
Abstract
Vaccination is the most effective and cost-effective way to treat hepatitis B virus (HBV) infection. Collective data suggest that helminth infections affect immune responses to some vaccines. Therefore, it is important to reveal the effects of helminth infections on the efficacy of protective vaccines in countries with highly prevalent helminth infections. In the present work, effects of Trichinella spiralis infection on the protective efficacy of HBV vaccine in a mouse model were investigated. This study demonstrated that the enteric stage of T. spiralis infection could inhibit the proliferative response of spleen lymphocytes to hepatitis B surface antigen (HBsAg) and lead to lower levels of anti-HBsAg antibodies, interferon-γ, and interleukin (IL)-2, along with higher levels of IL-4 and IL-5. However, these immunological differences are absent in the muscle stage of T. spiralis infection. The results suggest that the muscle stage of T. spiralis infection does not affect the immune response to HBV vaccination, while the enteric-stage infection results in a reduced immune response to HBsAg.
Introduction
H
About 60% of the world's population, especially in Southeast Asia and Africa, live in highly endemic HBV areas (Shepard et al., 2006). Helminth infections are reported to affect more than one billion people in developing countries (Brooker, 2010). Southeast Asia is classified as a highly HBV-endemic area, where helminth infections are also highly prevalent (Conlan et al., 2011; Cui et al., 2011; Luo et al., 2012). Epidemiological surveys in China find that the rate of nonresponse or weak response to HBV vaccine is much higher in rural children than that in urban children (Zai, 2002). The rate of nonresponses or weak responses to HBV vaccine was 83.3% in patients with schistosomiasis japonica, while it was only 7.7% in healthy persons (Song et al., 2005). Helminth infections might be an important cause for nonresponse or weak response to HBV vaccine (Michel and Tiollais, 2010; Chen et al., 2012).
Trichinellosis, caused by infection with the genus Trichinella, is one of the most widely distributed foodborne parasitic zoonoses worldwide (Conlan et al., 2011; Cui et al., 2011; Bruschi, 2012; Papatsiros et al., 2012). It is estimated that around 10 million people might be infected with long-lasting persistence of muscle larvae of Trichinella (Dupouy-Camet, 2000; Gottstein et al., 2009). For the establishment of T. spiralis infection, modulation of the host's immune response is necessary to avoid their own destruction (Else, 2005; Bai et al., 2012; Bruschi and Chiumiento, 2012). It has been shown that T. spiralis possesses a capacity to retune the immune cell repertoire and modulate the host response not only to itself but also to third-party antigens (Furze et al., 2006; Bruschi and Chiumiento, 2012).
Considering the extent of geographical overlap of trichinellosis and HBV infections (Shepard et al., 2006; Conlan et al., 2011), it is of great importance to uncover the effects of T. spiralis infection on protective efficacy of HBV vaccine. In the present work, effects of different stages of T. spiralis infection on the protective efficacy of HBV vaccine were investigated.
Methods
Mice and parasites
T. spiralis was isolated from domestic pigs in Hubei Province, China and confirmed by polymerase chain reaction as described previously (Zhang et al., 1997). The life cycle was maintained by serial passage in Kunming mice at 6–8-month intervals. Male BALB/c mice, aged 6–8 weeks, were purchased from Wuhan Institute of Biologic Products (Wuhan, China). The mice were maintained under a standard specific pathogen-free condition at Tongji Medical College, Huazhong University of Science and Technology (Wuhan, China). The experiment was approved by the Committee on Animal Research of Tongji Medical College. All experiments were in conformance to local government regulations that, in turn, complied with Chinese national laws on animal ethics.
