
Abstract
In France, Salmonella enterica serotypes Typhimurium and Derby are the most often isolated serotypes in pigs. Moreover, serotype Derby usually ranks between third and fourth in prevalence among human isolates in France. The aim of this study was to evaluate the genetic relationships between human and pig Salmonella Derby isolates based on their pulsed-field gel electrophoresis (PFGE) patterns after XbaI, BlnI, and SpeI restriction and on their antimicrobial resistance profiles. The 196 studied isolates were isolated in 2006 and 2007: 73 from fattening pigs, 27 from pork, and 96 from humans. Forty-four PFGE XbaI patterns were identified. A major pattern (SDX01) was identified for 96 isolates (49%). This pattern was common to pig, pork, and human isolates. Among the 146 isolates tested for their antimicrobial resistance, 84.2% (n=123) showed resistance to at least one antibiotic and 69.2% (n=101) were simultaneously resistant to at least streptomycin, sulfonamides, and tetracycline. Most of the isolates that are resistant to these three antibiotics also displayed the major SDX01 pattern. The use of two other restriction enzymes on a part of the panel (155 isolates) brought a significant increase in the discriminatory index, in particular for SDX01 strains. As Salmonella Derby is essentially isolated from pigs, and major resistance and PFGE patterns of isolates from pigs and pork were very similar to human isolates, human salmonellosis due to Salmonella Derby may be related to pigs.
Introduction
This serotype may also be implicated in larger outbreaks (Ebuchi et al., 2006) and has been among the 20 most frequently serotyped human Salmonella isolates in several European and Asian countries over the past decade (Hendriksen et al., 2011). Therefore, this serotype is a major public health concern.
This study aimed at evaluating the link between pigs, pork, and human isolates, in order to assess the farm-to-fork continuum. To demonstrate the spread of isolates from pigs via pork to humans, highly discriminatory typing methods are necessary. Traditional isolate typing methods such as serotyping or phage typing have been used for surveillance of Salmonella and outbreak investigations, but they are supplemented nowadays with the development of molecular methods (Wonderling et al., 2003; Gebreyes et al., 2006). Pulsed field gel electrophoresis (PFGE), which is currently the criterion standard, has generally good discriminatory power and has been proven to be highly useful and reliable, especially for tracking contamination sources (Botteldoorn et al., 2004; Vieira-Pinto et al., 2006) and for outbreak investigations (Noel et al., 2006; Dominguez et al., 2009). As the PFGE method with XbaI has shown low discriminatory power for Salmonella Derby (Kerouanton et al., 2007), we combined it with cluster analysis of multiple restriction enzymes (XbaI, BlnI, and SpeI) as recommended by Zheng et al. (Zheng et al., 2011). Because resistance of Salmonella to antimicrobial agents is a worldwide problem, and antimicrobial resistance has already been described in human (Ling et al., 2001) and nonhuman (Akiba et al., 2006) Salmonella Derby isolates, antibiotic drug susceptibility was also determined. The study was conducted over a 2-year period on 196 isolates isolated in 2006 and 2007, 73 of which were from fattening pigs, 27 from pork, and 96 from humans.
Materials and Methods
Salmonella Derby isolates
A total of 196 S. Derby isolates from various origins were considered in this study. The animal isolates (n=73) were collected from pig lymph nodes during the European baseline studies carried out in 2006–2007 (EFSA, 2008) by the National Reference Laboratory (NRL) for Salmonella (HQPAP unit ANSES Ploufragan). Isolates from pork (n=27), collected by the French Salmonella network (Associated NRL for Salmonella, ANSES Maisons-Alfort) in 2006 and 2007 were also studied. Moreover, the French National Reference Center for Salmonella (FNRC-Salm, Institut Pasteur, Paris) provided 96 Salmonella Derby isolates collected from human salmonellosis over the same period of time (47 isolates in 2006 and 49 in 2007). The set of isolates represented 34.3%, and 41.2% of all the Salmonella Derby isolates for 2006 and 2007, respectively. In terms of representativity, it has been estimated that the FNRC-Salm network detected 66% of confirmed human Salmonella infections in France at the time of the study (Jourdan-Da Silva and Le Hello, 2012). In each laboratory, isolates were serotyped, according to the White–Kauffmann–Le Minor scheme, on the basis of somatic O and phase 1 and phase 2 flagellar antigens by agglutination tests with antisera (Bio-Rad, Marnes la Coquette, France; Eurobio, Les Ulis, France; World Health Organization Collaborative Center for Reference and Research on Salmonella, Institut Pasteur, Paris, France).
