
Abstract
Foodborne illness due to Salmonella-contaminated pork products is an important public health problem, causing significant economic losses worldwide. The use of bacteriophages is a potential intervention tool that has attracted interest for the control of foodborne pathogens. The objective of this study was to detect the presence of Salmonella in commercial pig farms and to isolate specific autochthonous bacteriophages against Salmonella Typhimurium, to characterize them and to evaluate their lytic capacity against Salmonella Typhimurium in vivo and in vitro. Salmonella was isolated on 50% (4/8) of the farms, with serotype Typhimurium being the most prevalent, detected in 48.2% of samples (13/27). The isolated Salmonella Typhimurium bacteriophages belong to the Podoviridae family, were active against serotypes Abony, Enteritidis, Typhi, and Typhimurium, but not against serotypes Arizonae, Cholerasuis, Gallinarum, and Pullorum. In in vitro tests, bacteriophage at 107 PFU/mL and 109 PFU/mL significantly reduced (p<0.05) Salmonella Typhimurium counts in 1.6 and 2.5 log10 colony-forming units (CFU)/mL, respectively, after 24 h. Before the in vivo treatment with bacteriophages, Salmonella was identified in 93.3% (28/30) of the fecal samples from the pigs inoculated with 106 CFU/mL, and only in 56.6% (17/30) after the treatment consisting of oral administration of the pool of the bacteriophages after the fasting period, simulating a common preslaughter practice. These results indicate that the pool of bacteriophages administered was capable of reducing the colonization of Salmonella in pigs.
Introduction
F
The use of bacteriophages is an alternative to antibiotics that has been increasingly used on animal production experiments, and it was suggested for prophylactic control and reduction of pathogens (Hagens et al., 2010; Heringa et al., 2010; Santos et al., 2010; Sillankorva et al., 2010; Wall et al., 2010; Callaway et al., 2011; Mahony et al., 2011).
The objectives of the present study were to determine the presence of Salmonella in eight pig production farms, isolate specific autochthonous bacteriophages against Salmonella Typhimurium, and determine their specificity, morphology, survival at different pH levels and bile-salt concentrations, as well as their in vivo and in vitro lytic capacity against Salmonella Typhimurium.
Materials and Methods
Fecal sample collection
Fresh fecal samples (approximately 50 g, n=5 samples/pen) were collected from 8 different commercial finishing farms (n=5 pens/farm; n=25 fecal samples/farm) at Viçosa-MG, Brazil. A total of 200 samples from 200 different animals were collected and immediately placed in sealed individual plastic bags (Nasco Whirl-Pak, Fort Atkinson, WI), maintained on ice, and analyzed within 2 h of collection.
Salmonella isolation
Salmonella was isolated as described by Vico et al. (2011). Twenty-five grams of feces were homogenized (Stomacher 3500 series, Seward Laboratory Systems, Davie, FL) with 225 mL of 1% buffered peptone water (Oxoid, Basingstoke, Hampshire, UK) for 1 min and incubated at 37±1°C for 18±2 h. Samples were selectively enriched by inoculating a 100-μL aliquot of the previously homogenized sample into semisolid Rappaport-Vassiliadis medium (Difco, Sparks, MD) and incubating at 41.5±1°C for 24±2 h. One microliter of each Salmonella-suspect sample was transferred to the selective media XLDA (Oxoid) and BG agar (Oxoid) and incubated at 37±1°C for 24±2 h. Colonies were subjected to biochemical confirmation using triple sugar iron agar (TSI) agar (Difco, USA), iron lysine agar (IL) agar (Difco), and urea broth (Difco). Putative Salmonella isolates were submitted to the Enteric Bacterial Infections Laboratory of Fundação Oswaldo Cruz (FIOCRUZ), Rio de Janeiro, Brazil, for serotyping.
Bacteriophage isolation, purification, and concentration
Each farm was considered a sampling unit. Farms were at least 30 km distant from each other, and had different water and feed sources, as well as different rearing environments. Salmonella Typhimurium (ATCC 14028) was used as host for the propagation of specific bacteriophages. The bacteriophage isolation, purification, and concentration were performed according to methodology described by Húngaro (2013). Three purification cycles were performed for each phage.
