
Abstract
Despite the importance of feedstuffs and feed ingredients in the food chain, few studies have analyzed the effectiveness of usual methods for the microbial decontamination of feeds. This work aimed to study the combined effect of temperature and organic acids (formic or lactic) on the inactivation of 10 isolates of Salmonella enterica and Escherichia coli obtained from vegetable feed ingredients, including cereals and other processed materials, in cattle feed. A central composite design was used with acid concentrations ranging from 0 to 0.2% (vol/wt) and temperatures from 50°C to 70°C. Second-order models were obtained to predict the survival inhibitions. The results reported in the present study indicate that temperature affects the survivability more significantly than acid concentration, whose effect was isolate dependent. However, after 2 min of treatment, optimal microbial inhibitions were generally achieved at temperatures above 65°C, using 0.1% formic acid or 0.2% lactic acid for Salmonella isolates and around 0.1% lactic acid or 0.2% formic acid for E. coli isolates, respectively. This work provides a set of guidelines helpful to reduce microbial contamination of feeds and highlights the importance of feed-ingredient monitoring to reduce pathogen contamination risk during feed processing.
Introduction
Different approaches have been taken to minimize contamination with pathogenic microorganisms in the farm environment, from preharvest control strategies (Sargeant et al., 2007) to risk assessment of farm management practices (Van Immerseel et al., 2009). Among the wide number of decontamination methods available for feedstuffs, heat treatments are one of the most effective. During animal feed manufacturing, heat treatments occur either at conditioning, and/or during the pelleting process (Hutchison et al., 2007). Retention times of feed mash depend on the formulation and mass of feed, final temperature, and equipment used (Hutchison et al., 2007). Times of heat treatment from<1 min at 71.1°C (Matlho et al., 1997) to 3.5–4.0 min at 78°C–80°C (Blank et al., 1996) have been reported for decontaminating poultry feed.
Although heat treatments are usually effective, combinations of time/temperature for commercial feed production can be insufficient to eliminate high numbers of microorganisms (Maciorowski et al., 2006; Hutchison et al., 2007). Since feeds are nonsterile (Jones and Richardson, 2004) and sensitive to postprocessing recontamination (Maciorowski et al., 2006), usually more than one strategy is used to prevent microbial growth in feedstuffs.
Organic acids have been traditionally used as preservatives due to their antimicrobial activity. Although their antibacterial mechanism is not fully understood, it has been extensively studied (Giotis et al., 2007) and appears to be due primarily to the undissociated acid form interfering with cell metabolism (Cherrington et al., 1990). Among organic acids, short-chain fatty acids such as formic (Al-Natour and Alshawabkeh, 2005) and propionic (Ha et al., 1998) have been investigated as bactericidals in feed and feed ingredients. The antimicrobial effects of organic acids can occur during feed processing, prevent recontamination during storage (Wales et al., 2010), or exert their antimicrobial activity in the animal gastrointestinal tract (Cherrington et al., 1991).
The objective of this work was to study the combined effect of temperature and acid concentration on microbial inactivation in cattle feed, using response surface methodology. We studied the behavior of 10 isolates of Salmonella and E. coli from different vegetable feed ingredients in acidified feeds with formic or lactic acid, at temperatures between 50°C and 70°C.
Materials and Methods
Bacterial strains
Bacteria were isolated from vegetable feed ingredients: wheat bran (slSAL-1, ecSAL-3), corn (slMAZ-1, ecMAZ-4), soybean meals 47% (slSJ7-1, ecSJ7-1, ecSJ4-2), cottonseed (slALG-1), corn dry distillers grains with solubles (DDGS) (slDDM-1), and barley (ecCEB-1). Detection and isolation were performed using the ISO methods for Salmonella (ISO 6579: 2002) and E. coli (ISO 4831:2006 and 4832:2006) detection in food and animal feed. Biochemical confirmative tests were performed following preliminary identification of colony morphology on selective media. Isolates were preserved as frozen stocks at −80°C in Tryptic Soy Broth (TSB; Cultimed Panreac Química S.A., Barcelona, Spain) containing 30% (vol/vol) of glycerol, and propagated twice before use. Cultures were grown in Erlenmeyer flasks containing 50 mL of TSB at 37°C for 12 h.