PFGE typing
The genetic typing of the isolates was carried out by restriction fragment length polymorphism–PFGE with a CHEF-DR III system (Bio-Rad) according to the PulseNet protocol (Ribot et al., 2006). Salmonella Braenderup H9812 strain was used as a molecular size marker (Hunter et al., 2005). After staining with ethidium bromide, DNA patterns were visualized under ultraviolet light and images were taken with the Biovision+ 1500/36M X PRESS system (Fisher Bioblock). BlnI and SpeI enzymes were also used on a panel of 155 isolates. This panel is representative of isolates that did not give a unique pattern after XbaI restriction. For BlnI-digested DNA, migration conditions were the same as for XbaI: fragments were resolved on 1% agarose gel using electrophoresis conditions of 6.0 V/cm at 14°C for 20 h. Pulse times were ramped from 2 s (initial switch) to 64 s (final switch). For SpeI-digested DNA, migration was performed in two steps, a 11.5-h step with an initial switch time of 20 s and a final switch time of 40 s, followed by a 9.5-h step with an initial switch time of 7 s and a final switch time of 13 s, at a gradient of 6 V/cm and an included angle of 120°. For cases of DNA lysis, we used an adapted DNA extraction protocol with HEPES buffer instead of Tris buffer, and Tris borate EDTA (TBE) migration was performed with 100 μM thiourea (Liesegang and Tschape, 2002; Silbert et al., 2003).
Genetic profile analysis
Fragment size estimations and analysis of the similarities between genotypes were carried out using the BioNumerics® software (version 6.5, Applied Maths, Kortrijk, Belgium). The similarities were calculated using Dice's coefficient with a maximum tolerance of 1%, and dendrograms were built according to the unweighted-pair group method (Struelens, 1996).
The Simpson index (Hunter and Gaston, 1988) was calculated to estimate the diversity of the sample, and the 95% confidence intervals were also calculated based on the variance as suggested by Grundmann et al. (2001).
Antimicrobial susceptibility tests
All pig and pork isolates were tested for their antimicrobial susceptibility. Resistance phenotypes were determined using the disc diffusion method as recommended by the Antibiogram Committee of the French Society for Microbiology (