Experimental infection and immunization procedure
Male BALB/c mice were divided randomly into five groups (n=8): the immunization control (I), the enteric-stage infection plus immunization (E+I), the enteric-stage infection (E), the muscle-stage infection plus immunization (M+I), and the muscle-stage infection (M) groups. Infective muscle larvae of T. spiralis were recovered from the infected mice by acid pepsin digestion as described previously (Li et al., 2010). All groups excluding group I were infected by oral gavage with 500 larvae per mouse. The infection was confirmed by checking the carcass of each infected mouse under a microscope at the end of the study. Groups E+I and M+I were vaccinated at 7 days and 60 d after T. spiralis infection, respectively. Groups I, E+I, and M+I were vaccinated with the recombinant yeast-derived HBV vaccine (Kangtai Biologic Products Ltd., Wuhan, China) via subcutaneous inoculation in the back, 0.1 μg per gram body weight, for three times at an interval of 14 d (Chen et al., 2012).
Detection of anti-HBsAg antibodies in serum
Fourteen days after the final inoculation, about 500 μL of blood per mouse was collected from orbital veins and allowed to clot for 4 h at room temperature. Serum was subsequently separated by centrifugation at 3000×g for 15 min. All sera samples were then stored at −20°C for further antibody detection. Anti-HBsAg antibodies in sera were determined by the enzyme-linked immunosorbent assay (ELISA) Kit (Kehua Bio-engineering Ltd., Shanghai, China) according to the manufacturer's instruction. The reactions were measured at 450 nm by a MK3 microplate reader (Thermo Labsystems, Vantaa, Finland).
Splenocytes proliferation
All mice were anesthetized with isoflurane and sacrificed at 14 d after the final vaccination, and splenocyte suspensions were harvested. Splenocytes (2×105 cells per well) were cultured for 48 h in 96-well plates, in triplicate, in RPMI-1640 medium containing 5% fetal calf serum (FCS) at 37°C in a 5% CO2 incubator. Then cultures were incubated with HBsAg (2 μg/mL) for another 48 h, finally for an additional 4 h with 10 μL methylthiazolyl tetrazolium (5 mg/mL, Sigma). Unstimulated cultures were used as controls. Supernatants were then removed and discarded. Following addition of 100 μL of dimethyl sulfoxide (Sigma) per well, absorbances were read at 570 nm by an MK3 microplate reader. Splenocyte proliferation responses were expressed as stimulation index (SI), calculated as the ratio of the mean absorbance of triplicate wells stimulated with HBsAg to that of unstimulated ones.
ELISAs for cytokines in splenic cell culture supernatant
Splenocyte suspensions were prepared from all mice sacrificed at 14 d after the final vaccination (Zhao et al., 2013). Single cells (5×106 cells/well) were cultured in RPMI-1640 with 10% FCS and 1% penicillin and streptomycin. The cultures were then incubated at 37°C in 5% CO2 stimulated with HBsAg (2 μg/mL) for 72 h. The concentrations of interleukin (IL)-4, IL-5, interferon (IFN)-γ, IL-2, and IL-10 in spleen cell culture supernatants were determined according to the instruction of the ELISA kits (eBioscience, USA). In brief, IL-4, IL-5, IFN-γ, IL-2, and IL-10 of all the mice were detected by biotinylated monoclonal antibodies, which were evidenced by avidin-conjugated horseradish peroxidase and followed by incubation with substrate tetramethylbenzidine dihydrochloride (TMB, Tiangen, China). The reactions were measured at 450 nm using an MK3 microplate reader. Cytokine concentrations were determined by reference to standard curves constructed with known amounts of mouse recombinant IL-4, IL-5, IFN-γ, IL-2, and IL-10.
Statistical analysis
All data were expressed as means±standard deviation and analyzed using one-way analysis of variance. P value of<0.05 was regarded as statistically significant and p value<0.01 as highly significant. All procedures were repeated twice to evaluate the reproducibility of those results.
Results
Anti-HBsAg antibody levels in serum
The serum levels of specific anti-HBsAg antibodies were determined at 14 d after the final vaccination. As shown in Figure 1, there was no significant difference between groups M+I and I. However, the E+I group had a much lower anti-HBsAg level than the immunization control (group I, p<0.01).
