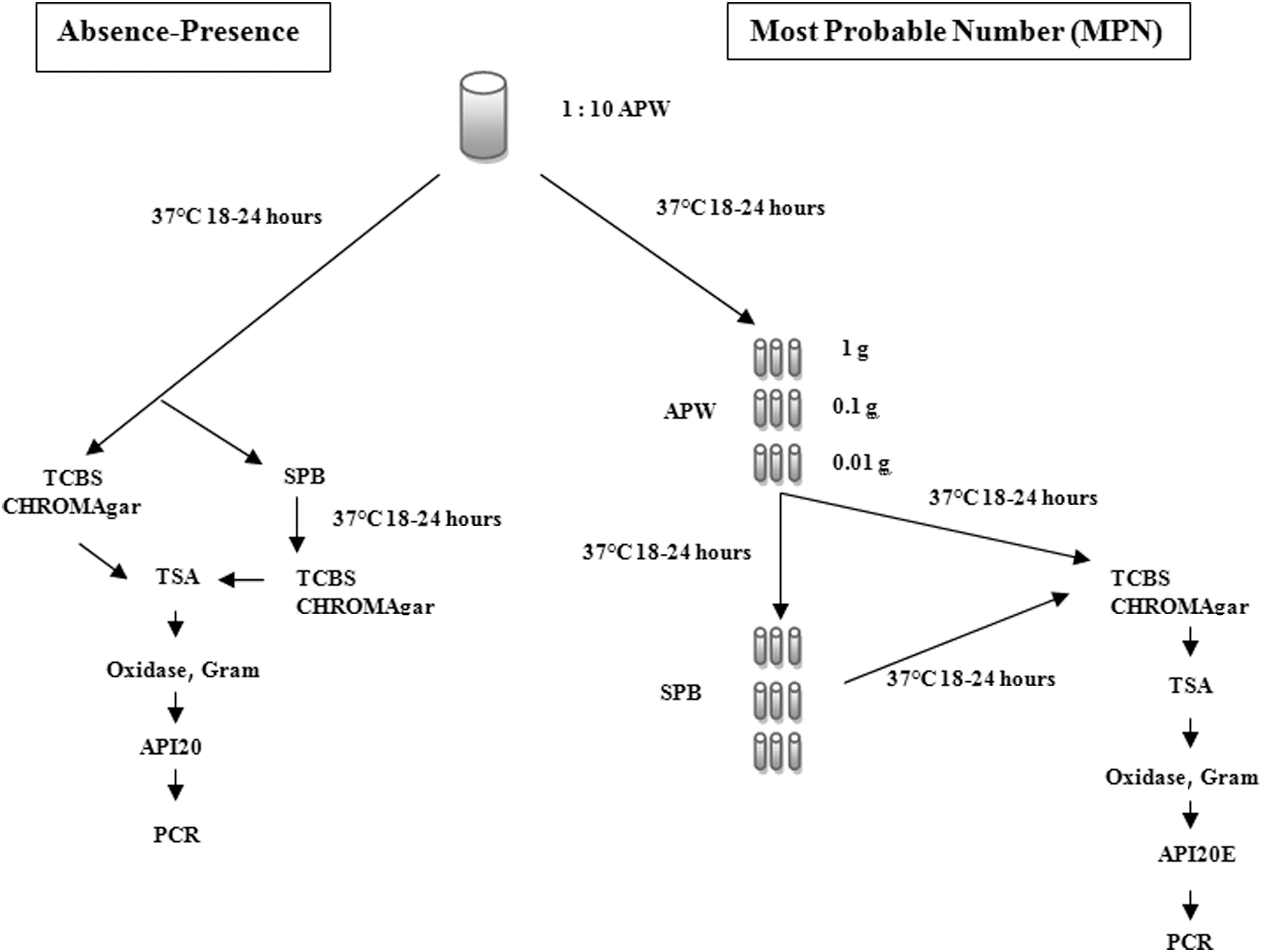
Presence–absence method: Samples of water were analyzed according the ISO 8914 standard method (ISO 8914, 1990). Most Probable Number (MPN) method: Samples of shrimp, shellfish, cephalopod, and flat fish were analyzed using the MPN technique according the procedure described by Blanco-Abad et al. (2009). APW, alkaline peptone water; TCBS, thiosulfate-citrate-bile salts-sucrose agar; CHROMAgar Vibrio uses chromogenic technology to allow for the isolation and detection of V. parahaemolyticus, V. vulnificus, V. cholerae, and V. alginolyticus. It is a selective and differential agar. TSA (2% NaCl): tryticase soy agar; SPB, salt polymyxin B broth; Oxidase, gram, Presumptive tests before biochemical identification; API20E, a biochemical micromethod identification; PCR, polymerase chain reaction.