Preparation of acidified contaminated feed
An acid-free pelleted cattle feed (Table 1) supplied by a local commercial manufacturer (COREN S.C.L., Ourense, Spain) was utilized in this work. On arrival, each feed batch was assessed for both Salmonella and E. coli using the ISO methods indicated above that confirmed no Salmonella nor E. coli were present in any of the batches analyzed. The commercial feed utilized (Table 1) was milled using a laboratory mill (IKA-Werke GmbH & Co. KG, Staufen, Germany) with a 3-mm sieve and sterilized by autoclaving (105°C, 5 min). Formic or lactic acid solutions were sprayed in a 1% (vol/wt) to achieve concentrations from 0.02 to 0.2% (vol/wt) (BASF, Ludwigshafen, Germany). Cultures were centrifuged (13,000×g, 10 min, 4°C), and cells were resuspended in phosphate-buffered saline (PBS) and added at 2% (vol/wt) to the feed. The number of cells in the contaminated feed was determined before heat treatment and was of 1×105 to 1×106 CFU/g for all assayed conditions and isolates. Both acid solutions and cultures were sprayed and then agitated end-over-end in a 1.5-L plastic beaker for 4 min, as previously optimized. Control samples with identical moisture levels (3%) were used to monitor the time required to achieve the selected temperatures.
Premix contains per kg: vitamin A 104 IU, vitamin D3 2×103 IU, and vitamin E 10 mg.
DDS, dry distillers solubles.
Thermal inactivation
Disks utilized in this work were similar to those described by Jin et al. (2008). However, in this article, thermal-death-time disks were reusable devices with a cavity of 4.5 mm height (i.e., 28% thicker than those utilized in the present work [3 mm]). Furthermore, our single-use disks consisted of a flat rubber o-ring completely sealed by two layers of 0.015-mm-thick aluminum, whereas those of Jin et al. (2008) were approximately 2 mm thick. Under these conditions, heat transfer is highly favored, being the time required to achieve the selected temperatures of 26 s on average. Devices were filled with 1 g of feed and submerged in a water bath at temperatures shown in Table 2, including a noninoculated sample to monitor each assayed temperature. After a 2-min treatment, triplicate samples were removed, cooled, and sanitized with sodium hypochlorite. After rinsing, the feed was homogenized with 10 mL of PBS to obtain the count suspension (Sc
). Serial dilutions were made from Sc
, 0.1 mL plated in duplicate in tryptic soy agar, and incubated at 37°C for 48 h. Enumeration was also performed before heat treatment, and the inhibition of survival was calculated as follows:
where I is the bacterial inhibition (% of CFU/g), N0 is the initial number of cells (CFU/g), and Nt is the number of survivors (CFU/g) after the heat treatment.
Experimental design and statistical analysis
Bacterial inhibition was studied using a second-order central composite orthogonal design (α=1.267), with quintuple replication in the center of the factorial design, but not in other points of the experimental domain (Akhnazarova and Kafarov, 1982; Box et al., 2005). The range of independent variables, temperature (T), and acid concentration (A) is shown in Table 2.
From the data, empirical equations were obtained describing the significant parameters in relation to temperature and acid concentration. The general form of the second-order polynomial equation is:
where I is the bacterial inhibition (% of CFU/g), b0 is the constant coefficient, bi is the linear effect coefficient, bij is the interaction effect coefficient, bii are the squared effect coefficients, and xi and xj represent the independent variables (T and A).
The goodness-of-fit was established as the adjusted determination coefficient, the statistical significance of the coefficients verified by means of the Student t-test (α=0.05), and the model consistency by the Fisher F test (α=0.05) using the following mean-squares ratios:
Deriving the equations with respect to T and A and equaling the expression to zero can calculate maximum values for each variable (Tm and Am ), while optimum inhibition (Im ) is calculated replacing Tm and Am .
