
Abstract
The interaction between the clinical isolate of enteropathogenic Escherichia coli (EPEC) SBANU8 and pea sprouts was compared with avirulent K 12. E. coli. This was carried out by repeated co-incubation with pea sprouts for 5 days, and the protein profile of the culture supernatant was analyzed by single and two-dimensional electrophoresis. Mass spectrometry analysis led to the identification of two serine protease inhibitors including a Bowman-Birk–type protein secreted by pea sprouts in response to clinical isolate. Expression of the E. coli intimin gene involved in animal host colonization and virulence was studied by reverse transcription polymerase chain reaction. Expression of this gene was high in SBANU8 when co-incubated with pea sprouts. The present study gives baseline data on the molecular level interactions of EPEC and pea sprouts, which are needed to design the outbreak control strategies.
Introduction
Consumption of sprouts has been identified as one of the potential causes of foodborne illness due to pathogen growth during the sprouting process. Pathogens presenting either in the interior of the seed or on the surface are favored for multiplication by the sprouting process (NACMF, 1999). Although contaminated seeds are the major vehicle for sprout-associated outbreaks, practices and conditions at sprouting facilities and handling by consumers may also impact safety of the sprouts. Massive outbreaks of E. coli O157:H7 in Japan (Michino et al., 1999), and of O104:H4 in Germany (Buchholz et al., 2011) and France (European Food Safety Authority, 2011) all were associated with sprouts. The majority of these outbreaks were linked to alfalfa, mung bean, clover, radish, mustard, cress, and fenugreek sprouts (Bang et al., 2011; Buchholz et al., 2011).
However, there is little research to date pertaining to the colonization of pea sprouts, which are commonly consumed in India. Moreover, most of the studies on sprout microbiology deal with detection and sanitation. Response of the plant host (the sprouts) to human pathogenic bacteria remains largely unknown. Insights into the molecular level interactions of foodborne bacteria and the seed sprouts are very much needed to redesign our outbreak control strategies for better containment. As an initial step toward the aforesaid approach, the response of pea sprouts to induced colonization by EPEC and E. coli K12 was studied.
Materials and Methods
Bacterial cultures
In a parallel study, we have been screening clinical isolates of E. coli from the diarrheal samples of children (kind gift from Dr. Rathinasamy Subashkumar, Kongunadu Arts and Science College, Coimbatore, India) for the presence of pathogenicity island genes (unpublished data). One of the isolates, SBANU8, which was shown to be positive to the marker genes of pathogenicity island I carrying TTSS and which also was recorded positive to salt aggregation and hemolytic tests, was chosen for this study. A virulent E. coli, K 12 substr. MG1655 (MTCC 1586), was obtained from Microbial Type Culture Collection and Gene Bank Facility, Institute of Microbial Technology, Chandigarh, India and used for comparison. Growth on eosin methylene blue (EMB) agar and standard biochemical tests were performed for all the isolates.
Sprouting of pea seeds
Pea seeds (variety Ooty 1) obtained from Tamil Nadu Agricultural University, Coimbatore, India, were surface sterilized according to the procedure of Barak et al. (2005) with slight modification. Seeds were treated with CIO2 (HiMedia, India) solution (500 μg/mL) for 5 min, dried at 45°C for 24 h, and dry heated at 70°C for 24 h. The seeds were aseptically placed in sterile Petri dishes with two layers of sterile filter papers (Whatman No. 1) wetted with sterile water. The plates were left undisturbed at room temperature in darkness for 3 days.