Samples of seafood, except shellfish, were cut into several portions. Twenty-five grams (25 g) of each sample were transferred into sterile plastic bags with 225 mL of APW to obtain a 1:10 dilution and subjected to further homogenization for 60 s using a stomacher. Fifteen (15) specimens of each sample of mussel and oyster were washed and scrubbed under running potable water and opened aseptically with a sterilized scalpel. Meat and shell liquid were collected in a sterile jar, cut with scissors, and mixed thoroughly. Fifty grams (50 g) of liquid and meat were transferred into sterile plastic bags with 450 mL of APW to obtain a 1:10 dilution and subjected to further homogenization for 60 s using a stomacher.
A total of 10 mL of the 1:10 APW dilution, and 1 mL of 1:10 and 1:100 dilutions were inoculated in the three tubes of three MPN series and were incubated at 37°C for 18–24 h. Aliquots of 0.1 mL of APW broths were transferred to tubes containing 10 mL of Salt Polymyxin B Broth and incubated at 37°C for 18–24 h.
Plating onto thiosulfate citrate bile salts sucrose (TCBS) and CHROMagar Vibrio
After incubation of the MPN dilutions, a loopful of each culture was streaked onto TCBS and CHROMagar Vibrio medium, followed by incubation for 24 h at 37°C. Five presumptive colonies of V. parahaemolyticus (green colonies on TCBS and violet colonies on CHROMagar Vibrio) and V. cholerae (yellow colonies on TCBS and blue colonies on CHROMagar Vibrio) were selected at random from each plate, streaked onto Trypticase Soy Agar (TSA) containing 2% NaCl (Difco), and incubated at 37°C for 24 h before performing presumptive tests (oxidase and gram stain). The identities of oxidase-positive, gram-negative colonies were confirmed as V. parahaemolyticus or V. cholerae using API20E strips, PCR analysis, and serological tests (V. cholerae).
Determination of O antigens of V. cholerae
Serological identification of V. cholerae isolates taken from TSA 2% NaCl agar plates were emulsified in a sterile saline solution (0.85% NaCl), and equal volumes of cell suspensions and agglutinating anti-O1 and anti-O139 sera was performed according to the manufacturer instructions (Bio-Rad, Marnes la Coquette, France).
Molecular characterization of V. parahaemolyticus
Isolates of V. parahaemolyticus obtained from API20E were analyzed by PCR for the presence of tlh, tdh, and trh genes.
DNA extraction
Colonies of V. parahaemolyticus obtained from TSA 2% NaCl were incubated at 37°C in a Luria-Bertani broth 2% NaCl overnight under agitation. After incubation, 1 mL of the culture was centrifuged at 10,000×g for 7 min. The supernatant was discarded and the resultant pellet was resuspended in 300 μL 1xTE buffer (10 mM Tris pH 8.0, 1 mM EDTA) and heated 100°C for 10 min. The lysate was centrifuged at 13,000×g for 5 min and the supernatant containing the DNA was transferred to a new tube and stored at −20°C until its use for PCR analysis.
PCR analysis
Detection of tlh, tdh, and trh genes was performed as described by Bej et al. (1999). The selected primers were as follows: tlh (5'-AAAGCGGATTATGCAGAAGCACTG -3', and 5'-GCTACTTTCTAGCATTTTCTCTGC-3'); tdh (5'-GTAAAGGTCTCTGACTTTTGGAC-3', and 5'-TGGAATAGAACCTTCATCTTCACC-3'); trh (5-TTGGCTTCGATATTTTCAGTATCT-3', and 5'-CATAACAAACATATGCCCATTTCCG -3'), with 50 μL of reaction mixture consisting of Taq polymerase buffer, 200 μM deoxynucleotide triphosphates, 50 pmol of each primer (Proligo, Paris, France), 1 U of Taq DNA polymerase, and 25 ng of DNA. PCR amplification was performed in a 2720 thermal cycler (Applied Biosystems, Netherlands) by using the following PCR parameters: predenaturation at 94°C for 5 min, followed by 35 cycles of 94°C for 1 min, 57°C (trh) or 58°C (tl, tdh) for 1 min, and 72°C for 1 min, with a final extension of 72°C for 10 min. Amplified DNA products were resolved by conventional electrophoresis in ethidium bromide-stained agarose gels.