Results
A central composite orthogonal design was chosen to optimize both variables. The upper limits of temperature and acid concentration were selected not exceeding the usual values of commercial feed nor affecting dry matter intake by the animals. The experimental domain (−1.267, 1.267) was 0–0.2% (vol/wt) of acid and 50°C–70°C (Table 2). Considering a possible synergistic effect between variables, their optimum levels would provide a set of safety values ensuring a feasible treatment to be implemented in the food industry. The time of heat treatment was fixed to 2 min, which has been reported as enough to achieve a significant reduction in the feed microbial population (Matlho et al., 1997; Hutchison et al., 2007).
Combined effect of formic acid and temperature
Bacterial inhibition was extremely isolate-dependent. Nonetheless, four patterns of behavior could be defined (Table 3 and Fig. 1):
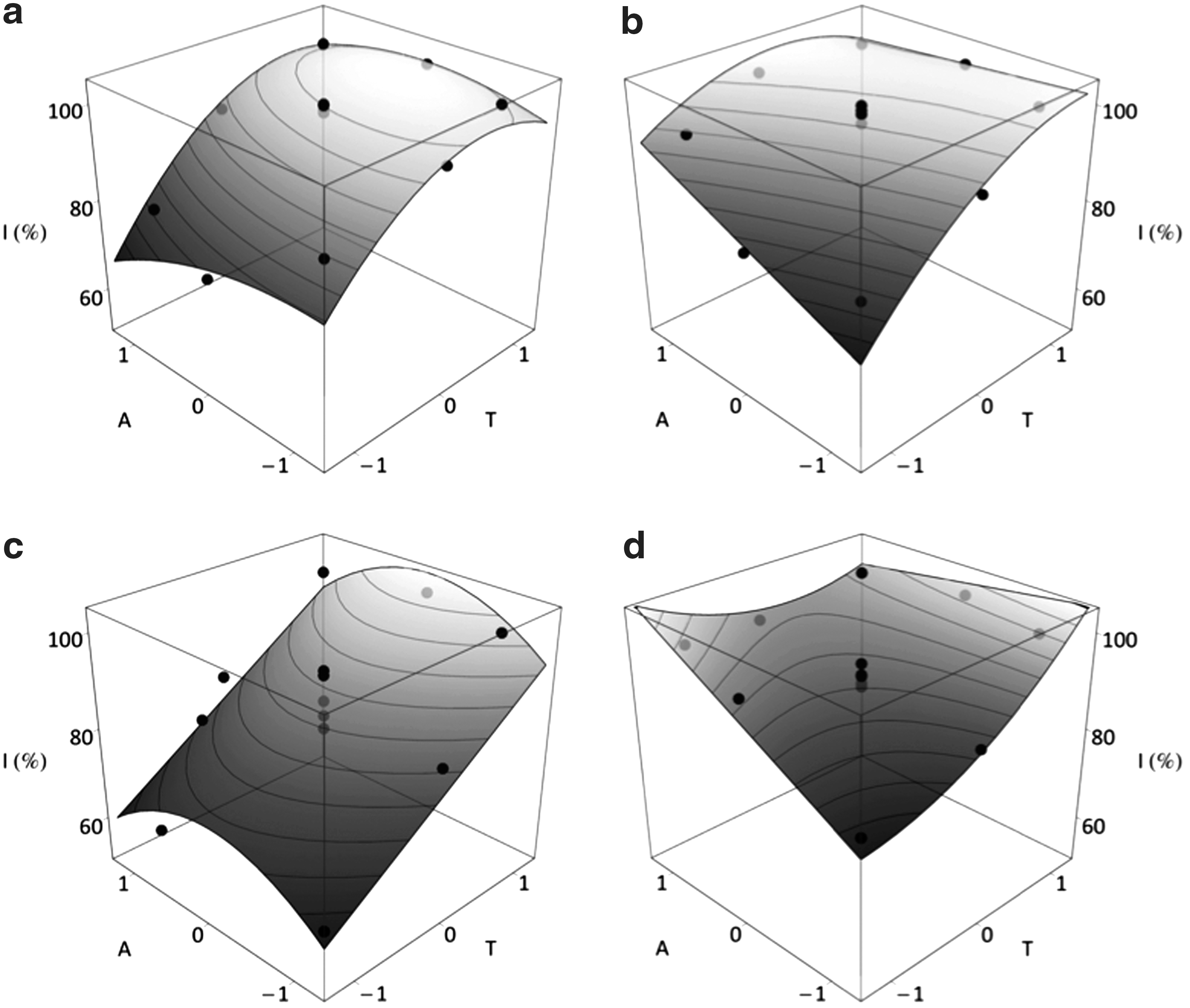
Selection of response surfaces for bacterial inhibition (I) as a function of formic acid (A) and temperature (T), according to the equations described in Table 3.