Induced colonization of pea sprouts by E. coli
Co-incubation of bacterial cultures with pea sprouts was done as previously reported with modifications. About five pea sprouts were added per 3 mL overnight cultures of strains SBANU8 and k12 grown in nutrient broth (HiMedia, India) and incubation at 37°C was continued for 4 h. Cultures without addition of pea sprouts were maintained as a control. After 4 h, the sprouts were removed and the cultures were transferred aseptically to fresh sterile test tubes to which another set of fresh pea sprouts was added. The co-incubation was continued for another 4 h in a shaker at 37°C. After this step, 10 μL of the cultures were transferred aseptically to 3 mL sterile nutrient broth in test tubes and new sets of pea sprouts were added. This was incubated overnight at 37°C with shaking. The procedure was repeated for 5 continuous days (each day with two 4-h incubation and an overnight incubation with pea sprouts). At the end of the 5th day, the cultures were centrifuged at 8000 × g for 10 min at 4°C. The supernatant and bacterial pellet were used, respectively, for extraction of protein and RNA.
Protein extraction
The culture supernatants were filtered through 0.22-μm filters and an equal volume of 40% trichloroacetic acid (wt/vol) was added and incubated at −20°C for 1 h. The mixture was centrifuged at 10,000×g for 10 min at 4°C. The supernatant was discarded and the protein pellet was washed with ice-cold acetone (0.5 mL of ice-cold acetone was added dropwise and centrifuged at 10,000×g for 10 min). Two more washings were done using 80% acetone. The protein pellet was dissolved in 300 μL of phosphate-buffered saline and stored at −20°C.
Sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) analysis
Protein samples were quantified by Bradford assay (Bradford, 1976) and about 50 μg of each of the protein samples were analyzed in SDS-PAGE in 12% separating and 4% stacking gel using the procedure described (Laemmli, 1970). After electrophoresis, the gel was stained with Coomassie brilliant blue staining solution.
Two-dimensional (2D) gel electrophoresis
Based on results of SDS-PAGE, protein samples of SBANU8 isolate (both control and co-incubated with pea sprouts) were used for 2D gel electrophoretic analysis. Sample solution containing 100 μg of protein in 300 μL of rehydration buffer was used for passive rehydration of 17 cm pH 4–7 linear IPG strips (Hoefer Scientific, Holliston, MA). Isoelectric focusing (IEF) was performed using Hoefer IEF 100 unit programmed to provide an optimum, maximum field strength of 600 V/cm and a 50 μA limit/IPG strip. Initially, a low voltage of 250 V was applied for 15 min to remove salt ions and charged contaminants after which a linear ramping step was applied to reach 10,000 V in 3 h, at which point focusing took place at 10,000 V for 60,000 Vh. After IEF, the strips were held at 500 V. When focusing was complete, the strips were removed from the focusing tray and placed gel side up in a reswelling tray for storage at −20°C overnight.
Prior to second-dimension electrophoresis, the focused proteins in the strips were equilibrated to reduce and alkylate proteins as described (Yajima et al., 2004). The proteins were separated by SDS-PAGE in the second dimension using 12% polyacrylamide gels in a SE 640 vertical gel electrophoresis unit (Hoefer Scientific, Holliston, MA). Electrophoresis was carried out in 20×20 cm gels at 75 V overnight followed by 150 V for an additional 5 h. Gels were stained using Silver Stain Plus Kit (Bio-Rad, Hercules, CA) and scanned with a densitometer (GS-700, Bio-Rad). The 2D electrophoresis experiment was repeated twice with similar results. Protein spots were analyzed using Melanie III (Gen Bio, Geneva, Switzerland) software. The protein spots were excised carefully and subjected to matrix-assisted laser desorption ionization time-of-flight mass spectrometry (MALDI-TOF MS)/MS analysis at the Spectroscopy/Analytical Test Facility, Indian Institute of Science, Bangalore, India.