The ATCC43996 (tdh+, trh−) and AQ4037 (trh+, tdh−) reference strains were used as positive controls in all biochemical tests and PCR assays.
Data analysis
The figures were carried out using the Excel histogram function of Microsoft Windows and the plot function of the statistic software R.
Results
A total of 123 crustacean and fish samples (99 of shrimp, 10 of shellfish, nine of cephalopod, and five of flat fish) were analyzed for the presence of Vibrio spp., and the salinity was determined for 36 estuarine water and 16 seawater samples, respectively. All water samples were analyzed for the presence of Vibrio spp. The different sampling points are shown in Figure 1.
Of the 175 samples analyzed, V. parahaemolyticus and V. cholerae were detected in 30.1% and 1.6% of these, respectively. All the V. cholerae isolates were determined to be non-O1/non-O139 or nonagglutinating V. cholerae. As shown in Table 1, V. parahaemolyticus and V. cholerae were detected in estuarine and seawater samples at higher incidence rates (28.8% and 5.7%, respectively). The levels of V. parahaemolyticus in seafood harvested from the estuarine area were low (<0.3 to 7.5 MPN/g), despite the existence of warmer waters in the area (from 22.8°C to 32.4°C). Water salinity in this area ranged from 34.5 to 63.1 g/L (Table 2).
S, Sokone; M-J, Mbour-Joal; D, Dakar; Gd, Goudomp; S-L, Saint-Louis; Ga, Gandiaye; K, Kafountine; Dj, Djiffer; Z, Ziguinchor; P, Pointe Saint Georges.
NAG, nonagglutinable.
P, Pointe Saint Georges; Z, Ziguinchor; Gd, Goudomp.
MPN, Most Probable Number; V. para., Vibrio parahaemolyticus; V. ch., Vibrio cholerae.
In seafood market samples, densities of V. parahaemolyticus ranged from 0.61 to >110 MPNg-1 and market product temperatures ranged from 13.5°C to 25°C, despite the transport from the fishing area on ice. The air temperature during the sampling period in these two seafood markets ranged from 17°C to 31.6°C. Densities of V. cholerae in the two positive seafood samples reached 0.36 and 0.61 MPN/g and are shown in Table 3. Data on ambient temperature and samples are mentioned in Table 3. Strains of V. cholerae were isolated only in samples from the markets. Data on water temperature and salinity at the sites of product origin were not available. All of the V. parahaemolyticus were tlh-positive. However, tdh and trh genes were absent, suggesting that Senegalese strains of V. parahaemolyticus may not possess these virulence factor genes.
The salinities recorded in the seawater samples ranged from 34.1 to 35.3 g/L, whereas in estuarine areas the salinities ranged from 37.4 to 63.1 g/L (Tables 2 and 4).
Interestingly, seafood samples positive for V. cholerae came from fishing areas where the salinity ranged from 34.1 to 35.3 g/L and are represented by two seafood samples each from Dakar and Kafountine and three seawater samples from Saint Louis, Mbour, and Joal, respectively (Table 1).
Discussion
This study's main objective was to provide baseline information for a seafood surveillance program to help the Senegalese public health authorities make prudent decisions and policy strategies for the management of risks associated with the exportation of Senegalese seafood products.
This study was in direct response to two very important facts. First, the fishing sector is one of the leading business activities in Senegal (ANSD/NACE-Sénégal, 2007, 2008, 2009, 2010, 2011). Second, detection of pathogenic Vibrio in seafood elaborated in facilities in Senegal has generated five food alerts in the European (Bilan des alertes sanitaires/Europe de 1999 à 2010, Sénégal). In this work, 123 samples of seafood products and 52 water samples were investigated for the presence of V. cholerae and V. parahaemolyticus. The percentage of V. parahaemolyticus and V. cholerae positive seafood samples were 30% and 2% (of 123 samples), and 28% and 5.7% of 52 water samples, respectively. These results and the previous food alerts indicate the need to identify additional virulence markers to improve risk assessment tools for the detection of these pathogens, as has been suggested by Paranjpye et al. (2012).