Response surface presents a saddle point. An absolute maximum cannot be calculated, so optimal values correspond to local maxima within the experimental domain (see text for more details).
1. Both variables showed a negative quadratic coefficient.
2. Only temperature revealed a quadratic effect with negative coefficient.
3. Only acid concentration had a negative quadratic term.
4. Only temperature showed a significant quadratic effect with positive coefficient.
Behavior 1
This was the major response, since half of the assayed isolates (slMAZ-1, slSJ7-1, slALG-1, ecCEB-1, and ecMAZ-4) showed this behavior. Empirical models were characterized by negative coefficients for quadratic variables, suggesting a saturation of the response at high values of T and A, or the existence of optimum values within the experimental domain. In fact, predicted temperatures for optimal antimicrobial effects were located close to the maximum levels assayed, being always higher than 65°C. By contrast, optimal formic concentrations were generally observed at intermediate concentrations of acid (Fig. 1a).
Except for slSJ7-1, empirical equations had interaction coefficients, so a different formic acid effect was observed depending on treatment temperature. However, the interaction sign was isolate dependent, obtaining (when this term was positive) maximal inhibition of one variable for maximum value of the other (Fig. 1a). This interaction was highest for ecMAZ-4, since the coefficient (TA) was close to those of the linear variables T and A. On the other hand, the negative interaction in slMAZ-1 indicates that at high temperature an increase in formic concentration does not produce remarkable effects on the response, whereas at low temperature a clear inhibitory effect is revealed. The lack of interaction term generates a symmetrical response surface where each variable has identical effect for high and low levels of the other one.
Behavior 2
The behavior of isolates slDDM-1 and ecSAL-3 is characterized by empirical models including a quadratic term for temperature and a linear dependence of inhibition with formic acid concentration. Equation obtained for slDDM-1 isolate showed that formic acid had no effect, although the response surface of ecSAL-3 revealed a clear inhibitory effect of formic acid at low temperatures (Fig. 1b).
Behavior 3
The response of slSAL-1 described a positive linear effect of T and a negative quadratic effect of A (Fig. 1c). The absence of interaction term leads to rising ridge, a convex surface with a line of maxima at 0.1% A but without an absolute maximum response within the experimental domain. However, in all cases the inhibition increases with temperature, reaching a 100% inhibition at 0.1% A and 70.1°C.
Behavior 4
Two E. coli isolates from soybean meal (ecSJ7-2 and ecSJ4-2) showed a different experimental tendency. Although empirical equations included quadratic terms for temperature, they had positive sign leading to slightly concave response surfaces (Fig. 1d). In ecSJ7-2, the concavity observed by effect of T and positive influence of A, produced a softly shift of the optimal values of both variables towards their maximum level (100% inhibition at 70.1°C and 0.2% A). At any level of A, lower inhibitions were observed at 55.2°C, with a minimum at 55.2°C and 0% A. The response surface model for ecSJ4-2 presents a saddle point, meaning an absolute maximum response cannot be obtained within the experimental domain. However, a negative interaction term and positive linear coefficients for T and A cause a response increasing with both T and A. Maximum inhibitions were obtained in the response surface limits (Tm =70.1°C, Am =0%) and (Tm =50.1°C, Am =0.2%). From a practical viewpoint, these results indicate that similar levels of inhibition could be obtained when feed is only treated by heat or acid conditions.
