
Abstract
Cross-contamination via equipment and the food-processing environment has been implicated as the main cause of Listeria monocytogenes transmission. The aim of this study, therefore, was to determine the occurrence and potential persistence of L. monocytogenes in 19 European cheese-processing facilities. A sampling approach in 2007–2008 included, respectively, 11 and two industrial cheese producers in Austria and the Czech Republic, as well as six Irish on-farm cheese producers. From some of the producers, isolates were available from sampling before 2007. All isolates from both periods were included in a strain collection consisting of 226 L. monocytogenes isolates, which were then typed by serotyping and pulsed-field gel electrophoresis (PFGE). In addition, metabolic fingerprints from a subset of isolates were obtained by means of Fourier-transform infrared (FTIR) spectroscopy. PFGE typing showed that six processing environments were colonized with seven persistent PFGE types of L. monocytogenes. Multilocus sequence typing undertaken on representatives of the seven persisting PFGE types grouped them into distinct clades on the basis of country and origin; however, two persistent strains from an Austrian and an Irish food processor were shown to be clonal. It was concluded that despite the fact that elaborate Hazard Analysis and Critical Control Point concepts and cleaning programs are applied, persistent occurrence of L. monocytogenes can take place during cheese making. L. monocytogenes sanitation programs could be strengthened by including rapid analytical tools, such as FTIR, which allow prescreening of potentially persistent L. monocytogenes contaminants.
Introduction
L
Having colonized a food-processing environment, L. monocytogenes can spread and consequently recontaminate the finished products (Ferreira et al., 2011; Fox et al., 2011). It was concluded that unhygienic design of equipment, inadequate hygiene measures, niche adaption, and biofilm formation may lead to cross-contamination with the pathogen (Keto-Timonen et al., 2007; Da Silva and De Martinis, 2013). The existence of persistent pulsed-field gel electrophoresis (PFGE) types of L. monocytogenes is usually demonstrated by applying molecular tools, such as PFGE and amplified-fragment length polymorphism. Performing genotyping methods, however, is usually laborious and requires technical expertise. The recognition of enduring L. monocytogenes colonization could be strengthened by a rapid screening tool, applicable for the identification and typing below the species level. The aim of this study was (1) to analyze the contamination with L. monocytogenes in 19 cheese-producing facilities located in three European countries; (2) to establish a strain collection based on persistent PFGE types; and (3) to test the applicability of Fourier-transform infrared (FTIR) spectroscopy as a rapid screening tool, for monitoring potential persistence of L. monocytogenes in cheese-processing facilities.
Materials and Methods
Sampling
The occurrence of L. monocytogenes in 19 mostly traditional cheese-processing facilities in the Czech Republic, Austria, and Ireland was determined in a sampling approach followed during January 4, 2007–May 31, 2008. Companies in Austria and the Czech Republic were industrial producers, whereas small farmhouse cheese makers were sampled in Ireland. The sampling protocols included environmental sampling external to the processing facility in cases where a transmission from the preharvest area was plausible (farmhouse cheese-makers located in Ireland). In Ireland, 607 samples were taken at six farmhouse cheese-making facilities from the farm environment (including water, soil, feces, bedding, and feed), whole milk, brine, cheese, and environmental swabs. Eleven Austrian dairies (producing acid curd cheese, ripened soft cheese, blue-veined, and semihard or hard cheeses) were sampled, resulting in 322 samples. Cheese samples, environmental swabs, drain water, and product-associated samples (smear, brine, drain water, and wash water) were collected according to the method of Asperger et al. (2001) (Supplementary Fig. S1; Supplementary Data are available online at
L. monocytogenes strain collection
In total, 226 L. monocytogenes isolates were available for the creation of a strain collection. The L. monocytogenes collection contained strains isolated during the sampling approach that lasted from 2007 to 2008 (50 isolates from Ireland, two isolates from Austria, 46 isolates from the Czech Republic). In all participating institutions, recovered isolates from the same cheese producer were stored and compared to isolates collected in the years before the basic sampling event (one isolate from Ireland, isolated in 2000; 116 isolates from Austria, recovered from 1992 to 2007; 11 isolates from Czech Republic, isolated from 1999 to 2007) (Supplementary Table S1).
