
Abstract
Contamination of retail meat with extended-spectrum β-lactamase (ESBL)–producing Escherichia coli has been reported, but only limited data have been documented in Japan. One hundred fifty-three retail foods including chicken meat, beef, pork, and vegetables were purchased from 29 supermarkets between January and October in 2010. ESBL producers were recovered from each food sample using McConkey agar plate supplemented with 1 mg/L of cefotaxime. ESBL type was identified by DNA sequencing analysis after polymerase chain reaction amplification. Antibiogram, O serotype, plasmid replicon type, pulsotype, and multilocus sequence type were also determined. Fifty-two epidemiologically unrelated Escherichia coli isolates producing ESBL were recovered from 35 (22.9%) of 153 samples, all of which were chicken meat. ESBL types were mainly CTX-M-2 group followed by CTX-M-1 group and CTX-M-8 group. The numbers of bacterial isolates (8 of 21, 38.1%) harboring bla CTX-M-8 recovered from imported meat samples were significantly larger than those of domestic ones (one of 31, 3.2%) (p<0.05). Nine O serotypes (mainly O8, O25, and O1) were found, together with O-antigen untypable (OUT). Four E. coli belonging to the O25b:H4-ST131 clone were recovered from domestic (n=1) and imported meat samples (n=3), respectively. These four isolates were susceptible to fluoroquinolones, although the E. coli O25b:H4-ST131 clone producing CTX-M-15, which is predominant in human isolates, is usually resistant to fluoroquinolones. By contrast, five CTX-M-15-producing E. coli strains were recovered only from domestic meat samples, and their serotypes were O8 or OUT instead of predominant serotype O25b. Our results showed that ESBL-producing E. coli isolates recovered from retail chicken meat samples in Japan are generally divergent in both genetic and serological aspects. Further comparative analyses of bla CTX-M-mediating genetic elements would be continued in the next step to characterize the ESBL producers from retail foods in Japan.
Introduction
P
In many food animals such as chickens, pigs, and cattle, certain antimicrobials such as colistin sulfate, avilamycin, and monensin were approved to add to animal feeds for preventing infections in many countries. By contrast, the veterinary use of fluoroquinolones such as enrofloxacin and cephalosporins such as ceftiofur and cefquinome have been approved only for therapeutic purpose for livestock infected with pathogenic bacteria. Indeed, the veterinary use of these antimicrobials has been strictly restricted, but the use of such antimicrobials may well contribute to the emergence and spread of ESBL producers in livestock farming environments through selection of bacterial strains harboring genetic determinants responsible for cephalosporin resistance such as bla CTX-M and bla CMY. Actually, Ma et al. have reported the occurrence of ESBL-producing E. coli in fecal samples of healthy ducks and environmental samples from a duck farm in South China (Ma et al., 2012), and an increasing amount of data regarding this issue has been accumulated to date (Dhanji et al., 2010; Ewers et al., 2010; Ho et al., 2011). Food animals and their products are now suspected to be one of the potential sources in the dissemination of the antimicrobial-resistant bacteria, especially ESBL-producing E. coli (Overdevest et al., 2011). In Japan, Hiroi et al. and Asai et al. have reported the prevalence of ESBL-producing bacteria in food-producing animals (Asai et al., 2011; Hiroi et al., 2012), however, there have been few reports about the dissemination of ESBL-producing E. coli in samples derived from food animals and/or their products collected since 2007. The aim of our study was to assess the recent state of contamination with ESBL-producing E. coli in retail food samples and to characterize the types of ESBLs, together with the serotype and genotype of ESBL-producing E. coli isolates in Japan. Moreover, we also compared the prevalence of ESBL producers in domestic and imported meat samples, since imported meat products have recently become a commonplace in the Japanese market.