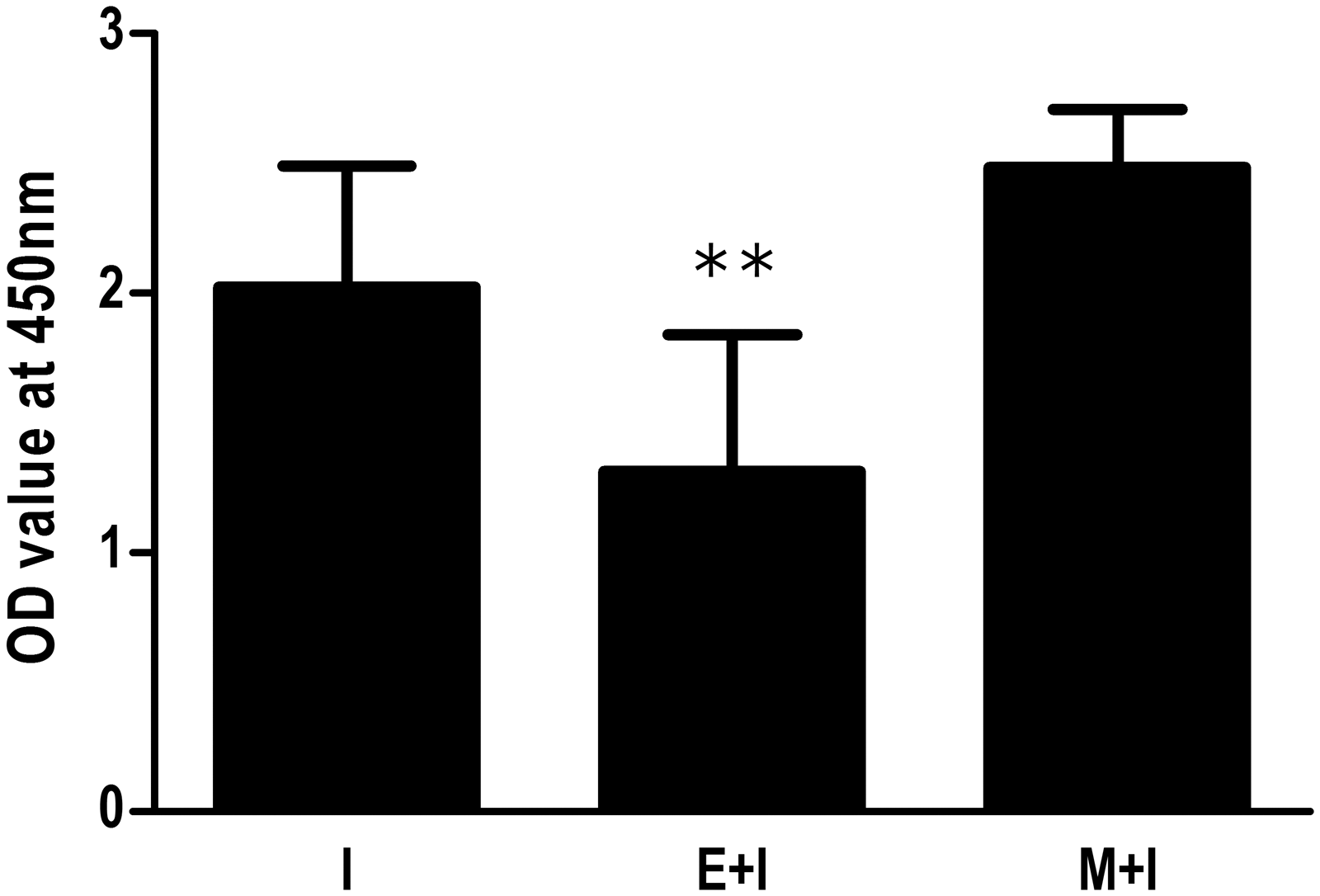
Effect of T. spiralis infection on serum levels of anti-HBsAg antibodies after HBV vaccination. The enteric stage infection plus immunization (E + I) and muscle stage infection plus immunization (M + I) groups were infected by oral gavage with 500 larvae of T. spiralis per mouse. Groups E + I and M + I were vaccinated at 7 days and 60 days after infection, respectively. All groups were vaccinated with the recombinant yeast derived HBV vaccine via subcutaneous inoculation in the back, 0.1 μg per gram body weight, for 3 times at an interval of 14 days. The serum levels of specific anti-HBsAg antibodies were determined at 14 days after the final vaccination. Antibody levels were expressed as the mean OD value ± SD of the individual serum from two independent combined experiments. OD, optical density. **p < 0.01, versus the immunization control group (I).