Results
PFGE types after XbaI restriction
The 196 isolates generated 44 different PFGE XbaI patterns (Fig. 1). The discriminatory ability (D-value) of the method for the entire panel was 0.75 (95% confidence interval 0.68–0.81).
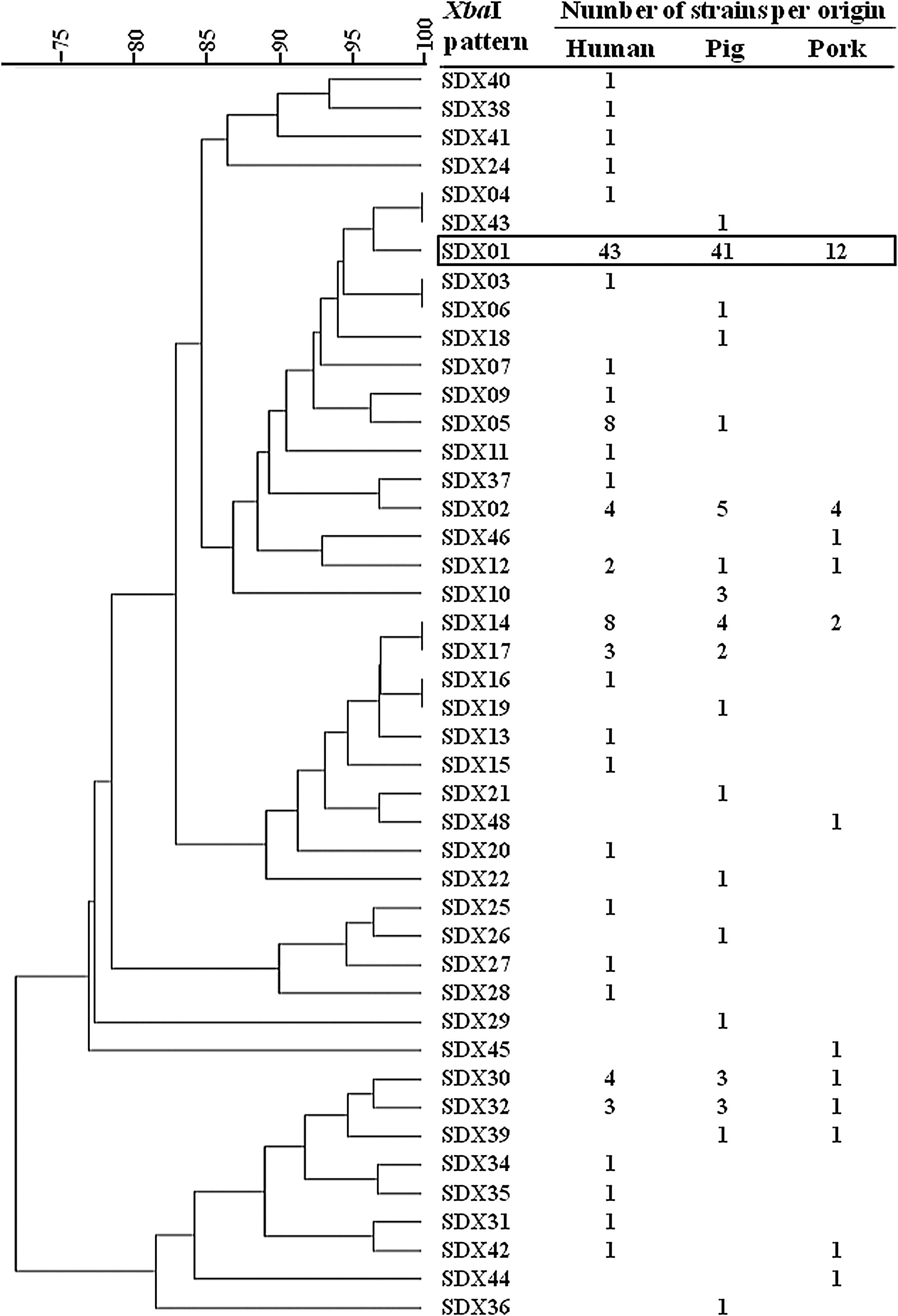
Dendrogram representing the 44 pulsed-field gel electrophoresis (PFGE) XbaI patterns designated SDX01 to SDX48. The number of isolates of each source (human, pig, and pork) is shown on the right side. The major PFGE profile SDX01 is circled. NB: Some patterns appear to be 100% identical with BioNumerics® analysis (1% tolerance), but they have been visually identified as different from each other.
Thirty-three PFGE patterns (75%) were encountered for a single isolate and then specifically associated to a single origin: four PFGE patterns were attributed to a single pork isolate, 9 to pig isolates, and 20 to human isolates.
One other pattern was also related to a single origin but grouped together three pig isolates.
Conversely, 10 patterns grouped together isolates of different origins. Six PFGE patterns were common to pig, pork, and human isolates (SDX01, 02, 12, 14, 30, and 32), two were common to human and pig isolates (SDX05 and 17), one to human and pork isolates (SDX42), and one to pig and pork (SDX39). Among these 10 patterns, SDX01 pattern was identified for 96 isolates among the 196 studied with 44.8% of those of human origin (43/96), 56.2% of those of pig origin (41/73), and 44.4% of those of pork origin (12/27). Actually, there was no significant difference (chi square, p=0.30) allowing this major pattern to be assigned to one of the origins.
Fourteen PFGE patterns—SDX02, SDX03, SDX04, SDX05, SDX06, SDX07, SDX09, SDX10, SDX11, SDX12, SDX18, SDX37, SDX43 and SDX46 patterns—were very similar to the major SDX01 pattern (86% similarity), and comprised 39 isolates. Thus, a total of 135 isolates among the 196 studied (68.9%) were grouped into a single major cluster. This cluster included 65.6% of the human isolates, 74% of the pig isolates and 66.7% of the pork isolates.