Microbiological methods and polymerase chain reaction (PCR) confirmation of isolates
The cheese- and production-associated samples were analyzed according to ISO 11290-1 (1996). For the detection of L. monocytogenes, 25 g of solid sample were homogenized and enriched in 225-mL Half-Fraser Broth (Merck KGaA, Darmstadt, Germany). After incubation for 24 h at 30°C, 0.1 mL of the primary enrichment was transferred to 10-mL Fraser Broth (Merck KGaA) and incubated for 48 h at 37°C. Ten microliters was inoculated onto Agar Listeria according to Ottaviani and Agosti (ALOA; Oxoid Ltd., Basingstoke, UK) and Palcam agar (Merck KGaA). Selective agars were incubated for 24–48 h at 37°C. In Austria, cheese and product-associated samples were investigated according to the protocol published by Asperger et al. (2001) (Supplementary Fig. S1). Further subcultivation and plating steps, selective media, incubation times, and temperatures were applied following the ISO 11290 protocol. Colonies suspected as L. monocytogenes on ALOA (Oxoid Ltd.) and Palcam agar (Merck KGaA) were confirmed by PCR detection of the hly gene (Border et al., 1990), which encodes the virulence factor listerolysin O, and the highly conserved 23S rDNA gene of Listeria spp. (Border et al., 1990; Rodriguez-Lazaro et al., 2004).
Pheno- and genotyping
Classical serotyping against the somatic (O) (and on occasion the flagellar [H]) antigens was determined by the scheme established by Seeliger and Hohne (1979), using the Listeria antisera set (Denka Seiken, Japan). Subsequently, the serotype was further defined using a multiplex PCR developed by Doumith et al. (2004). The sample preparation for the infrared absorbance measurements was performed as described by Oberreuter et al. (2002). Fourier transformation, calculation of derivatives, and normalization was performed with the OPUS software version 5.5 (Bruker, Germany). Hierarchical cluster analysis (HCA) of the spectra was performed as described previously (Rebuffo et al., 2006) using the Ward's algorithm of the OPUS software and the second derivative of the original spectra as input in the regions ranging from 700 to 1200, 1500 to 1800, and 2800 to 3100/cm.
DNA macrorestriction digest of L. monocytogenes strains with AscI and ApaI was performed according to the PulseNet protocol (
Results
In Ireland, a total of 607 samples along the cheese production chain, from farm to finished product, were analyzed from six cheese-processing facilities. The samples were collected from the farm environment (n=33), from unpasteurized milk (n=6), brine (n=16), food production environment swabs (n=447), and cheese products (n=105). The 13.5% L. monocytogenes confirmed positive samples originated from three facilities.
The sampling in the Czech processing facilities A and B resulted in 16.1% (n=205) L. monocytogenes positive environmental swabs. Thirteen of 200 cheese samples (6.5%) were confirmed positive for L. monocytogenes. Most of them (4.5%, n=9) originated from a producer of blue-veined cheese. In Austria, 322 samples were collected in 11 dairies consisting of drain water (n=55), smear/brine/washing water (n=254), milk samples (n=5), and cheese products (n=8). Two hard cheese samples were positive for L. monocytogenes.
Serotyping of the 226 isolates revealed that the majority of isolates were classified as serotype 1/2a (67.5%) followed by serotype 4b (17.6%), 1/2c (8.8%), 1/2b (3.5%), and 3b (0.4%). Five isolates were not typeable. The rarely found serotype 1/2c was exclusively isolated from an Irish facility that was intensely sampled. Furthermore, 171 isolates were assigned to genetic Lineage II (1/2a, 1/2c) and 50 isolates were assigned to Lineage I (1/2b, 3b, 4b).