However, the abundance of total V. parahaemolyticus in samples from estuarine area was low in general and was affected more by salinity than by temperature (Figs. 3 and 4). This situation was illustrated by two different arguments:
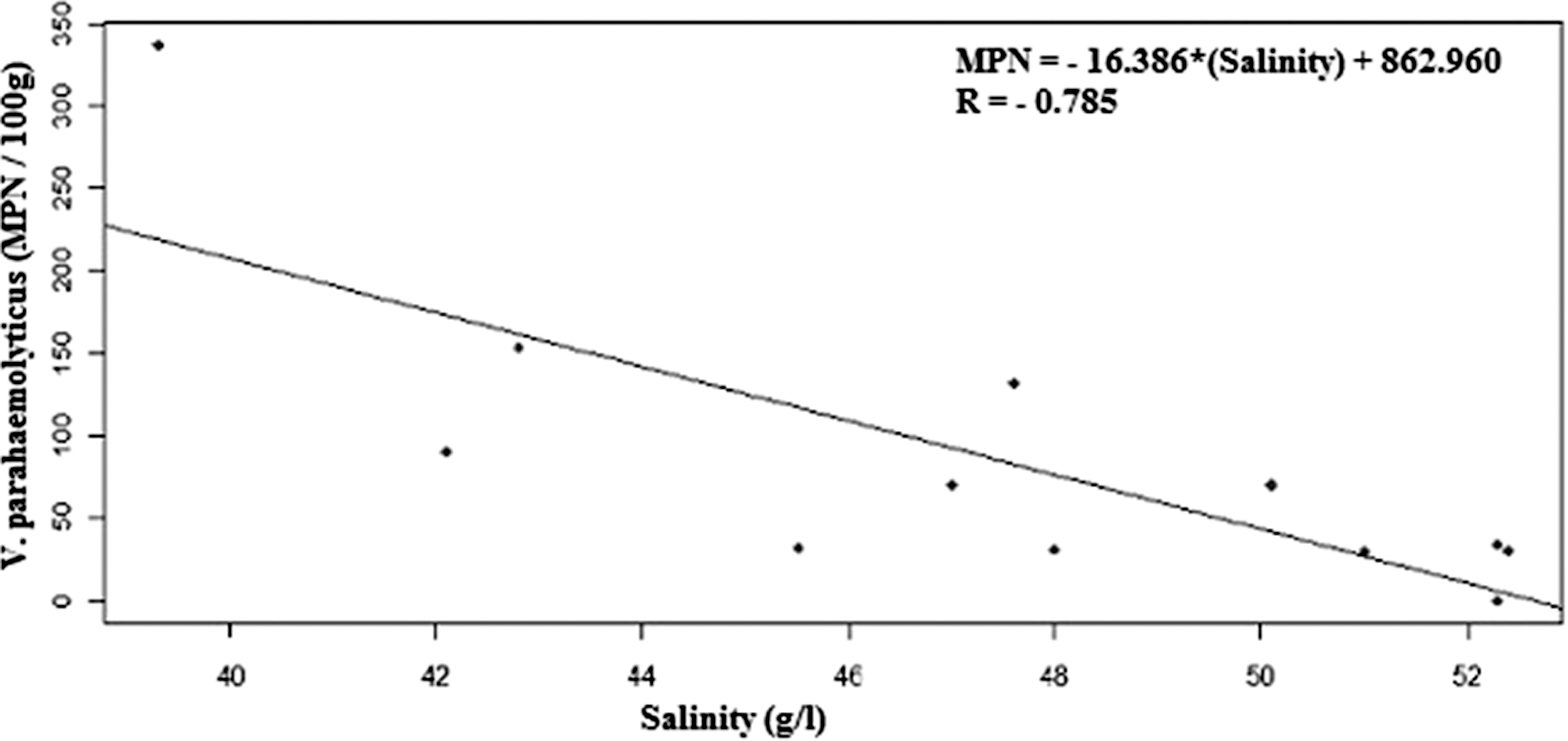
The figure shows the relationship between water salinity and Vibrio parahaemolyticus density in shrimp from the estuarine area in Ziguinchor. The figure was carried out using the plot function of the statistic software R that gives the equation Most Probable Number (MPN)=−16.386*(salinity)+862.960 and the regression value R=−0.785.