Combined effect of lactic acid and temperature
Less variability was observed when lactic acid was used as antimicrobial agent, leading to different behaviors according to the three mathematical models (Table 4 and Fig. 2):
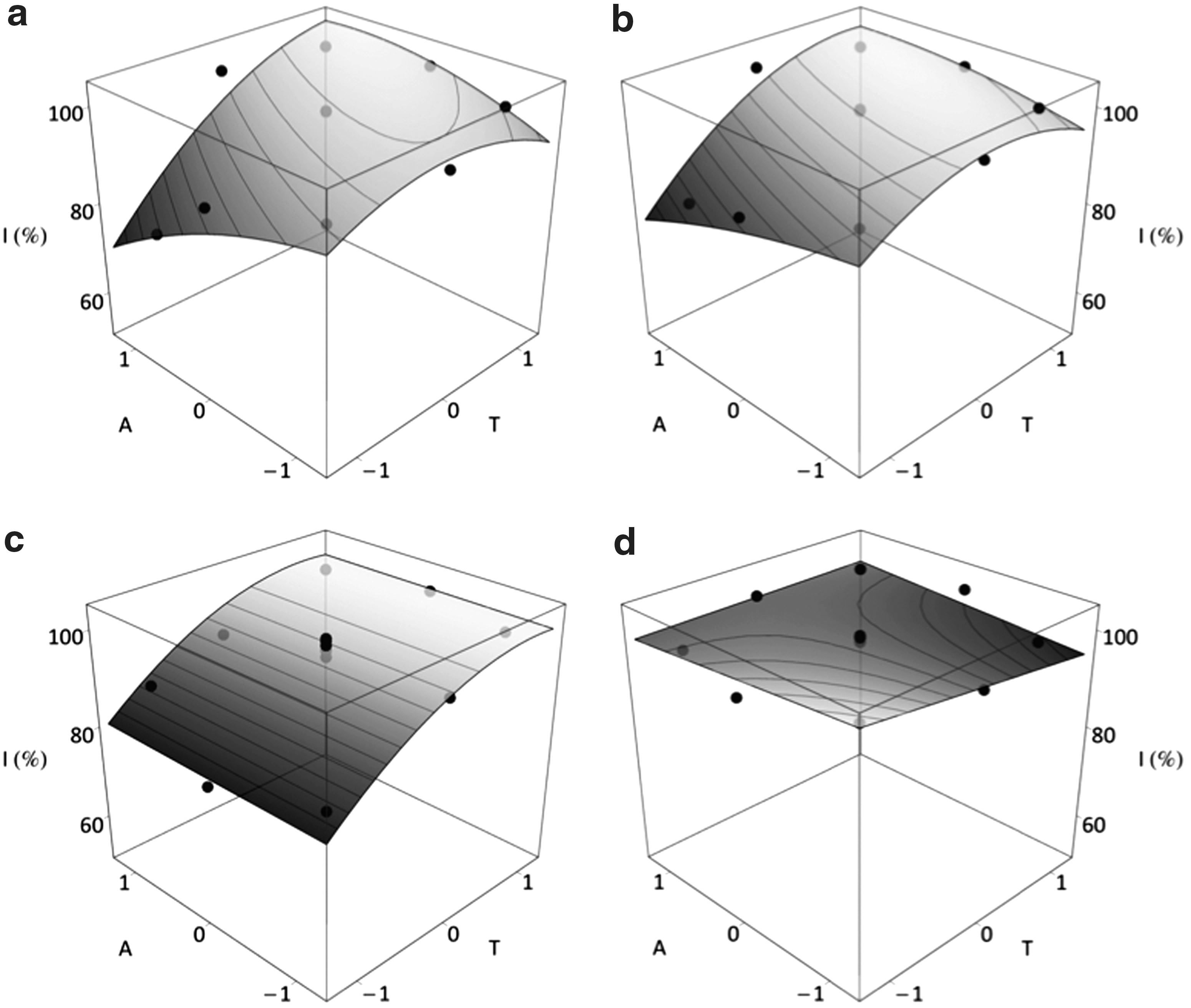
Selection of response surfaces for bacterial inhibition (I) as a function of lactic acid (A) and temperature (T), according to the equations described in Table 4.