The PFGE analysis on 226 L. monocytogenes resulted in 38 (AscI) and 37 (ApaI) profiles. The same ApaI restriction pattern CzA1 was obtained for isolates from both the Czech facilities A and B (Fig. 1), which did not share any raw material, equipment, or geographical proximity. Colonization with L. monocytogenes for more than 1 year could be observed in seven cheese-processing facilities (36.8%, n=19). Five L. monocytogenes PFGE types were recurrently isolated for at least 2 years from Austrian producer A, B, and C (Supplementary Table S1). By including the set of stored isolates, we found evidence for some very long-lasting contamination periods. One L. monocytogenes PFGE type (PFGE type AuA2) was re-isolable over a period of 11 years. Two genetically indistinguishable isolates (PFGE type IrlA1) from an Irish dairy were recovered at a time distance of 8 years. A high genetic similarity (90%) was found between isolates of Austrian dairy B and the Irish dairy A, both producing smear-ripened cheeses. Two L. monocytogenes 1/2a PFGE types (CzA1, CzB1) could be isolated within a timeframe of 5 years.
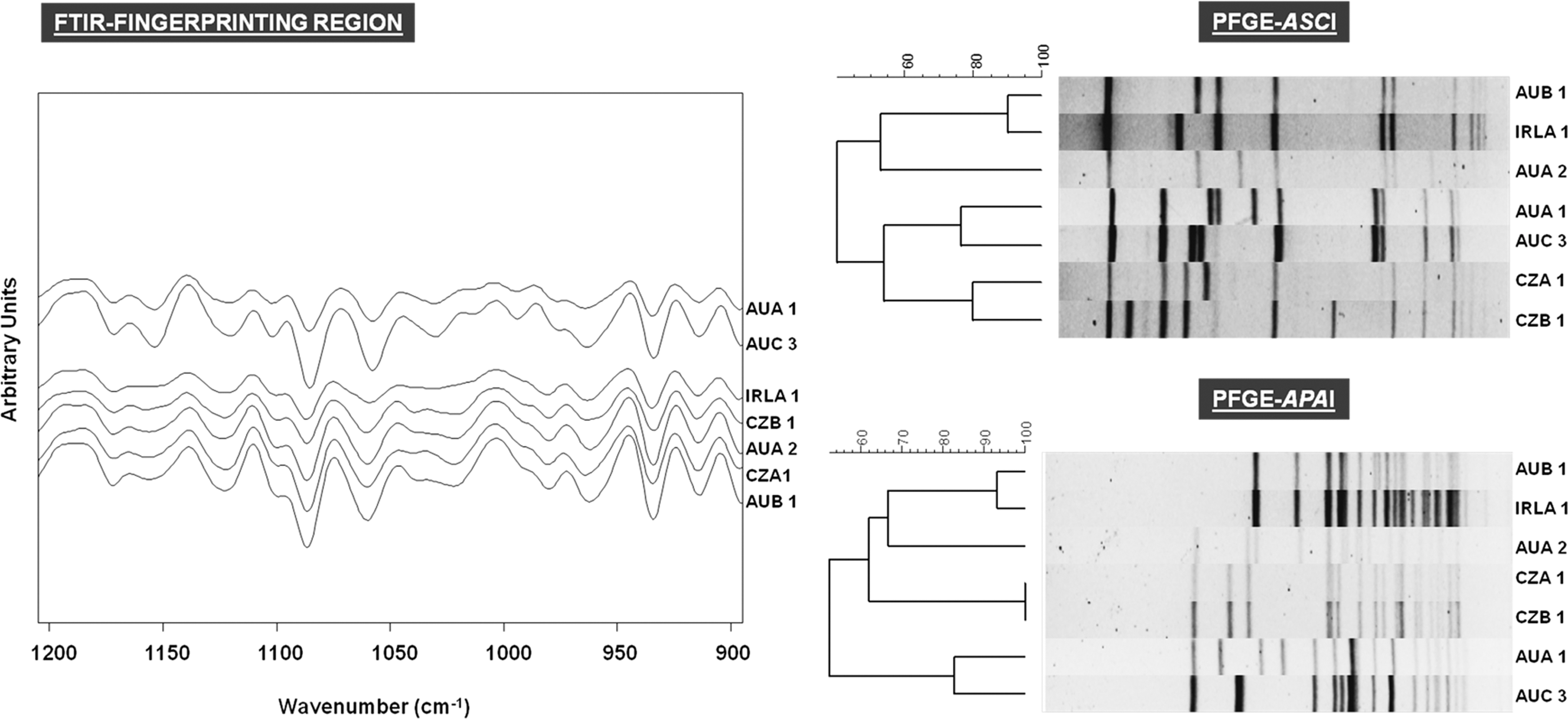
Seven representative persistent Listeria monocytogenes pulsed-field gel electrophoresis (PFGE) types from six European dairies including a comparison of the Fourier-transform infrared (FTIR) fingerprinting region (900–1200/cm) and the PFGE cluster analysis (restriction enzymes AscI and ApaI). Abbreviations for representative L. monocytogenes persistent PFGE types are located to the right of each figure part: AU, Austria; IRL, Ireland; CZ, Czech Republic.