Bacteriophage specificity evaluation
Tests were carried out with Salmonella Abony (NCTC 6017), Salmonella Arizonae (ATCC 13314), Salmonella Choleraesuis (ATCC 10708), Salmonella Enteritidis (ATCC 13076), Salmonella Gallinarum (ATCC 9184), Salmonella Pullorum (ATCC 9120), Salmonella Typhi (ATCC 6539), and Salmonella Typhimurium (ATCC 14028). The lytic activity of the selected bacteriophages was evaluated according Clokie and Kropinski (2009), using a bacteriophage suspension at 1010 plaque-forming units (PFU)/mL.
Bacteriophage morphology analysis
Bacteriophage morphology was analyzed according to Atterbury et al. (2007). Eight microliters of the bacteriophage suspension was placed on the scanning microscopy grid surface lined with formvar resin (Koch Electron Microscopy, São Paulo, Brazil) and treated with uranyl acetate water solution (Sigma-Aldrich, St. Louis, MD) at 2% (w/vol). The grid was washed, dried at room temperature (approximately 24±2°C), and then observed under a transmission electron microscope (JEOL JEM 1220, Peabody, MA). Digital images were captured using an SIS Megaview III (Philips, Essen, Germany) camera.
In vitro bacteriophage viability at different pHs and bile salt concentrations
One hundred microliters of the bacteriophage suspension were added to 900 μL of saline magnesium (SM) buffer with pH adjusted to 2, 3, or 4 using HCl (1 mol L−1) (Vetec, São Paulo, Brazil). For the bile test under different bile salt concentrations, 100 μL of the bacteriophage suspension was added to solutions containing 900 μL SM buffer at three different bile salt (Difco) concentrations (0.1%, 0.5%, or 1%). In pH resistance test, samples were serially diluted in phosphate-buffered saline to neutralize the solution pH before plating. Both tests were conducted at 37±1°C and samples were removed at 30-min intervals for 2 h. Bacteriophages were counted using the brain-heart infusion (BHI) overlaying technique. The reduction of bacteriophage was calculated as the difference between the initial and final concentration at different pH levels and at different bile salt levels.
Evaluation of the in vitro activity of the bacteriophage pool
A pool of bacteriophages was prepared by mixing 1 mL of individual bacteriophage stocks containing 1010 PFU/mL each. Pooled suspensions at multiplicity of infection (MOI) values of 0.01, 1.0, 100, and 1000 were added to tubes containing Salmonella Typhimurium at 5.0 log10 CFU/mL in BHIb. A culture with no bacteriophage addition was used as control. Bacteriophage activity was determined by measuring absorbance in a spectrophotometer (SP22, Biospectro, Curitiba, Brazil) at 600 nm at 1-h time intervals after the bacteriophage suspensions were added until the optical density (OD) values remained constant. Pathogen counts were carried out at that point of each MOI using the surface-spotting technique, inoculating 20 μL in standard plate count agar (Difco). Plates were incubated at 37±1°C for 24±2 h, under aerobic conditions.
Experimental design and evaluation of the in vivo activity of the bacteriophage pool
All protocols involving animals were conducted under the approval of the Federal University of Viçosa Committee of Animal Health Care. Thirty pigs with similar body weight (90–100 kg) were housed in individual metabolic cages and randomly assigned to one of five treatments, with six replicates of one pig each. Each animal represented an experimental unit. Treatments included one control group (T1: no bacteriophage) and four different bacteriophage concentrations expressed in PFU/mL: 103 (T2); 105 (T3); 107 (T4); and 109 (T5).