Splenocytes proliferation
The results of splenocytes proliferation assay are shown in Figure 2. The muscle stage of T. spiralis infection did not affect SI to specific HBsAg, while mice from the E+I group were found to produce a significantly lower SI (p<0.01).
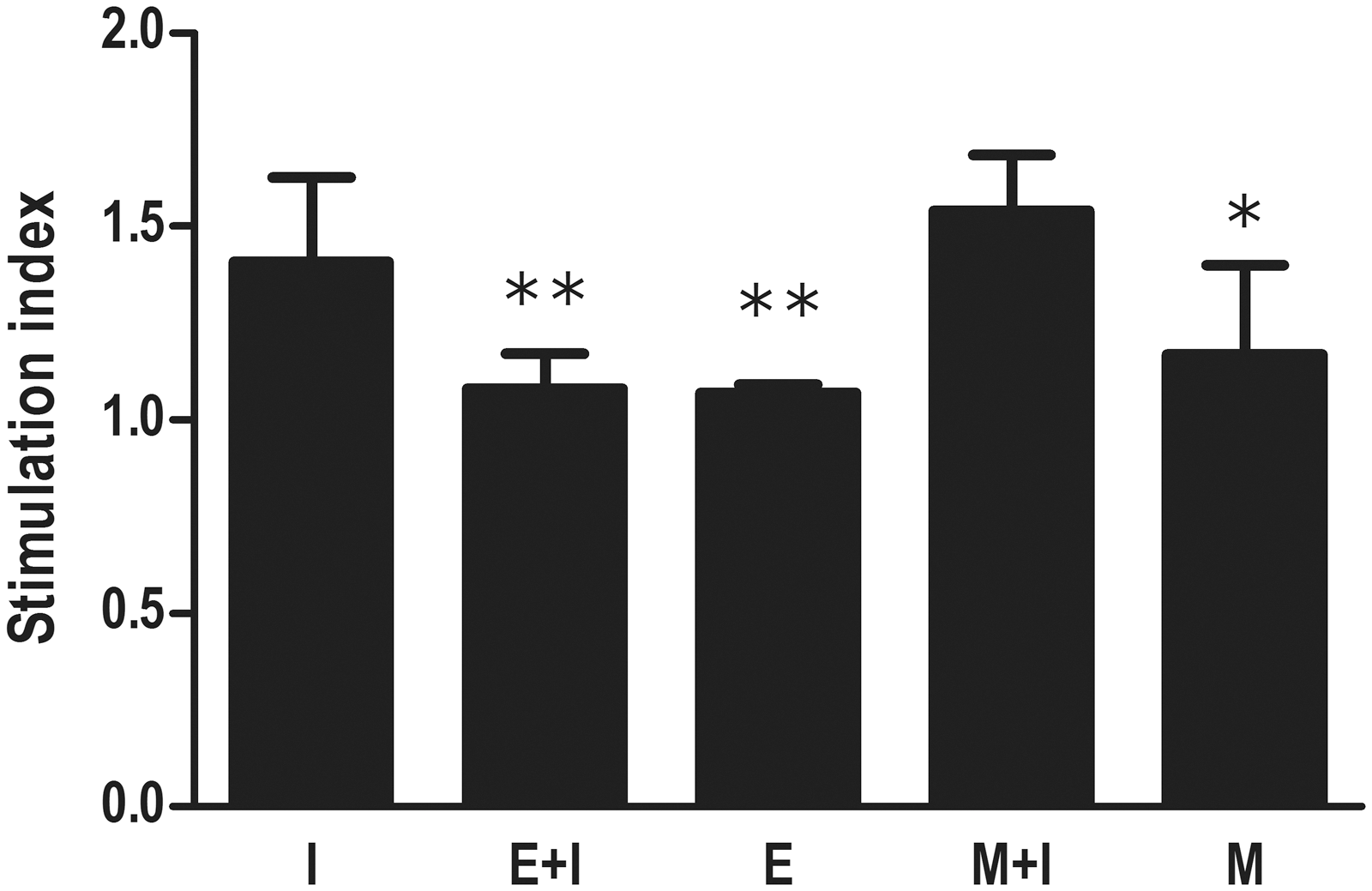
Effect of T. spiralis infection on splenocytes proliferation after HBV vaccination. All groups excluding the immunization control (I) were infected by oral gavage with 500 larvae per mouse. The enteric stage infection plus immunization (E + I) and muscle stage infection plus immunization (M + I) groups were vaccinated at 7 days and 60 days after T. spiralis infection, respectively. Groups I, E + I, and M + I were vaccinated with HBV vaccine via subcutaneous inoculation for 3 times at an interval of 14 days. Splenocytes proliferation was performed using an MTT assay at 14 days after the final vaccination. The level of splenocytes proliferation was expressed as simulation index (SI). Data were expressed as the mean SI ± SD from two independent combined experiments. E, the enteric stage infection group; M, the muscle stage infection group. *p < 0.05, **p < 0.01, versus group I.