These values are theoretical maxima within the experimental domain (see text for more details).
Response surface presents a saddle point. An absolute maximum cannot be calculated, so optimal values correspond to local maxima within the experimental domain (see text for more details).
1. Both variables had second-order terms.
2. Only the temperature showed a quadratic coefficient.
3. Equation without quadratic terms.
Behavior 1
The major behavior observed included quadratic terms for both variables, although the sign of the quadratic coefficients was not always negative. Salmonella (slALG-1 and slDDM-1) and E. coli isolates (ecCEB-1, ecSJ7-2 and ecMAZ-4) showed inhibition surfaces (Fig. 2a and b) similar to those of formic acid, but with a higher inhibitory effect (Fig. 2a). For Salmonella isolates, the maxima of both variables were predicted out of the experimental domain (Tm >73°C and Am >0.21%), although the combinations T=70.1°C and A=0.17% for slDDM-1, and T=70.1°C and A=0.19% for slALG-1 provided inhibitions only 0.2% lower than the maxima. However, clear optima were calculated for E. coli isolates: 65.8°C and 0.137% (ecCEB-1) and 66.8°C and 0.14% (ecMAZ-4).
The negative interaction term and the difference between linear and quadratic coefficients in ecSJ7-2 resulted in an Im (112.9%) at high temperature (116.9°C). Nonetheless, this result should be considered as a mathematical artefact since the maximum acid concentration value was negative (Am =−0.102%).
Equations for wheat bran isolates (ecSAL-3 and slSAL-1) had positive quadratic terms for lactic acid. In slSAL-1, the effect of T was higher than A, whereas for ecSAL-3 both contributions were similar. Antimicrobial optimal conditions were 67°C, 0.17% and 64.5°C, 0.1% for slSAL-1 and ecSAL-3, respectively.
Behavior 2
This group included two isolates (slMAZ-1 and slSJ7-1) in which only the temperature had influence on survival inhibition. For both bacteria, there was an increased inhibition with increasing temperature until a temperature optimum: 67°C for slMAZ-1 and 68.2°C for slSJ7-1 (Fig. 2c).
Behavior 3
Empirical equation for ecSJ4-2 did not present second-order coefficients for T and A nor a linear term for A, which is only present in a positive interaction term (Fig. 2d). In this case, the low value of the coefficients determined minor effects on the response, with inhibitions near 100% within the entire experimental domain.
Discussion
Although microbial inactivation in cattle feed has been scarcely investigated, some studies have analyzed bacterial survival either under thermal treatments (Blank et al., 1996; Hutchison et al., 2007) or acidic conditions (Ha et al., 1998; Al-Natour and Alshawabkeh, 2005). Nevertheless, some reports suggest that heat treatment alone may be insufficient to eliminate high numbers of microorganisms (Maciorowski et al., 2006; Hutchison et al., 2007) or that under certain circumstances, organic acids can promote microbial growth in feeds (Lynn et al., 1998). Since acidifiers are usually added to raw materials and finished feeds, it is a key factor to know whether they ensure significant reductions in the microbial load at typical feed-manufacturing temperatures. The most efficient methodology to evaluate the combined effect of two factors at the same time is the use of response surface methodology (RSM), allowing the prediction of optimal values of the independent variables and minimizing experiments.