PFGE types after BlnI and SpeI restriction
Isolates (n=155) of various origins grouped into eigh of the 10 PFGE XbaI patterns gathering several isolates (isolates that did not give a unique pattern after XbaI restriction) were also analyzed after BlnI and SpeI restriction. A total of 18 BlnI PFGE profiles and 27 SpeI PFGE profiles have been highlighted. Eight isolates were nontypable with BlnI enzyme despite the addition of thiourea. A combination of the results obtained with the three enzymes gave 42 combined PFGE patterns (Fig. 2). The discriminatory index for this panel of 155 isolates was of 0.53, 0.60 and 0.68 when using respectively BlnI, XbaI, and SpeI, and increase to 0.72 when considering the results obtained with the three enzymes.
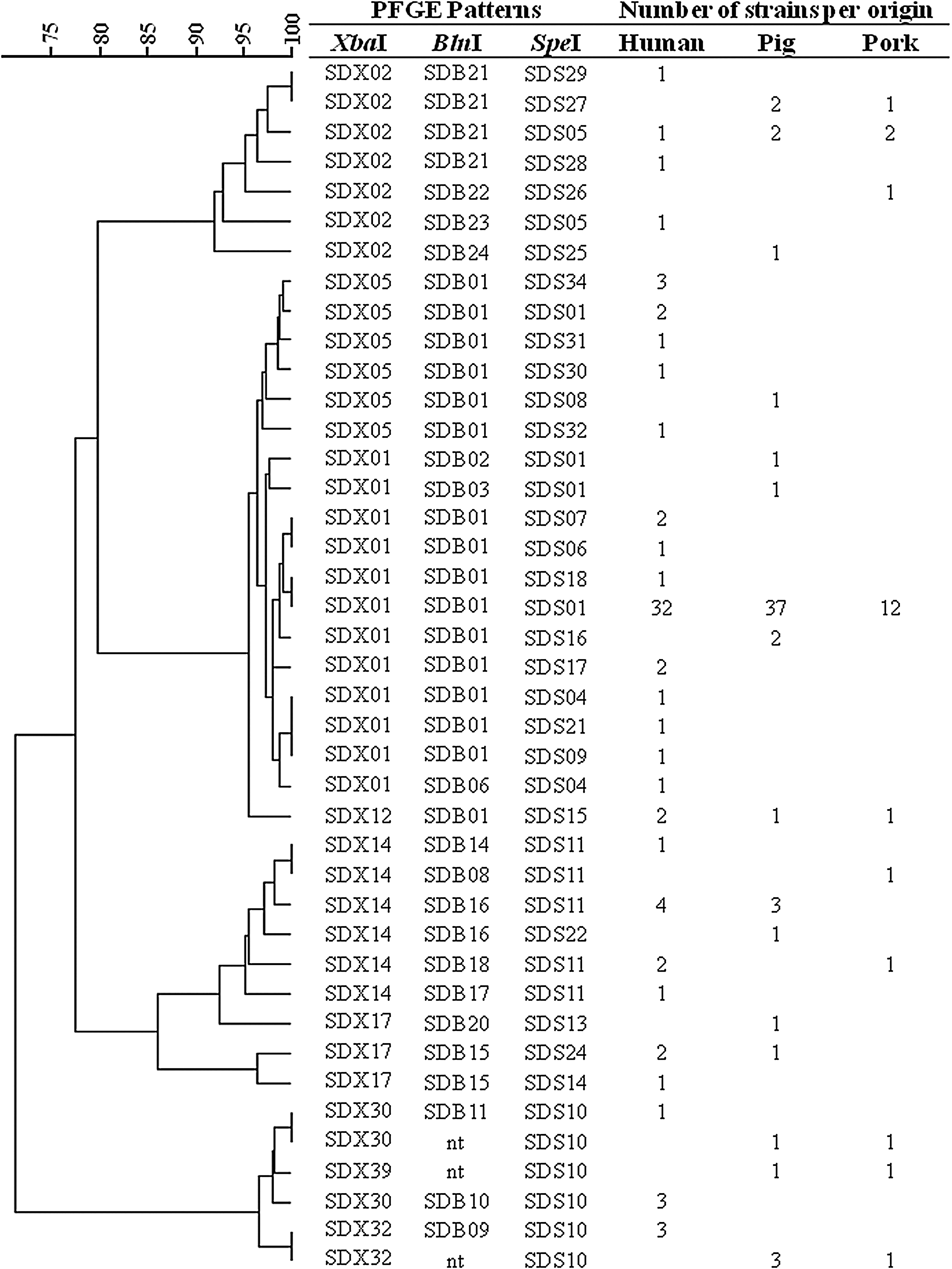
Dendrogram representing the 42 XbaI-BlnI-SpeI combined patterns obtained on 170 strains (nt, nontypeable strains). The number of strains of each origin is shown on the right side. PFGE, pulsed-field gel electrophoresis.