Five milliliters of SM buffer containing 5.0 log10 CFU/mL Salmonella Typhimurium (ATCC 14028) was administered orally for 2 consecutive days. On the third day, fecal samples were collected to confirm the pathogen presence. Ten milliliters of a sodium bicarbonate solution (Na2HCO3, Sigma-Aldrich) (4% [pH 8.1]) was administered orally followed by 10 mL of SM buffer containing the bacteriophage pool, after which pigs were fasted for 18±2 h, simulating a common preslaughter practice. After the fasting period, pigs were sacrificed and 10 g of ileal digesta, 10 g of cecal content, and 10 g of the excreted fecal material were collected. Each sample was transferred to a flask containing 90 mL SM buffer. Salmonella was enumerated by the direct plating method using Rambach selective agar (Merck KGaA, Darmstadt, Germany) and incubated at 37±1°C for 24±2h.
Statistical analyses
In vitro results were evaluated by the test of Student-Newman-Keuls (SNK) at p<0.05. In vivo results were tested by the chi-square test (p<0.05). All data were analyzed using the SAS statistical package (SAS Institute, Cary, NC).
Results
Salmonella Typhimurium was detected in 48.2% of fecal samples (13/27). Six bacteriophages with desirable lysis characteristics (i.e., inhibition zone diameter between 1.0 and 3.0 mm) were isolated (Table 1). Neither the host pathogen nor bacteriophages with desired characteristics were isolated from farms A and B. On two other farms (G and H), the bacteriophage, but not the pathogen, was isolated. The selected bacteriophages presented lytic activity against more than one Salmonella serotype, as shown in Table 2. Bacteriophages analyzed by transmission electron microscopy presented 64-nm-diameter icosahedral head and small 16-nm-long tails, and were classified as belonging to the Caudovirales order and to the Podoviridae family (Fig. 1).

Transmission electron microphotograph of the isolated bacteriophages. Bacteriophages presented similar characteristics (i.e., short tail and icosahedral head, at 85,000 magnification).
According to serological identification; (+) Positive; (ND) Not detectable;
Final concentrations after enrichment.
PFU, plaque-forming unit.
−, Negative lysis result; +, positive lysis result.
The bacteriophages were more sensitive to pH 2 (p<0.05) as compared to pH 3 and 4 at 37±1°C after 120 min of contact. F3, F4, F6, and F7 bacteriophages showed higher pH sensitivity (p<0.05) compared to bacteriophages F5 and F8, and consequently, were less tolerant to pH 2. On the other hand, bacteriophage survival was not statistically different at pH 3 and 4 (Table 3).
Value followed by capital letters in the same column and small letters in the same row are statistically different at p<0.05 (Student-Newman-Keuls).
Table 4 shows that bacteriophage counts were not influenced by bile salt concentrations (p>0.05) after 120 min of contact at 37±1°C. However, when mean bacteriophage counts were compared among each other, bacteriophage F6 was the most sensitive and F5, the most resistant (p<0.05).
Figure 2 shows Salmonella Typhimurium culture absorbance reductions (p<0.05) after 6 h of challenge when different amounts of bacteriophage pool were added. The most significant reductions in pathogen counts (10.9 log10) were obtained when 107 and 109 PFU of bacteriophages were used, resulting in 1.6 and 2.5 log10 CFU/mL reductions, respectively, relative to the control group. Considering whether there is a low level of initial contamination of the animal, 1–2 decimal reductions can represent an increase in safety of the raw material.