In this work, a central composite orthogonal second-order design (Akhnazarova and Kafarov, 1982; Box et al., 2005) was used to study the combined effect of temperature and organic acids on the inactivation of foodborne isolates in cattle feed. The obtained models can be considered good predictors for the experimental data, since the equations are consistent according to the values of mean-squares ratios (F1
, F2
, F3
, and F4
). The adjusted determination coefficients (
According to the response surfaces obtained, bacterial inhibition was primarily affected by heat. These results reveal that when using acidification as the sole feed treatment, longer storage times are needed to achieve significant reductions in the microbial load. In fact, Al-Natour and Alshawabkeh (2005) reported a 2 log-units decrease of Salmonella enterica Gallinarum in broiler feed treated with formic acid (0.5%–1.5%) after 7 days. Even minor reductions (<1 log-unit) in Salmonella Typhimurium counts were observed in soybean meal–based poultry mash treated with 1% buffered propionic acid after the same storage time (Ha et al., 1998).
Despite heat being the variable that most influenced bacterial inhibition, significant interactions were generally observed, allowing the use of lower temperature and antimicrobial agent than when these treatments are used alone. Nevertheless, synergistic interactions between heat and antimicrobial acids did not always occur, with negative interaction terms primarily in the presence of formic acid. These results, leading to lower inhibitions than expected at high temperatures and acid concentrations, could be due to the high volatility of formic acid (Al-Natour and Alshawabkeh, 2005) or to the buffering capacity of some components in the feed matrix (Pursiainen and Tuori, 2008). Similarly, Matlho et al. (1997) did not find interactions between heating time and acid concentration when evaluating the reduction of Salmonella Enteritidis in poultry feed treated with propionic acid (0%–0.2%) at 71.1°C.
Formic acid has greater antimicrobial activity against enterobacteria compared to lactic acid (Ostling and Lindgren, 1993). Its low molecular weight and high hydrophobicity might favor its ability to diffuse freely across the cell membrane, interfering with intracellular pH (Cherrington et al., 1991). By contrast, our results showed that lactic acid was better for the decontamination of cattle feed inoculated with E. coli isolates, since milder conditions were needed to achieve similar inhibitions than using formic acid.
Since the effect of both variables on the inhibition depended largely on the type of acid and isolate, it was not possible to generalize a combination of variables achieving a complete survival inhibition of all isolates. However, process parameters were effectively optimized using RSM, obtaining maximal survival inhibitions (>98%) at temperatures above 65°C in cattle feed acidified with 0.1% formic for Salmonella isolates and 0.1% lactic acid for E. coli isolates after 2 min of treatment. Then, for the same temperature and treatment time, nearly twice the concentration of lactic acid for Salmonella and formic acid for E. coli were required to achieve similar inhibitions. Under these conditions, population declines between 2 and 4 log-units are reached. These results are comparable to those obtained by Matlho et al. (1997), who reported an approximately 10,000-fold reduction in living Salmonella after 80-s heating at 71.1°C in poultry feed acidified with 0.2% propionic acid.
Conclusions
Our results provide a set of guidelines useful to reduce contamination with foodborne pathogens during feed processing, highlighting the importance of ingredient monitoring during animal feed production. We report optimal inhibitions at temperatures above 65°C with 0.1% formic acid or 0.2% lactic acid for Salmonella isolates and around 0.1% lactic acid or 0.2% formic acid for E. coli isolates, respectively. Although these treatments represent significant reductions of target microorganisms (>98%), they should be validated under commercial production conditions.
Footnotes
Acknowledgments
We thank COREN, S.C.L. for their collaboration in the elaboration of this work. This research was financially supported by the Xunta de Galicia, Spain (project PGIDIT03TAL07E). Isabel Rodríguez Amado was funded by a PhD fellowship from the Plan Galego de Investigación, Desenvolvemento e Innovación Tecnolóxica-Incite (2006–2010) (Xunta de Galicia).
Disclosure Statement
No competing financial interests exist.