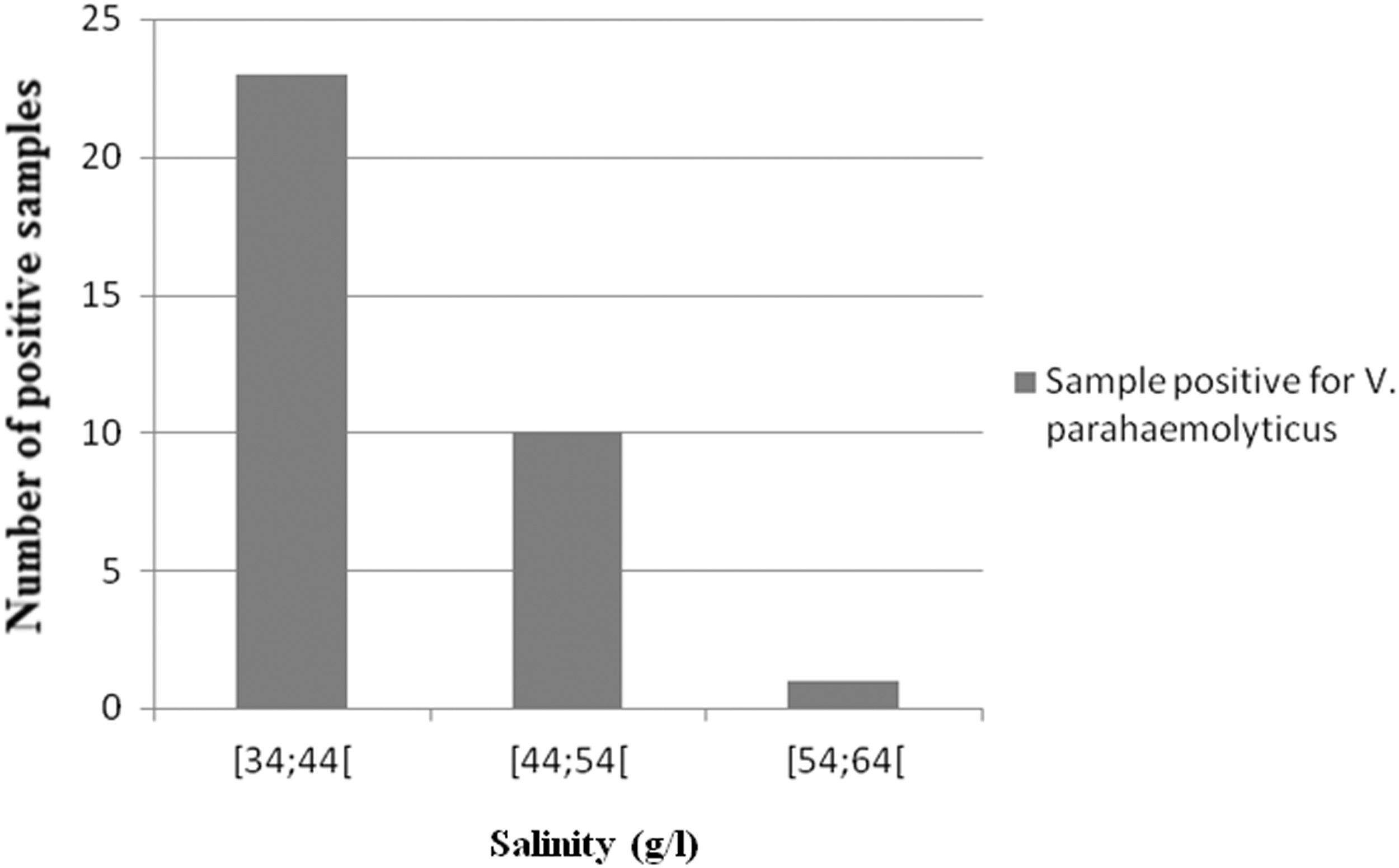
The figure was carried out using the Excel histogram function of Microsoft Windows. The histogram bars give the number of positive samples for Vibrio parahaemolyticus in each class of salinity (with a number of positive samples falling dynamic from [34;44[ to [54;64[). The x-axis shows the class of salinity. The y-axis shows the number of samples positive for V. parahaemolyticus.