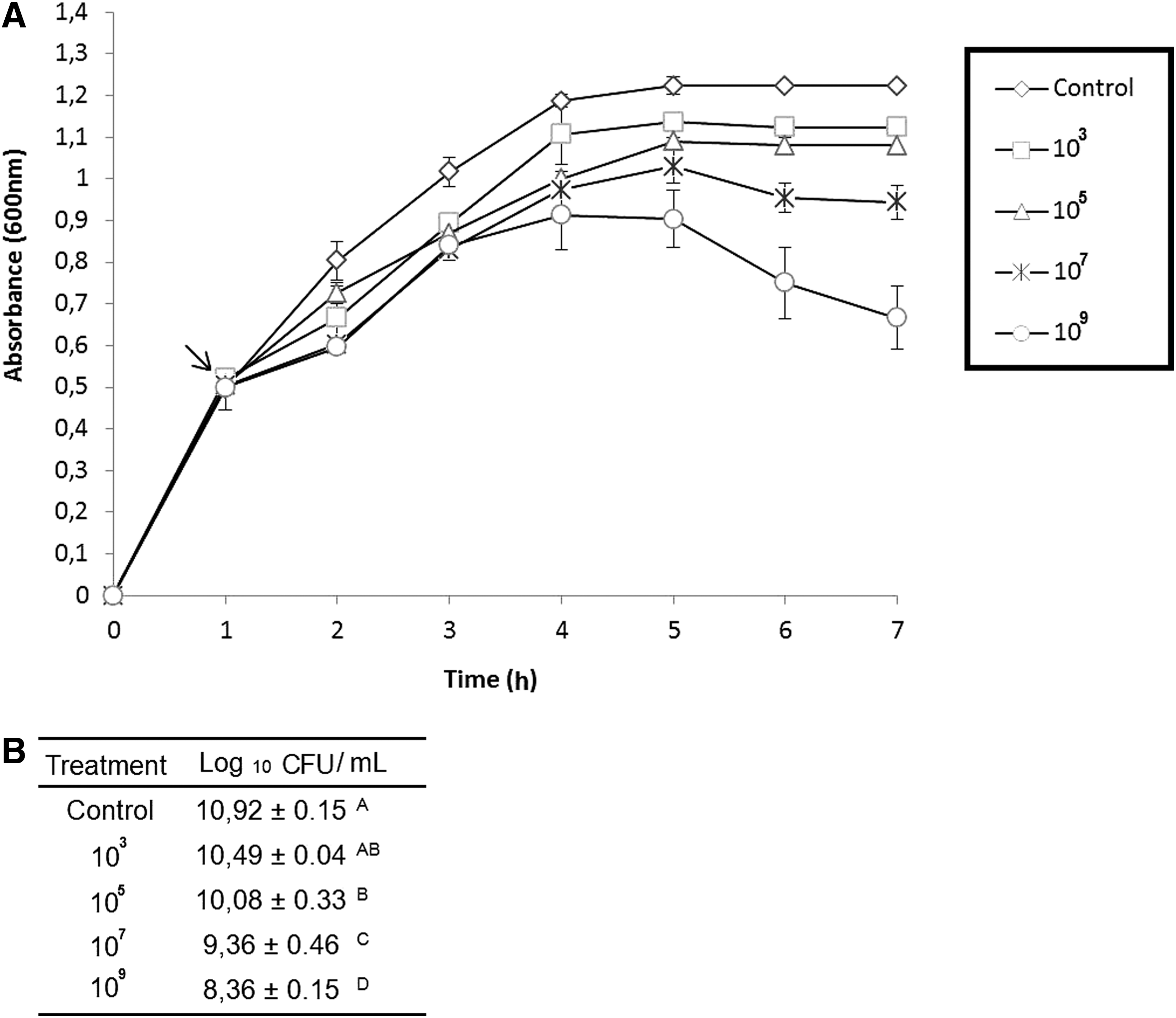
In vitro evaluation of the effect of the bacteriophage pool at different concentration against Salmonella Typhimurium.
When the efficiency of the bacteriophage pool was evaluated in vivo, there was a mean reduction of 1.0 log10 CFU/g both in the ileum and cecum with concentration of 107 and 109; however, these results were not statistically significant (p>0.05, Fig. 3). Before the beginning of the in vivo treatment with the different concentration of the bacteriophage pool, Salmonella was identified in 93.3% of the inoculated pigs (28/30), but only in 56.6% (17/30) after the treatment. Most of the nondetectable samples were obtained from pigs treated with concentration 107 and 109 (p>0.05, Fig. 3).