Data analysis
The MS/MS spectra were used to search against the NCBI nonredundant protein database using an MS/MS ion search engine, a computer software program (
RNA extraction and reverse transcription–polymerase chain reaction (RT-PCR)
Total RNA was isolated from a bacterial cell pellet using RaFlex Kit (Genei, Bangalore, India) as per kit instructions. Primers to amplify the intimin gene of E. coli (Gen Bank accession AF319597) were designed using Primer3 software and validated using primer-BLAST tool of NCBI. Primers were synthesized at Bioserve Biotechnologies India Private Limited, Hyderabad, India. The forward and reverse primers are 5’ATGGCAGTATTGACGGATAC3’ and 5’GAGACATCACATAATCCGCC3’. First-strand cDNA synthesis was carried out with 20 ng RNA, avian myeloblastosis virus (AMV) reverse transcriptase, and reverse primer using a cDNA Synthesis Kit (Genei, Bangalore, India) as per the kit instructions. The reaction mixture was incubated at 42°C for 1 h and terminated at 70°C for 10 min followed by chilling on ice. PCR was carried out using 5 μL of cDNA (synthesized from 20 ng RNA), 2 U Taq polymerase, 200 μM each dNTP, and 1.5 mM MgCl2. Primers were used at 0.2 μM final concentration. Amplification was done with initial denaturation at 95°C for 2 min followed by 30 cycles of 95°C for 1 min, 49°C for 1 min, and 72°C for 1 min. This was followed by a final extension step of 72°C for 7 min. The PCR products were analyzed in 1% agarose gel stained with ethidium bromide.
Results and Discussion
Expression of intimin gene by E. coli
We studied the expression of this eae gene in pathogenic E. coli compared to K12, both in the presence and absence of pea sprouts. The results of semiquantitative RT-PCR analysis are shown in Figure 1. The primers designed were targeted to amplify a 700-bp fragment of the 2.6-kb gene. The expression of this gene was not found in the K12 strain in both cases because the strain lacks the pathogenicity island carrying this gene. The presence of pea sprouts and induced colonization has increased the expression of this gene in SBANU8. Based on the observation, we propose that the eukaryotic host colonization strategy by E. coli (whether animal or plant) overlaps at least with expression of the intimin gene. Nevertheless, the possibility of expression of intimin in the process of an attempt of E. coli to colonize pea sprouts also cannot be overlooked. Studies on successful/unsuccessful colonization of E. coli based on comparison with other reported seed sprouts and analysis of molecular processes in the plant host after colonization are needed for a clearer understanding of the event.

Reverse transcription–polymerase chain reaction analysis of eae gene in Escherichia coli co-incubated with or without pea sprouts. Lane 1: 100–1000-bp DNA ladder; lane 2: K12 without pea sprouts; lane 3: SBANU8 strain without pea sprouts; lane 4: K12 with pea sprouts; lane 5: SBANU8 strain with pea sprouts.
Protein-level response of pea sprouts to E. coli colonization
When the SBANU8 strain expressed intimin during attachment or attempted attachment, the pea sprouts responded with secreted proteins into the interactome. The SDS-PAGE analysis of culture supernatants of the SBANU8 strain with and without pea sprouts is shown in Figure 2. The protein profile showed a prominent protein of 26-kDa size. This protein was not seen in K12 (with or without pea sprouts) as well as in SBANU8 in the absence of pea sprouts. At this stage, it was not clear whether the protein is exclusively produced by the SBANU8 strain in response to pea sprouts or by the pea sprouts in response to colonization by the strain.

Sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis of culture supernatants of Escherichia coli strains incubated with and without pea sprouts. Lane 1: protein molecular weight marker; lane 2: SBANU8 strain without pea sprouts; lane 3: SBANU8 strain with pea sprouts; lane 4: K12 without pea sprouts; lane 5: K12 with pea sprouts.