On the other hand, the densities of V. parahaemolyticus in samples from markets were higher than samples harvested from estuarine areas. From the harvest site to the markets, sea products were cooled with ice during transport so that their temperature was often below 10°C. The high densities of V. parahaemolyticus recorded in samples from markets were due to their exposure to the ambient temperature during the period of sale (Table 3). The Codex Committee on Food Hygiene considers adequate a temperature of 10°C or less to control the growth of Vibrio spp. in seafood (Commission du Codex Alimentarius, 2009; CAC/GL 73, 2010; WHO/FAO, 2011).
In Senegal, apart from the cholera epidemics, gastroenteritis cases often are not the subject of bacteriological analysis. Previous studies have shown that many gastroenteritis cases were due to Kanagawa-positive strains of V. parahaemolyticus (David-Prince et al., 1980; Schanddevyl et al., 1984; Sow et al., 1997; Soumaré et al., 2007). Obviously, during cholera outbreaks, all the gastroenteritis cases are the subject of bacteriological analysis supported by the government. These authors have shown that the gastroenteritis cases due to pathogenic strains of V. parahaemolyticus represented about 2% of the choleriform syndrome (Soumaré et al., 2007). The question is: Does the lack of surveillance in interepidemic periods mask the true clinical burden of many infections?
The deteriorating socioeconomic conditions in recent years in many developing countries has increased the number of people exposed to cholera (WHO—Wkly Epidemiol Rec, 2011, 2012; Talkington et al., 2011). The management of cholera outbreaks in Senegal has been made more complex by the annual religious events which millions of people attend, after which we noted a resurgence of the epidemic (
Several authors pointed out climate anomalies as major events that increased cholera oubreaks and other Vibrio diseases in many regions (Koelle et al., 2005; Sedas, 2007; Paz et al., 2007; Martinez-Urtaza et al., 2008; Paz, 2009; Baker-Austin et al., 2010; Martinez-Urtaza et al., 2010). However, since 2008 no cholera outbreak has been declared in Senegal in a context of general cholera expanding in Africa (CDC—Cholera outbreaks, 2010a–d, 2012; WHO—Wkly Epidemiol Rec, 2012).
Most of the environmental studies link the presence of pathogenic strains of V. parahaemolyticus with moderate salinities (<32 g/L) and high temperatures (Liston, 1980; Kelly and Stroh, 1988; DePaola et al., 2003). The prevalence of high salinities in coastal and estuarine environments of Senegal (from 34.1 to 63.1 g/L) limits the occurrence of V. parahaemolyticus and V. cholerae. The low incidence of V. cholerae in seafood products (1.6%) and in water samples (5.7%) is due to the very weak survival capacity of this microorganism in the coastal and estuarine environments studied (salinity from 34.1 to 63.1 g/L). However, more research would be needed to elucidate the ecological factors governing the pattern of presence and population dynamics of pathogenic and nonpathogenic Vibrios in the area.
The results obtained in this study show that it is important that the Senegalese public health authority set up a surveillance and seafood control plan for the following reasons: • to avoid the ocurrence of food alerts, which can severely affect the exportation of seafood products; • to avoid a serious health problem of foodborne disease in Europe after the consumption of contaminated seafood products originating from Senegal; and • to avoid a health problem of foodborne disease in Senegal after the consumption of seafood sold in the markets by the local population.
At the local level, the products sold at the market must be rigorously controlled and submitted to regulation.
The control should be focused around the following points: 1. Verification of the catch size; 2. Imposition of the early rapid freezing of capture and strict observation of cold chain maintenance; 3. Guarantee that the vehicles and boats that are used to carry products must maintain hygiene; 4. The unloading and handling of the product in markets must be under the supervision of a veterinarian or any other person who can authorize its commercialization because the safety and hygiene criteria; 5. Prohibition of ambient temperature exposure of the product during its sale; 6. The training of all actors involved in the sector; and 7. The setting up of a code to help in the standardization of good harvesting, processing, and retailing practices across the food chain.
Footnotes
Disclosure Statement
No competing financial interests exist.