The HCA on a selection of 58 L. monocytogenes strains using second derivate FTIR spectra revealed three major clusters (Fig. 2; Supplementary Table S1). Cluster I comprises 32 L. monocytogenes isolates of different geographic origin, belonging to serogroup 1/2 and 3. The second cluster includes 14 Czech L. monocytogenes isolates from facilities A and B (serogroup 1/2). Finally, the third cluster includes 10 isolates from Austrian facilities A and C and four isolates from Irish manufacturers (mainly isolates of serovar 4b). As depicted in Figure 1, variations in the polysaccharide region of the FTIR spectra significantly contributed to the distinct grouping of serovar 4b strains (AuA1 and AuC3).
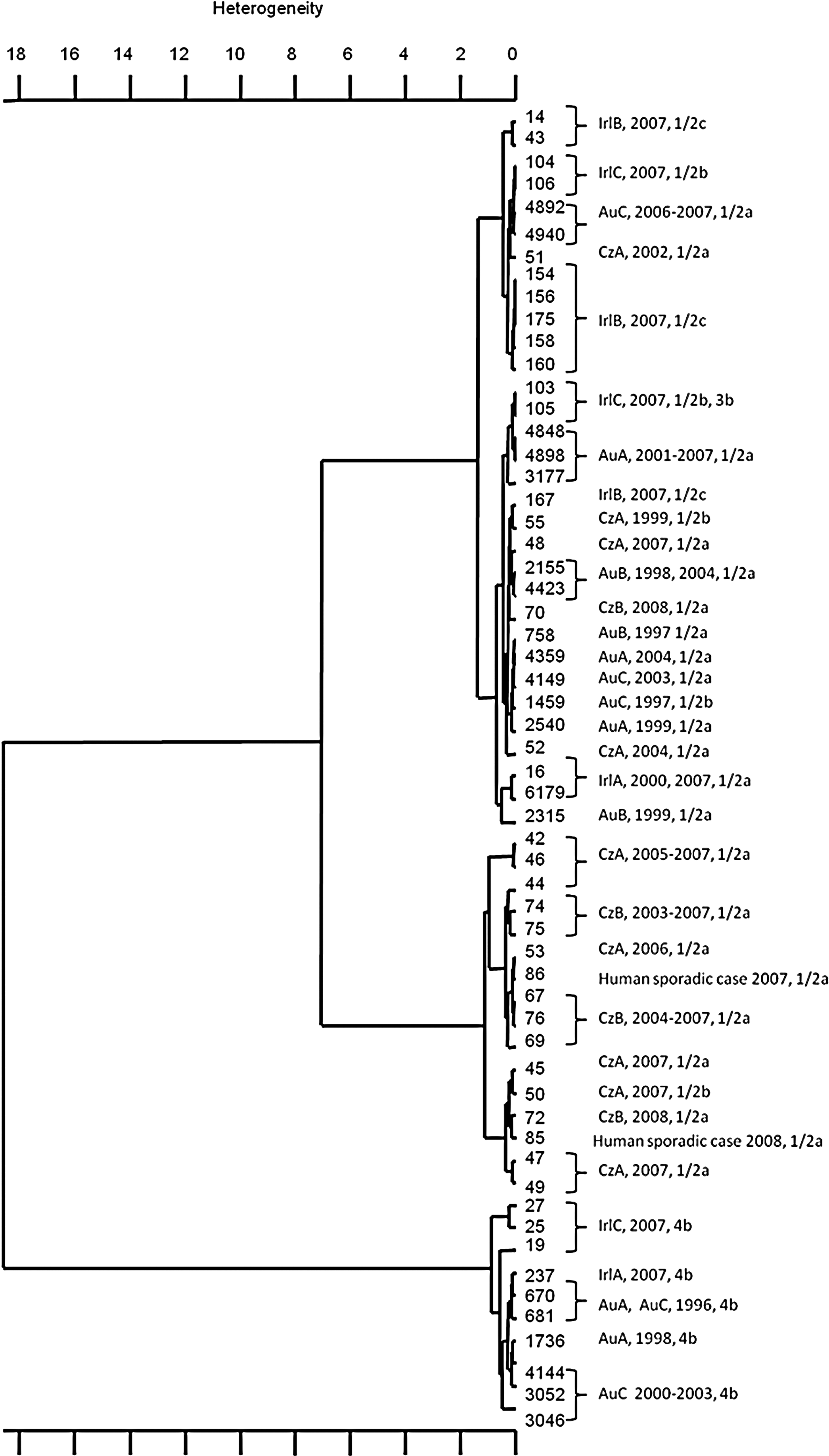
Fourier-transform infrared hierarchical cluster analysis using the Ward algorithm on Euclidian spectral distances of 58 Listeria monocytogenes isolates from the cheese-processing environment. Strain and producer code, date of isolation, and serovar are listed beside the Hierarchical Cluster Analysis.