Pea sprouts secrete trypsin inhibitors in response to pathogenic E. coli
To further confirm and identify the protein we observed in SDS-PAGE, we performed 2D electrophoresis with the protein samples obtained from SBANU8. The images of 2D gels are shown in Figure 3. Fewer protein spots were found in the interactome. However, there were few proteins (spots 1, 2, and 3) that were unique and protein spot 4, which was down-regulated in the sample from SBANU8-pea sprout interaction. These four spots were subjected to mass spectrometry and two proteins (spots 1 and 3) turned out with a high ion score and peptide matching in the database search. One of the proteins (spot 1) was identified as a Bowman-Birk inhibitor since the peptides matched with the trypsin-binding domain of this protein in the database (Table 1). The other protein (spot 3) was also a trypsin inhibitor. Both hits matched with the respective proteins of animal origin, which may be understood as pointing out to the possible conserved domains in the serine protease inhibitors across life forms.
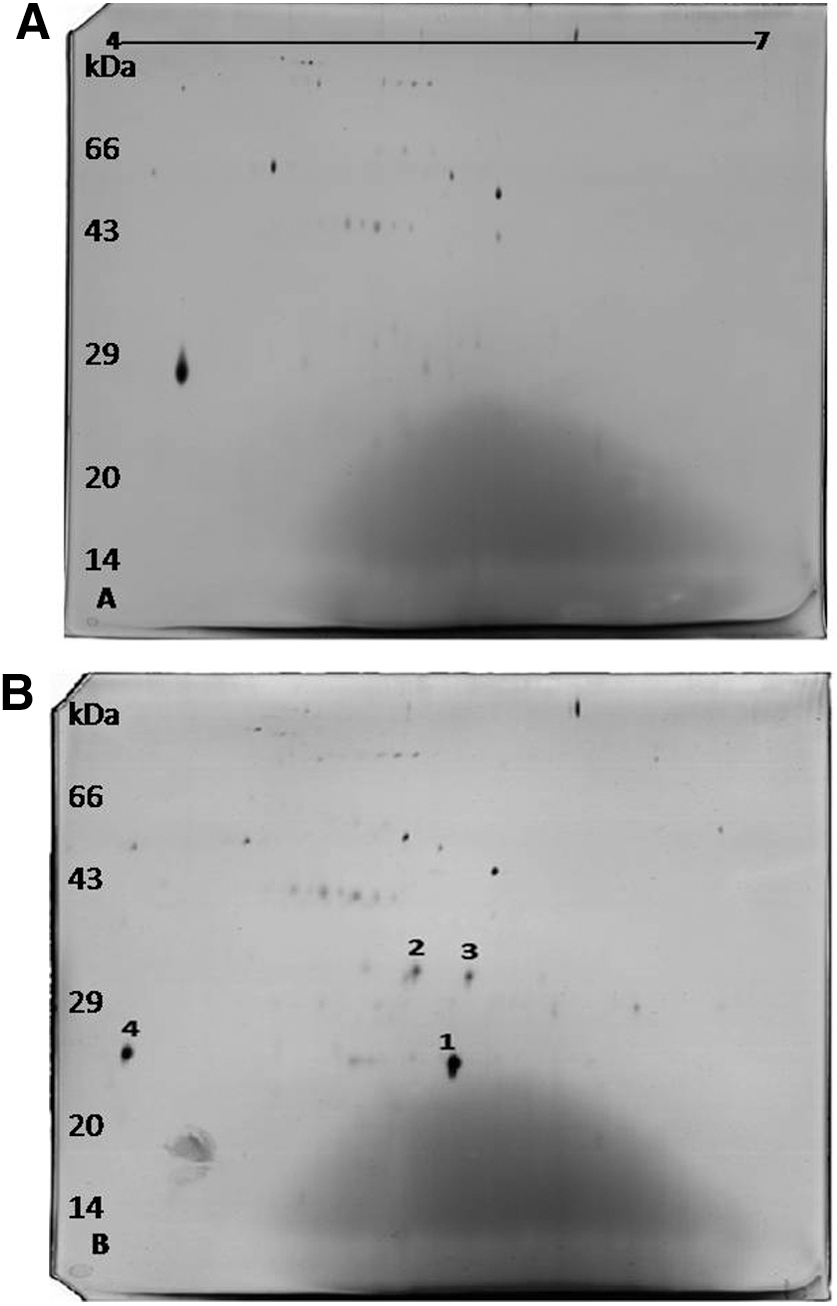
Two-dimensional electrophoretic gel images of culture supernatants of enteropathogenic SBANU8 strain incubated with and without pea sprouts.