Cytokine production by splenocytes
The cytokine production of spleen cells in response to HBsAg was measured at 14 d after the final vaccination. As Figure 3 shows, the E+I group had higher levels of IL-4 and IL-5, along with lower levels of IL-2 and IFN-γ when compared to group I (p<0.01). However, there were no differences in all the above cytokine levels between groups M+I and I.
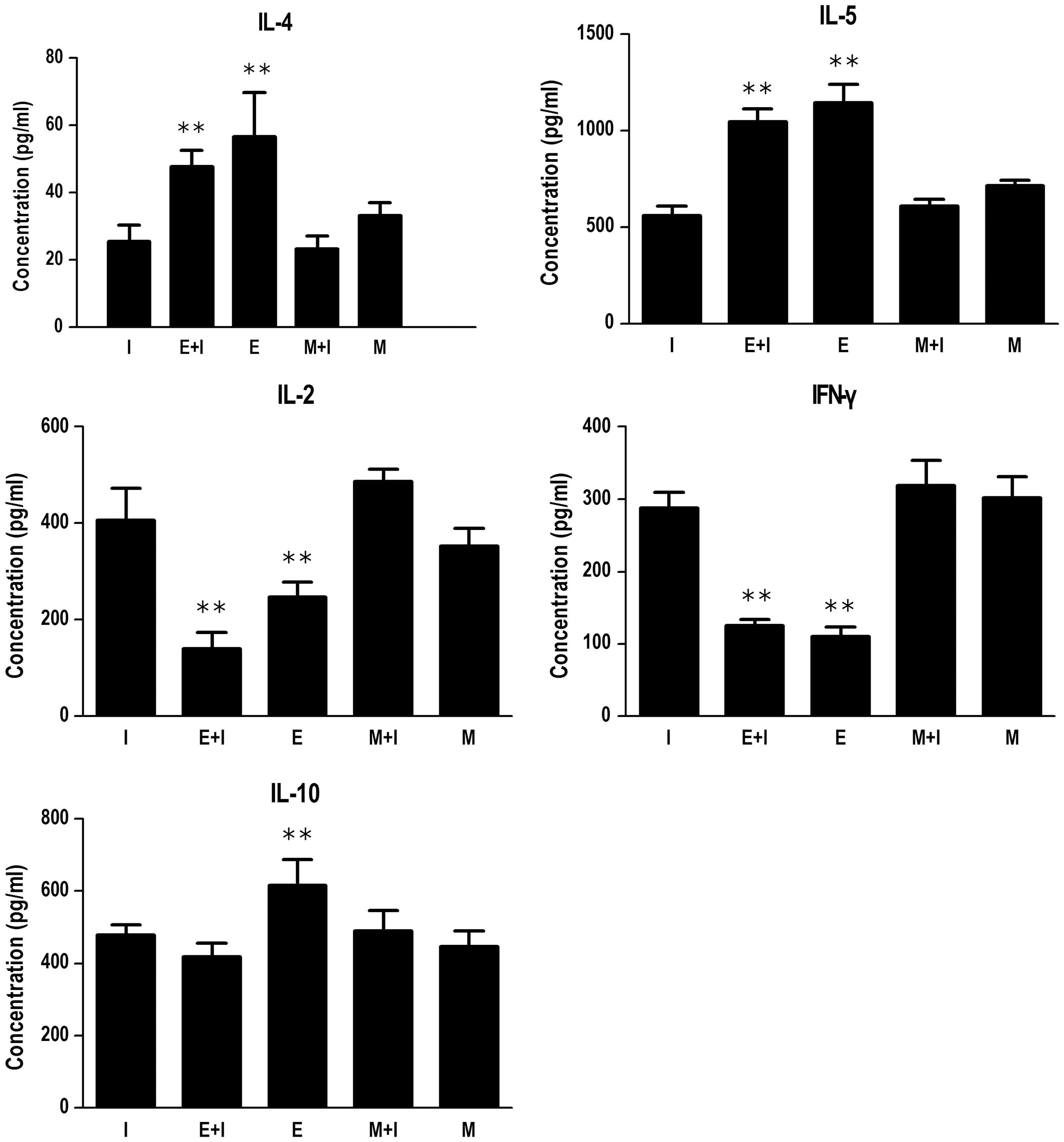
Cytokine expression levels in splenocytes of T. spiralis–infected mice after HBV vaccination. All groups excluding the immunization control (I) were infected by oral gavage with 500 larvae per mouse. The enteric stage infection plus immunization (E + I) and muscle stage infection plus immunization (M + I) groups were vaccinated at 7 days and 60 days after T. spiralis infection, respectively. Groups I, E + I, and M + I were vaccinated with HBV vaccine via subcutaneous inoculation for 3 times at an interval of 14 days. The levels of cytokines were measured in the supernatant using ELISA kits in accordance with the manufacturer's instructions. The cytokine production of spleen cells in response to HBsAg was measured at 14 days after the final vaccination. Data were expressed as the mean ± SD from two independent combined experiments. E, the enteric stage infection group; M, the muscle stage infection group; IL, interleukin; IFN, interferon. **p < 0.01, versus group I.
Discussion
Universal HBV vaccination is the key to reduce the infection and chronicity rates of HBV (Luo et al., 2012). However, 5–10% of individuals do not experience a response with an adequate antibody level to HBsAg (Sjogren, 2005). As evidenced by several studies, the ability of the host to mount an immune response is greatly determined by the pre-existing state of the immune system (Borkow and Bentwich, 2000, 2002; Elias et al., 2006). Chronic helminth infection is reported to be one of the contributing causes of absent or weak response to some vaccines, and consequently vaccines fail to confer protection against their respective infections in helminth-endemic areas (Borkow and Bentwich, 2000; Elias et al., 2006; Rafi et al., 2012). Our group also demonstrates that chronic Schistosoma japonicum infection reduces immune response to HBV vaccine (Chen et al., 2012).