MLST typing of representatives of the seven persistent L. monocytogenes PFGE types resulted in two strains, AuC3 (isolated during 2000-2003) and AuA1 (isolated during 1996–1998), corresponding to ST 1. Two Czech L. monocytogenes strains (CzA1 and CzB1) were both ST 204 (isolated during 2003–2008). The L. monocytogenes strain AuA2 (1996–2007) corresponded to ST 14. The high similarity in the PFGE analysis for IrlA1 (Irish strain isolated through 2000–2008) and AuB1 (Austrian strain isolated through 1997–2004) was confirmed by MLST typing since both persistent PFGE types were ST 121. SplitsTree analysis of L. monocytogenes ST 14, 121, and 204 in comparison to 16 STs of genetic Lineage II, serovar 1/2a sharing more than four housekeeping genes (available from:
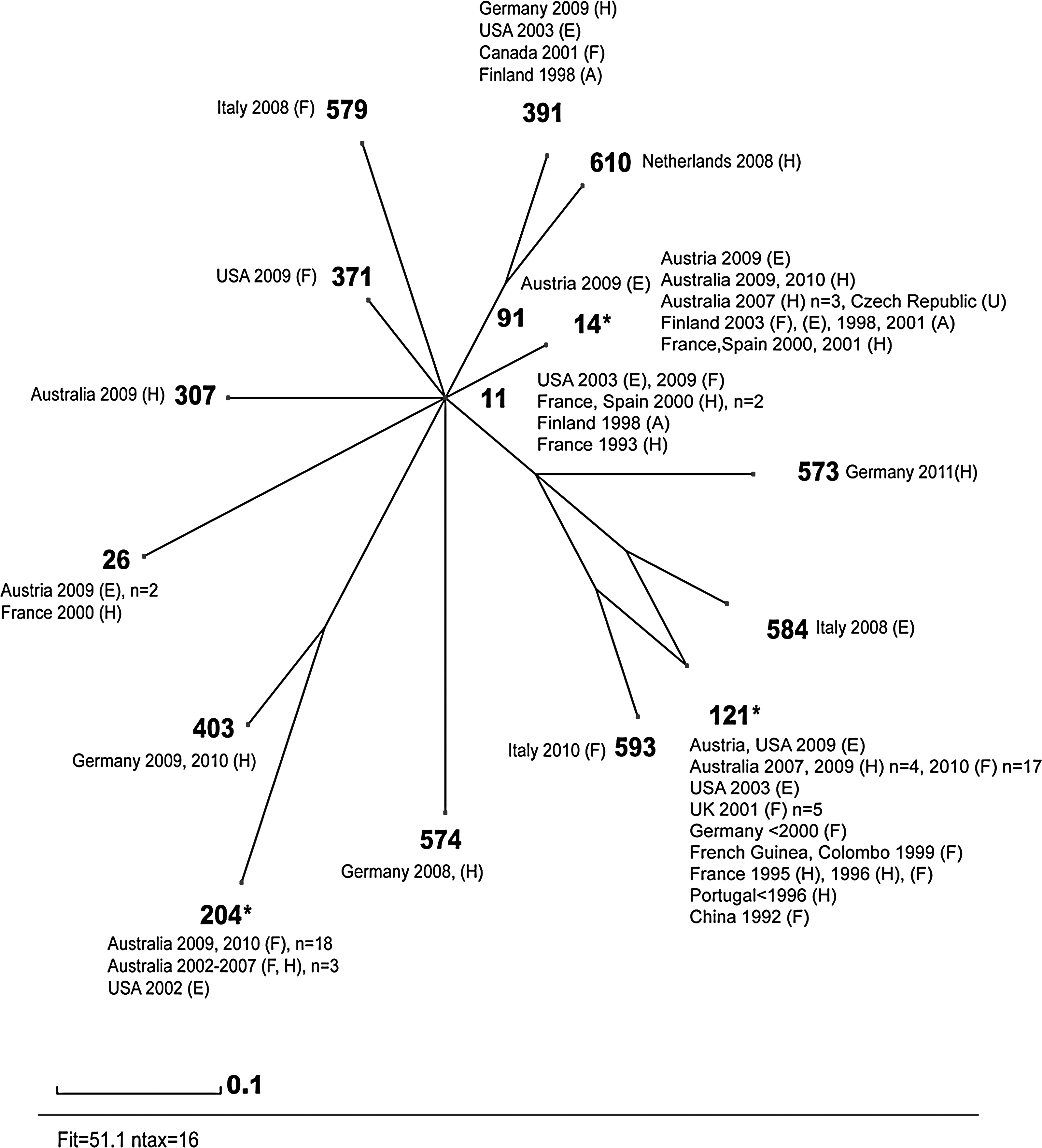
Genetic relationships of Listeria monocytogenes isolates from this study based on multilocus sequence typing (MLST). Phylogenetic relation of L. monocytogenes ST14, 121, and 204 (each marked with an asterisk) is shown in comparison to 16 sequence types (ST) of genetic Lineage II, serovar 1/2a, including abcz allelic types 5, 7, and 8, available from the MLST database of Institute Pasteur. The graphic is a split decomposition analysis based on allelic profiles by using a PubMLST software tool written by Huson (1998). Each number represents a multilocus sequence typing genotype (ST). Isolates were selected for the comparison within a timeframe (1992–2011). The abbreviations for the source as food, factory (F), human (H), animal (A), environment (E), unknown (U), and the number of isolates with more than one representative are listed after the date of isolation.
Discussion