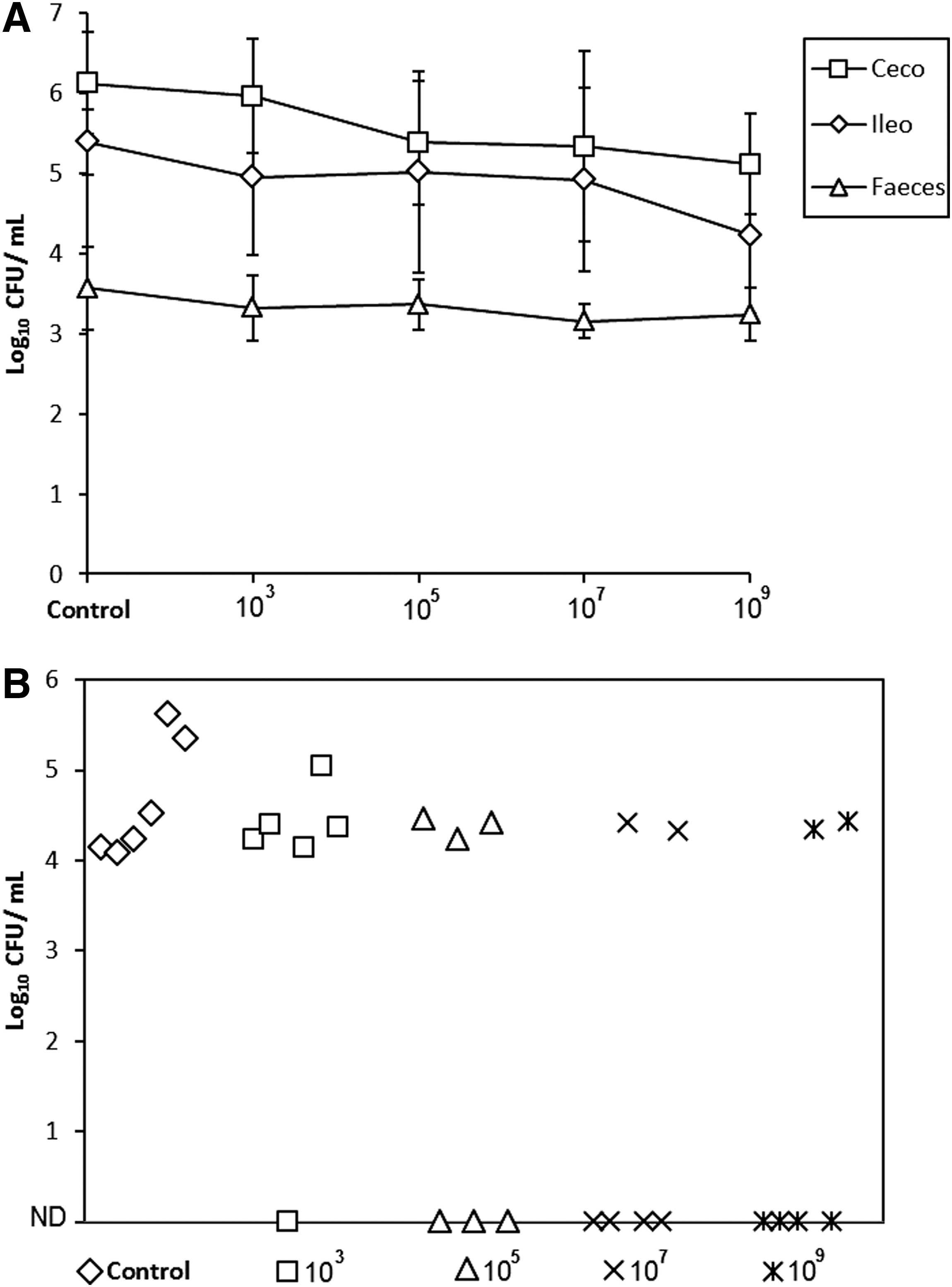
In vivo evaluation of the effect of the bacteriophage pool at different concentrations on Salmonella Typhimurium colonization.