Most of the time, the use of the two others enzymes increased the discriminatory power of the PFGE method. For SDX05, SDX30, and SDX32 patterns, the use of BlnI and SpeI separated the human isolates from the others sources.
Sometimes, the use of BlnI and SpeI gave exactly the same discrimination than that obtained with XbaI. For example, for the four isolates carrying SDX12, the combined pattern SDX12/SDB01/SDS15 always correspond to two human, one pig, and one pork isolates, as observed after only XbaI restriction.
Similar BlnI and/or SpeI patterns were also observed for different XbaI patterns. For example, the SDB01 pattern was highlighted for isolates carrying SDX01, SDX05, and SDX12 patterns; and SDS10 for isolates carrying SDX30, SDX32, and SDX39 patterns. Moreover, we also noted that all animal isolates from these 3 XbaI patterns were nontypable by BlnI enzyme.
Antimicrobial susceptibility
No more than 15.5% (23/147) of the tested isolates were susceptible to all antimicrobials. For the remaining isolates, 11 different patterns of resistance were observed, with from one to six resistances (Table 1). Seventy-nine percent of the isolates showed at least tetracycline (TE) resistance. TE resistance was often associated with streptomycin (S) and sulfonamides (SSS) resistance. Hence, 98 isolates (66.7%) displayed the S SSS TE pattern of resistance. Four isolates were resistant to more than three antimicrobials. However, no resistance to fluoroquinolone or third-generation cephalosporins was detected. For each resistance pattern, Table 1 shows the corresponding PFGE patterns. The 102 isolates harboring at least resistance to TE, S, and SSS were subdivided into 18 PFGE patterns while 75 isolates (73.5%) belonged to the major PFGE pattern SDX01.
No., number of isolates; TE, tetracycline; S, streptomycin; SSS, sulfonamides; TMP, trimethoprim; A, ampicillin; K, kanamycin; TOB, tobramycin; GM, gentamicin.
Discussion
The genetic diversity of Salmonella Derby isolates has rarely been investigated. In this study, a large panel of 196 isolates was studied, and molecular characterization by PFGE showed that the genome of this serotype seems to have a highly homogeneous genetic composition. The discriminatory ability (D value) of the method was 0.75 (0.68–0.81) for the entire panel, confirming a previous study (Kerouanton et al., 2007). A major XbaI pattern, SDX01, was found for 49% (96/196) of the isolates. This PFGE pattern was also observed in a German study by Hauser et al. (2011) but only for one isolate, while their main PFGE pattern, corresponding to our SDX42 pattern, was only found for two of our isolates (one pork and one human). This suggests that each country could have its own prevalent Salmonella Derby pattern.
In our study, nine patterns presented more than 90% similarity with the major PFGE pattern and 14 patterns more than 86% similarity.
Among the 96 tested isolates carrying the pattern SDX01, four BlnI and nine SpeI PFGE profiles were identified. However, among these 96 isolates, 81 isolates (32, 37, and 12 of human, pig, and pork origin, respectively) remain identical after the use of BlnI and SpeI enzymes. Isolates with identical genetic profiles came from all tested sources. This result could suggest either that these isolates are closely related and can be carried by different hosts or that PFGE is not an appropriate method for Salmonella Derby. Several other typing methods based on polymerase chain reaction amplification and capillary electrophoresis or sequencing such as multilocus variable-number of tandem-repeats analysis (MLVA) (Lindstedt et al., 2003; Boxrud et al., 2007; Bergamini et al., 2011) and MLST (Torpdahl et al., 2005; Ben-Darif et al., 2010; Achtman et al., 2012) and CRISPR (Fabre et al., 2012) are now used for Salmonella discrimination, but they have rarely been described for Salmonella Derby isolates. Genetic diversity of Salmonella Derby evidenced after use of numerous methods was recently discussed (Hauser et al., 2011), but the conclusion was that PFGE remained the method with the highest index of diversity followed by VNTR sequence typing (STTR5 primer only) and sop genes (sopA, sopB, and sopD) sequence typing (sop-ST). MLST or DNA microarray on 275 genes provided lower discrimination of the 82 isolates studied. Consequently, according to the observations of Hauser et al. (2011) with regard to PFGE, our results seem to indicate that our isolates, with similar profiles after three different digestions, are genetically similar and have the ability to colonize different hosts.