Persistence of genetically indistinguishable L. monocytogenes strains over a long time period is a well-described contamination scenario in food processing (Keto-Timonen et al., 2007; Ferreira et al., 2011; Holch et al., 2013). This study is the first that focused on the occurrence and potential persistence of L. monocytogenes across a whole food category, since 19 producers of traditional Irish, Czech, and Austrian cheese products were included. L. monocytogenes persistence was a widespread epidemiological event in the investigated cheese-processing facilities. The reasons for long-time persistence are still unknown (Ratani et al., 2012; Da Silva and De Martinis, 2013; Holch et al., 2013). Contamination with L. monocytogenes could be traced back for more than a decade, but the niche of in-house strain survival was unknown. It is likely that they will remain discrete, since even in outbreak investigations, the source is identified in only one third of the cases (EFSA and ECDS, 2013).
Serotyping results showed serotype 1/2a as being the most prevalent L. monocytogenes serotype in the cheese-processing facilities analyzed in this study. These findings are in accordance with previous reports (O'Connor et al., 2010). Until recently, serovar 1/2a was not considered to be the main contributor to the public health burden since the majority of human listeriosis cases were caused by infection with serovar 4b isolates (Kathariou, 2002). A review of the outbreaks over the last 4 years, however, shows the increasing importance of serovar 1/2a, which has caused epidemic listeriosis transmitted by ready-to-eat meat products (Canada), acid curd cheese (Austria, Czech Republic, Germany), Mexican-style cheese (United States), hard cheese (Belgium), and contaminated cantaloupe (United States) (Fretz et al., 2010; Gilmour et al., 2010; Jackson et al., 2011; Laksanalamai et al., 2012; Yde et al., 2012).