Materials and Methods
Sample collection and bacterial isolation
One hundred fifty-three retail food samples (chicken, beef, pork, and vegetables) were purchased from 29 separate supermarkets in Aichi Prefecture, Japan, between January and October 2010. As for raw chicken meat, 42 samples were domestic products and 26 samples were ones imported mainly from South America, and the remaining samples were mincemeat mixed of domestic and imported meat. Approximately 10 g of each sample was incubated in brilliant green lactose bile broth (SYSMEX bioMérieux Co., Ltd., Tokyo, Japan) at 35°C for 24 h; aliquots (100 μL) of the incubated bacterial preculture were inoculated onto McConkey agar plates (Eiken Chemical Co., Ltd., Tokyo, Japan) supplemented with 1 mg/L of cefotaxime (Jouini et al., 2007), and the plates were incubated at 35°C for 24 h. The two to four colonies growing on the CTX-MacConkey agar plate were picked up and identified in terms of bacterial species by using the API-20E system (SYSMEX bioMérieux). These isolates were further subjected to screening of ESBL producers and pulsed-field gel electrophoresis (PFGE) analysis.
Screening and genetic identification of ESBL
ESBL screening was performed on the basis of the double-disk synergy test by using three different commercially available discs: ceftazidime, cefotaxime, and amoxicillin/clavulanic acid, according to the protocol recommended by the Clinical and Laboratory Standards Institute (CLSI, 2009b) and confirmed using Etest (SYSMEX bioMérieux).
The presence of CTX-M-type β-lactamase genes was determined by polymerase chain reaction (PCR) using primers specific to the CTX-M-1 group, CTX-M-2 group, CTX-M-8 group, or CTX-M-9 group as described elsewhere (Shibata et al., 2006; Dallenne et al., 2010). The TEM- and SHV-type β-lactamase genes were detected by PCR, and their genotypes were further determined by sequencing analysis (Yagi et al., 2000). The nucleotide sequences were analyzed with BLAST software (
Cluster analysis by PFGE
PFGE typing of the ESBL-producing isolates was performed as described elsewhere (Barrett et al., 1994). Plug containing whole genomic DNA was digested with XbaI (Takara Bio. Inc., Tokyo, Japan), and electrophoresis was performed using a CHEF-DR III system (Bio-Rad Laboratories, Hercules, CA) with pulses ranging from 2.2 to 54.2 s at a voltage of 6 V/cm at 14°C for 19 h. A dendrogram showing genetic relatedness among the isolates was constructed with Fingerprinting II software (Bio-Rad Laboratories). In the isolates from each food sample, when ≥85% genetic similarity was observed, these were classified as a clone with a common genetic background, and one representative isolate was further studied. When <85% genetic similarity was observed, these were considered as a different clone and separately treated in this study (Carrico et al., 2005).
Serotyping of ESBL-producing E. coli
Serotyping of each E. coli isolate was performed with the E. coli antisera “SEIKEN” Set 1 (Denka Seiken, Tokyo, Japan) for O-antigen and Set 2 (Denka Seiken) for H-antigen according to the manufacturer's instructions. Serotypes that could not be distinguished by this method were designated OUT (O-antigen untypable) or HUT (H-antigen untypable). Genetic serotyping of each bacterial isolate was performed according to the procedures described elsewhere (Clermont et al., 2008).
Antimicrobial susceptibility testing
The antimicrobial susceptibility of each isolate was determined by the agar dilution method, according to the protocol recommended by the CLSI in document M07-A9 (CLSI, 2009a). The antimicrobial agents were obtained from the following sources: piperacillin, cefotaxime, ceftazidime, imipenem, aztreonam, gentamicin, minocycline, fosfomycin, Wako Pure Chemical Co., Inc., Tokyo, Japan; cefmetazole, amikacin, chloramphenicol, Sigma-Aldrich Japan, Tokyo, Japan; ciprofloxacin, levofloxacin, Daiichi-Sankyo Company, Tokyo, Japan; and flomoxef, Shionogi & Co., Ltd., Tokyo, Japan. Susceptibilities of ESBL-producing E. coli to each antimicrobial agent were categorized into susceptible, intermediate, or resistant according to the CLSI criteria. Type Culture Collection (ATCC) 25922 (Microbiologics, Inc., MN) was used as reference strain.