The antimicrobial susceptibility tests revealed a low percentage (15.5%) of wild-type Salmonella Derby isolates (i.e., with no acquired mechanisms of resistance). This rate of wild-type Salmonella Derby isolates differs greatly, depending on studies: from 0% (Ellerboek et al., 2010) to 82% (Piras et al., 2011). In our study, the majority of isolates (70.7%) were resistant to three antimicrobials, including the S SSS TE pattern that accounted for 66.7% of the isolates. Resistance to these three antimicrobials was already highlighted for pig isolates (Michael et al., 2006b) and pork meat (Mürmann et al., 2009; Anjum et al., 2011). Antimicrobial resistance has been commonly associated with extended use in farm animals (Wegener et al., 2003). While it is true that, in France, tetracycline is the most commonly used antimicrobial for pigs (Chevance and Moulin, 2011), consumption of antibiotics in pigs and resistance in Salmonella Derby cannot be systematically linked. Although penicillins are used as often as sulfonamides, no resistance to β-lactam antibiotics has been detected in any isolate from pigs or pork, whereas resistance to sulfonamides was detected in most isolates.
A correlation between macrorestriction patterns and resistance patterns was previously reported (Botteldoorn et al., 2004; Michael et al., 2006a; Michael et al., 2006b). This correlation was also clearly observed in our study, as the major PFGE pattern SDX01 was linked to the major antimicrobial resistance pattern: 75 (76.5%) of the 98 isolates resistant to streptomycin, sulfonamides, and tetracycline were from the SDX01 pattern. All SDX01 isolates tested (n=75) were resistant to these three antibiotics. A recent study (Cardoso et al., 2011) showed that each Salmonella Derby isolate presenting a major PFGE pattern also exhibited the S SSS TE pattern and carried a class 1 integron. Indeed, class 1 integrons carry the sul1 gene encoding sulfonamide resistance and frequently contain the aadA cassette gene, which is associated with streptomycin/spectinomycin resistance (Michael et al., 2006a). Presence of a class 1 integron and aadA gene in Salmonella Derby from pigs was also reported by Gebreyes et al. (2004). Less frequently, Salmonella Derby isolates from Asia have been found to contain the Salmonella genomic island 1 (SGI1) variant, which carries aadA2, sul1, floR, and tet(A) resistance genes (Chiu et al., 2007).
As reported by Huang et al. (2009), in the pig, Salmonella Derby isolates showed a low resistance rate to β-lactams. Ampicillin resistance was found only in one human isolate. In our study, only four isolates showed resistance to more than three antibiotics; two were isolated from humans, one from pigs, and one from pork. Pigs and pork isolates only showed resistance to streptomycin, sulfonamides, trimethoprim, and tetracycline. Nevertheless, these antimicrobials are not the first used as a treatment for human salmonellosis.
A statistical attribution study based on outbreak data (Pires et al., 2010) estimated that less than 1% of outbreak-associated salmonellosis cases were attributed to pork in Europe in 2005 and 2006. However, in a report based on community observations, the European Food Safety Authority (EFSA) estimated that 10–20% of human Salmonella infections in the European Union (EU) may be attributable to the pig reservoir (EFSA, 2010), and a more recent study (Pires et al., unpublished data) found the pig reservoir to be the second largest contributor to human salmonellosis in the EU, responsible for 29.6% (95% confidence interval: 28.9–30.3%) of reported cases. Pires et al. (unpublished data) also showed that pigs were the main source of human salmonellosis in eight countries.
In this epidemiological study, isolates of human and pig origin showed a very high similarity in PFGE and in antimicrobial susceptibility. As serotype Derby is mostly associated with pigs and is the most prevalent, along with serotype Typhimurium, in pig production in France, we can conclude, as did Finley et al. (2008), that human salmonellosis due to this serotype is in all likelihood linked to pigs. Implementation of another molecular method that is more discriminatory than PFGE, as a complete MLVA scheme, and additional studies on isolates from diverse origins could help us to confirm or disprove this hypothesis.
Footnotes
Disclosure Statement
No competing financial interests exist.