MLST typing facilitates a comparison of newly recovered strains to known clonal complexes. Two Austrian dairies harbored the well-described L. monocytogenes ST1 during 1996–2003, which could be linked to epidemic outbreaks (France 1989, Sweden 1995) and is mainly involved in sporadic human cases. Additional persistent dairy strains found in this study were linked to ST 204 (Czech Republic), ST 121 (Austria, Ireland), and ST 14 (Austria). A comparison to the MLST database from Institute Pasteur showed that the majority of ST 204 strains were isolated from food and the environment (soil, water, vegetation). ST 121 isolates are also strongly associated with the food-processing environment and animals but have caused sporadic human infections in Germany, Finland, France, and Australia. ST 14 was involved in human illness, as demonstrated by the Austrian outbreak history of 1986 (Allerberger and Guggenbichler, 1989). Environmental ST 14 representatives were isolated from feed, animals, and in one case from a food factory (Finland 2003).
Due to high discriminatory power, genotyping, including PFGE, is the criterion standard for reference laboratories, but of very limited use for at-line control and rapid contamination management. PFGE is laborious, lacks effective data storage, and does not facilitate subtype data comparisons of multiple origins (Vangay et al., 2013).
FTIR spectroscopy rapidly and cost efficiently reveals profiles with a high throughput capacity that refer to biomolecules from the whole bacterial cell such as polysaccharides, cell wall–associated lipids, cytosolic proteins, and nucleic acids (Rebuffo et al., 2006; Davis and Mauer, 2011). It was found to be suitable for discriminating Listeria species and typing bacteria at subspecies level (Rebuffo et al., 2006; Davis and Mauer, 2011). In this study, FTIR data revealed persisters of a certain origin being interspersed with those of other sources, which indicates that genetically distinguishable persisters have the same or similar metabolic traits. Combining rapid and simple FTIR profiling with technically sophisticated and laborious genotyping is a promising approach, capable of advancing Listeria epidemiological investigations.
It is still unclear whether the persistence phenomenon in L. monocytogenes is genetically encoded or purely a result of better adaptation of some “survivors” to food and environmental factors. Carpentier and Cerf (2011) suggested that hard- or impossible-to-clean sites, cracks in flooring and conveyor belt materials, or corroded spots harbor L. monocytogenes and may cause repeated contamination cycles. A gap in the market of hygienically designed food-processing equipment has been identified by other authors (Keto-Timonen et al., 2007). Persistent contamination could be also explained by bacteria adapting to sublethal concentrations of disinfectants such as benzalkonium chloride and to heavy metals such as cadmium (Ratani et al., 2012). Furthermore, L. monocytogenes survival in biofilms was shown to be influenced by complex and situation-related environmental conditions rather than strain-specific attributes (Nilsson et al., 2011). Genetic studies on L. monocytogenes adaptation to environment and ready-to-eat foods has shown point mutations in the virulence genes, such as premature stop codons in the inlA gene, the presence of inlF genes, and the presence of a prophage in comK (Verghese et al., 2011; Holch et al., 2013).
Whatever drives the persistence phenomena, the current data demonstrate the difficulties in sanitizing and controlling Listeria contamination in the food-processing environment. In this respect, it is a regrettable disadvantage that as yet no rapid method exists to speed up tracing of persistent PFGE types at-line. FTIR could fill this gap to some extent. The data support the opinion that management of critical control points in the processing environment as prescribed in Article 5 of DIR (EC) 2073/2005 (European Commission (EC), 2005) should be more rigorously adhered to.
Footnotes
Acknowledgments
We wish to thank the following for their collaboration during the sampling and characterization period: Sonja Klinger (Vetmeduni Vienna, Austria), Romy Renner (TUM, Technical University of Munich, Freising, Germany), Martina O'Brien and Triona Hunt (Teagasc Food Research Centre, Moorepark). Additionally, we would like to thank the Institute Pasteur for providing the MLST database of L. monocytogenes in the Genotyping of Pathogens and Public Health Platform. This work was supported by the European Union–funded Integrated Project BIOTRACER (contract 036272) under the 6th RTD Framework.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.