Discussion
The frequent reports from around the world showing pigs carrying and shedding Salmonella Typhimurium in finishing herds and slaughter facilities indicates the need for multiple interventions to effectively reduce the contamination with Salmonella. However, the worldwide trend for reducing use of antimicrobials in animal production enforces the development of alternative strategies to reduce the incidence of pathogens in commercial farms (Davies, 2011; Kich et al., 2011; Wang et al., 2011; Stojanac and Stevancevic, 2013; Wales et al., 2013). The recent increased number of studies on the application of phages for the control of Salmonella and other pathogens in livestock and poultry shows the interest in and potential of this approach (Mahony et al., 2011).
Although Brazil is considered a major swine-producing country, there is a lack of data on the prevalence of Salmonella species in commercial swine farms, with only a few studies reported (Viott et al., 2013). In the present study, Salmonella Typhimurium was found on 50% of the commercial farms sampled. The incidence and prevalence of the serotype Typhimurium were consistent with the results reported by others in various countries throughout the world, as reported by Lee et al. (2011), Vico et al. (2011), Arguello et al. (2013), and Li et al. (2013). The occurrence of Salmonella Typhimurium in commercial farms can vary depending on the country, the nature of the production system, and the specific control measures in place. Moreover, differences in prevalence can be explained by additional factors, such as the low detection sensitivity of fecal sampling, the occurrence of intermittent shedding following exposure, and evolution and resolution of Salmonella epidemics without clinical manifestations (Rostagno et al., 2012).
Bacteriophage therapy has gained increasing credibility as an alternative to traditional antibiotic therapies and has been utilized as treatment for bacterial diseases caused by several foodborne pathogens (Hagens and Loessner, 2010). The most frequent serovar involved in Salmonella infections in food animals and/or contamination of pork production is Salmonella Typhimurium (Hooton et al., 2011), and therefore, we chose to focus on Salmonella Typhimurium. Here, Salmonella Typhimurium bacteriophages were isolated in 6/8 (75%) of the studied farms, values higher than those obtained by Callaway et al. (2010) and O'Flynn et al. (2006). The higher frequency of isolated bacteriophages that we obtained on commercial farms could be explained by a high frequency and/or concentration of Salmonella Typhimurium on the farms and its samples (Callaway et al., 2011).
The six isolated bacteriophages belong to the Podoviridae family. Podovirus are double-stranded DNA bacteriophages with genome size of 42 kbp that infected Salmonella serotypes belonging to groups B and D1 (Casjens and Thuman-Commike, 2011). According to Joerger (2003), Salmonella bacteriophages present binding specificity to some lipopolysaccharides (LPS), such as O12, found in the Enteritidis and Typhi group D1 serotypes, as well as Abony and Typhimurium group B serotypes. Some authors reported that bacteriophages against Salmonella Typhimurium isolated from poultry feces or sewage (but only effective against serogroups B and D1) belonged to the Myoviridae and Siphoviridae families (Heringa et al., 2010; Zhang et al., 2010). Usually, tailed phages use specific attachment fibers or spike proteins to establish first host cell surface contacts. Siphovirus uses its tail spike protein to recognize a long O chain LPS receptor (which starts the infection process) that is structural and functional homologue to podovirus surface attachment organelle, showing similar kinetics and enzymatic activities (Andres et al., 2012). It is clear that additional studies are still necessary to better characterize the different viruses in terms of genomics and their relationship with bacterial and animal species, life cycles, and survival characteristics to improve the phage therapy technology.
Bacteriophages can be found in all kinds of environments where bacteria grow such as soil, water, wastewater, even feces, indicating their ubiquity. Bacteriophages, which infect typically enteric bacteria such as Salmonella spp., should be resistant to the acid environment of the gastric juice, since it influences the stability, replication, and survival of the bacteriophage (Grabow, 2001). The effect of different pH conditions on the isolated bacteriophages sensitivity was consistent with the response obtained in studies on bacteriophages in other hosts (O'Flynn et al., 2006; Ma and Lu, 2008; Knezevic et al., 2011). It was shown in this study that the phages examined were sensitive to pH 2, but showed considerable stability at pH 3–4. This sensitivity is very important for the phages, based on their need for surviving within the gastrointestinal tract, where the pH can oscillate from acid to basic, forcing the bacteriophage to adapt in order to resist in a wide pH environment. These data demonstrate that pH sensitivity is one of the most important factors to formulate a phage cocktail that is going to be orally administered.