It has been shown that T. spiralis possesses a capacity to retune the immune cell repertoire and modulate the host response. Furze et al. (2006) reports that the enteric stage of T. spiralis infection can suppress the pathological reaction caused by influenza. Trichinella infection in animal models ameliorates the outcome of immunological diseases or co-infection (Bruschi and Chiumiento, 2012). The present data showed that the enteric-stage infection had a decreased cell-mediated response to specific HBsAg in vivo. Also, such effects apparently were associated with lower levels of IL-2 and IFN-γ, along with higher levels of IL-4 and IL-5. The production of anti-HBsAg antibodies is dependent on the stimulation of Th1 helper lymphocytes driven by IL-2 and IFN-γ (Böcher et al., 2000; Michel and Tiollais, 2010). The failure to produce anti-HBsAg antibodies might be a consequence of the insufficient HBsAg-specific Th1-cell response (Aypak et al., 2012). T. spiralis larva intestinal invasion induces the production of Th2-related cytokines (Kang et al., 2012). The immune modulation from Th1 to Th2 induced by helminth infections can impair the specific immunity induced by tetanus toxoid (Sabin et al., 1996), bacille Calmette-Guérin, and human immunodeficiency virus vaccines (Borkow and Bentwich, 2002). Human ascariasis is reported to suppress the IL-2 response to recombinant cholera toxin B subunit and thus reduce the protective efficacy of the live oral cholera vaccine (Cooper et al., 2001). Thus, the inability to mount a protective level of protective antibodies after vaccination is presumably the consequence of a defect in the cellular immune regulation.
Perhaps somewhat surprisingly, both the enteric-stage and muscle-stage infections did not display a high level of IL-10 produced by splenocytes after HBV vaccination. IL-10 is a potent anti-inflammation cytokine and has been shown to prevent inflammation during the enteric stage of T. spiralis infection (Furze et al., 2006; Bruschi and Chiumiento, 2012). However, the chronic inflammation of T. spiralis infection is controlled independently of IL-10 at the muscle stage (Furze et al., 2006). In the current study, there were no changes in the two experiment groups. The low level of IL-10 might be associated with reduced numbers of IL-10-secreting lymphocytes (Furze et al., 2006; Bruschi and Chiumiento, 2012). Further research is needed to understand the fine mechanisms of such an observation.
It is generally believed that chronic helminth infections could reduce the body's immune response to HBV vaccine (Borkow and Bentwich, 2002; Chen et al., 2012). However, our result showed that the muscle stage of T. spiralis infection did not affect the immune response to HBV vaccination in vivo. This is different from the immunomodulation seen during the chronic infection with most other helminths, due to biological differences of parasites (Maizels and Yazdanbakhsh, 2003). Compared to the parasite in chronic Schistosoma infection being continuously exposed to the immune system, T. spiralis, which is usually encysted in the muscle stage, might be less actively dealt with by the immune response, and thus the muscle stage of T. spiralis infection leads to different results regarding vaccination attempts. Furthermore, the immune modulation caused by T. spiralis infection is closely related to the infection stage (Lubiniecki and Cypess, 1975; Furze et al., 2006; Wang et al., 2009). Lubiniecki and Cypess (1975) report that the muscle stage of T. spiralis infection has no antitumor effect, while the enteric-stage infection can inhibit tumor growth. Long-term residence of T. spiralis in skeletal muscle has no effect on influenza-induced pathology (Furze et al., 2006). The main reason is that Trichinella antigens are stage-specific and can induce specific host immune responses in different infection stages (Liu et al., 2007; Zocevic et al., 2011; Bruschi and Chiumiento, 2012). Muscle larvae and adult worms generally induce a Th2-bias response in the enteric stage, while newborn larvae are considered to induce Th1/Th2 mixed immune response during the muscle phase (Grencis et al., 1991; Kelly et al., 1991). Therefore, with respect to T. spiralis infection, the specific immune responses, caused by different-stage antigens, play a crucial role in determining the outcome of vaccination.
Taken together, to the best of our knowledge, this is the first report indicating that the muscle stage of T. spiralis infection does not affect the protective efficacy of HBV vaccination, while the enteric phase results in a reduced immune response to HBsAg. The finding has important clinical relevance to the immunization strategies against HBV in highly T. spiralis-endemic areas.
Footnotes
Acknowledgment
This work was supported by the National Natural Foundation of China (project grant number 81000739). The research performed in this study was in compliance with the laws of China and the authors' respective institutions.
Disclosure Statement
No competing financial interests exist.