Possible roles of trypsin inhibitors in plant–bacterial interaction
Bowman-Birk inhibitors are major groups of serine protease inhibitors in plants, especially the leguminous species. These proteins have two reactive sites and are able to independently as well as simultaneously inhibit trypsin and related enzymes such as chymotrypsin (Drame et al., 2013). These proteins, in addition to playing a role in seed germination and development (Deshimaru et al., 2003), are also part of defense mechanisms against pathogens (Qu et al., 2003). Supported from previous reports, the trypsin inhibitors observed in the present study can also be classified as defense proteins of plants against bacteria. Che et al. (2002) reported the expression of the Bowman-Birk coding gene during incompatible interactions of rice and Pseudomonas avenae. Transcriptional profiling studies have also revealed high expression of Bowman-Birk in rice–bacterial leaf blight pathogen interaction (Kotapalli et al., 2007). Miche et al. (2006) reported the protein as jasmonic acid inducible, along with other defense-related proteins including PR proteins.
The molecular mass of the Bowman-Birk inhibitor-like protein we observed in 2D gel was 23 kDa. The second unique protein of 26 kDa observed in 2D gel (trypsin inhibitor–based on mass spectrometry results) could be the same that was observed in SDS-PAGE analysis (Fig. 2). So far, Bowman-Birk inhibitor proteins are known as low molecular weight of 8-kDa proteins in legumes and 16-kDa proteins in monocots. Pea protease inhibitors characteristically have a molecular mass of 6–8 kDa (Ferrasson et al., 1997). The molecular mass of>20 observed in the present study may represent dimers or trimers of these inhibitors. Bowman-Birk as well as other trypsin inhibitors have been observed to self-associate in aqueous solutions to give dimers and trimers (Morrison et al., 2007). Trypsin inhibitors with greater molecular masses have been previously reported in peas (Tome’ et al., 1981). Similar to our observation in the present study, unusual Bowman-Birk protein with greater molecular mass of 29 kDa was observed previously (Miche et al., 2006) in rice. These observations indicate that the protein may form dimers and trimers, or the isoforms may be much more diverse in plants than was originally thought.
Serine proteases are known to play a role in colonization of the host plant by pathogenic bacteria (Stork et al., 2008). Production of extracellular serine protease by human pathogenic E. coli has also been reported (Brunder et al., 1997). The secretion of serine protease inhibitors by pea sprouts, only when challenged with a pathogenic strain, provides molecular clues for the possible production and involvement of E. coli serine protease in colonizing pea sprouts. Further large-scale proteome profiling and analysis with different varieties of pea and various time points of colonization is needed for confirmation. Nonetheless, this assumption adds to the growing evidence of similarities between plant pathogenic and human pathogenic bacteria in their plant-colonizing strategies.
Conclusions
Bowman-Birk inhibitors and their variants from legumes, such as soybean, pea, lentil and chickpea, are known for their potential health-promoting properties within the gastrointestinal tract (Clemente et al., 2011). They resist acidic condition and action of proteolytic enzymes to exert anticarcinogenic and anti-inflammatory properties. EPEC causes inflammation in the gastrointestinal tract as part of the processes in diarrheal diseases. The observed production of Bowman-Birk type protein by pea sprouts in response to pathogenic E. coli warrants correlated understanding with their well-known anti-inflammatory activity. This creates a strong base of evidence to further our understanding for food-processing technology applications.
Footnotes
Acknowledgment
The authors acknowledge the support of VIT University, Vellore and Tamil Nadu Agricultural University, Coimbatore in performing this research.
Disclosure Statement
No competing financial interests exist.