Salmonella bacteria spread easily in pigs by direct contact and environmental exposure on farms, during transit, and in lairage. Traditionally, some interventions, such as feed withdrawal before transport to the slaughterhouse, have been commonly applied to reduce the risk of Salmonella contamination, based on a reduced weight of the gastrointestinal tract resulting in a lower risk of lacerations during evisceration (Verbrugghe et al., 2011). However, the success of these interventions has been very limited, and Salmonella contaminations continue to be common along the pork-processing lines. Reducing the pathogen before it enters the slaughterhouse has been suggested to enhance the effectiveness of commonly applied interventions, leading to reduced human foodborne illnesses (Callaway et al., 2011). The implementation of bacteriophages as a preharvest strategy to kill Salmonella resident of the gastrointestinal tract has been used in several in vivo research studies. Lee and Harris (2001) reported a significant reduction of Salmonella in pig tonsils (4-log reduction) and cecum (2-log reduction) after bacteriophage administration. Wall et al. (2010) administered a pool of microencapsulated Salmonella bacteriophages, and observed reduced Salmonella counts in ileum (2-log reduction), tonsils (3-log reduction), and cecum (3-log reduction) in young pigs; but in the older pigs, reduction was less marked, both in ileum (1-log reduction) and cecum (1-log reduction). The observed difference between pig sizes/weights was explained by the intestinal microbiota characteristic of each pig-growing stage, suggesting that pig weight and development stage may influence the response to Salmonella infection, and consequently, the effectiveness of its control with bacteriophages. Saez et al. (2011) administered a pool of microencapsulated Salmonella bacteriophages mixed in the feed or by gavage and observed Salmonella reduction in the ileum and in cecum of market pigs. The feed group presented a 2-log reduction of ileum and cecum Salmonella counts, and the gavage group showed a reduction of 1-log just in the ileum. The counts in the cecum were not different from those in the control group; however, both administration routes did not completely eliminate the pathogen. Bacteriophages were recovered from the ileum and cecum in pigs that received bacteriophages in feed more frequently compared to the pigs receiving the phage by gavage. This contrast was probably due to the protection provided by feed particles to the low stomach pH. Although being encapsulated, bacteriophages were not able to resist passage through the gastric environment, where the pH at fasting conditions can be as low as 1.5–2.5 (DeRouchey et al., 2009).
The bacteriophage pool was capable of significantly reducing Salmonella counts in vitro (p<0.05), while the in vivo treatment results were not statistically significant (p>0.05) in any of the analyzed intestinal locations (ileum, cecum, feces), although Salmonella was detected in the feces of fewer animals after treatment with the bacteriophage pool at concentrations of 107 and 109 PFU/mL. These results differ from those obtained by Wall et al. (2010), who observed reduced Salmonella contamination in tonsils, ileum, and cecum by 2, 3, and 3 log10 CFU/mL, respectively, while it was not possible to detect Salmonella in 83.3% of the samples (5/6).
Conclusions
Based upon our data, it can be concluded that bacteriophages show promise as a potential viable strategy for the biocontrol of Salmonella Typhimurium in commercial swine farms. Also, our results show a high incidence of Salmonella Typhimurium contamination of Brazilian commercial swine farms, demonstrating a need for the development and implementation of prevention and control programs.
The sensitivity of bacteriophage exposed to pH 2 demonstrates the need for protective strategies for their oral administration, such as microencapsulation or other alternatives. Additional research is needed on the characterization and efficacy of bacteriophages, as well as on their relationship with bacterial and animal species, contributing to the improvement of phage therapy technology.
Footnotes
Acknowledgments
The research study was funded by Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq).
Disclosure Statement
No competing financial interests exist.