Multilocus sequence typing
Multilocus sequence typing (MLST) of ESBL-producing E. coli isolates was performed with seven conserved housekeeping genes (adfK, fumC, gyrB, icd, mdh, purA, and recA). The MLST procedure, including allelic type and sequence type (ST) assignment methods, was done according to the website (
Conjugation studies and plasmid replicon typing
The conjugation experiments were performed with rifampin-resistant E. coli CSH-2 (metB F-, nalidixic acid resistant, rifampicin resistant) as the recipient strain by the filter mating methods. Transconjugants were selected on LB agar plates supplemented with rifampin (100 mg/L) and cefotaxime (2 mg/L) (Wako Pure Chemical Co., Inc., Tokyo). For resultant transconjugants and the parent isolates, plasmid replicon typing was performed as described elsewhere (Carattoli et al., 2005).
Statistical analysis
Comparisons of proportions of ESBL producers recovered from both imported meat samples and domestic products and comparisons of proportions of susceptible isolates between imported and domestic products were made by continuity-adjusted x 2 test with SPSS software (version 20.0 for Windows; SPSS Inc., Chicago, IL); p<0.05 was considered as denoting a significant difference.
Results
Isolation of ESBL producers from food samples
Thirty-five (22.9%) of 153 food samples were contaminated with ESBL-producing bacteria. The ESBL producers were only isolated from chicken meat samples, and the isolation rate (15 of 26, 57.7%) of imported chicken meat samples was slightly higher than that (19 of 42, 45.2%) of domestic chicken meat samples. Of 85 isolates recovered from 35 chicken meat samples, clonally related isolates recovered from the same food samples were excluded based on the cluster analysis by PFGE. The range of clonally different isolates was one to three per chicken sample. As a result, 53 clonally unrelated isolates (domestic products, 32; imported products including one mix product, 21) were recovered from 35 chicken meat samples as probable ESBL producers. Of the 53 isolates, 52 were identified as E. coli and the remaining one was Klebsiella pneumoniae. Fifty-two E. coli isolates were subjected to further studies.
Genotyping of ESBL-producing E. coli
Forty-three E. coli isolates produced CTX-M-type ESBLs, while the remaining nine isolates produced TEM-derived or SHV-derived ESBLs (Table 1). Of the bla CTX-M genes, 22 (51.2%), nine (20.9%), five (11.6%), five (11.6%), and two (4.7%) coded for CTX-M-2, CTX-M-8, CTX-M-1, CTX-M-15, and CTX-M-3 ESBL, respectively. The number of bacterial isolates (eight of 21, 38.1%) harboring bla CTX-M-8 recovered from imported meat samples was significantly larger than that of domestic ones (one of 31, 3.2%) (p<0.05), but the number of the isolates harboring bla CTX-M-2 in imported ones (11 of 21, 52.4%) was slightly higher than that of domestic products (11 of 31, 35.5%). Of the 52 ESBL-producing E. coli isolates, seven and one were SHV-12 and SHV-2 producers, respectively, and seven of eight SHV-producers were from domestic meat samples (Table 1).
One sample was an imported product mixed with domestic product, and classified into the imported products.
One isolate harbored both bla CTX-M-8 and bla TEM-135.
Serotype of 52 ESBL-producing E. coli isolates
PFGE profiles of 52 ESBL-producing E. coli isolates were dissimilar to each other (Fig. 1). However, several strains carrying the same ESBL-genes were found to be genetically different (e.g., S7, S416), while closely related strains (e.g., S150, S424) harbored different ESBL genes. Thirty-one (59.6%) were O untypeable, and the serogroups of the remaining isolates were O8 (seven of 52, 13.5%), followed by O25 (five of 52, 9.6%) and O1 (two of 52, 3.8%), respectively. Of the five E. coli O25 isolates, four and one isolates were identified as O25b and O25a types, respectively, by PCR. In addition, all four O25b isolates, which harbor bla CTX-M-2 (n=2), bla CTX-M-8 (n=1), and bla SHV-12 (n=1), belonged to ST131 by MLST analysis.
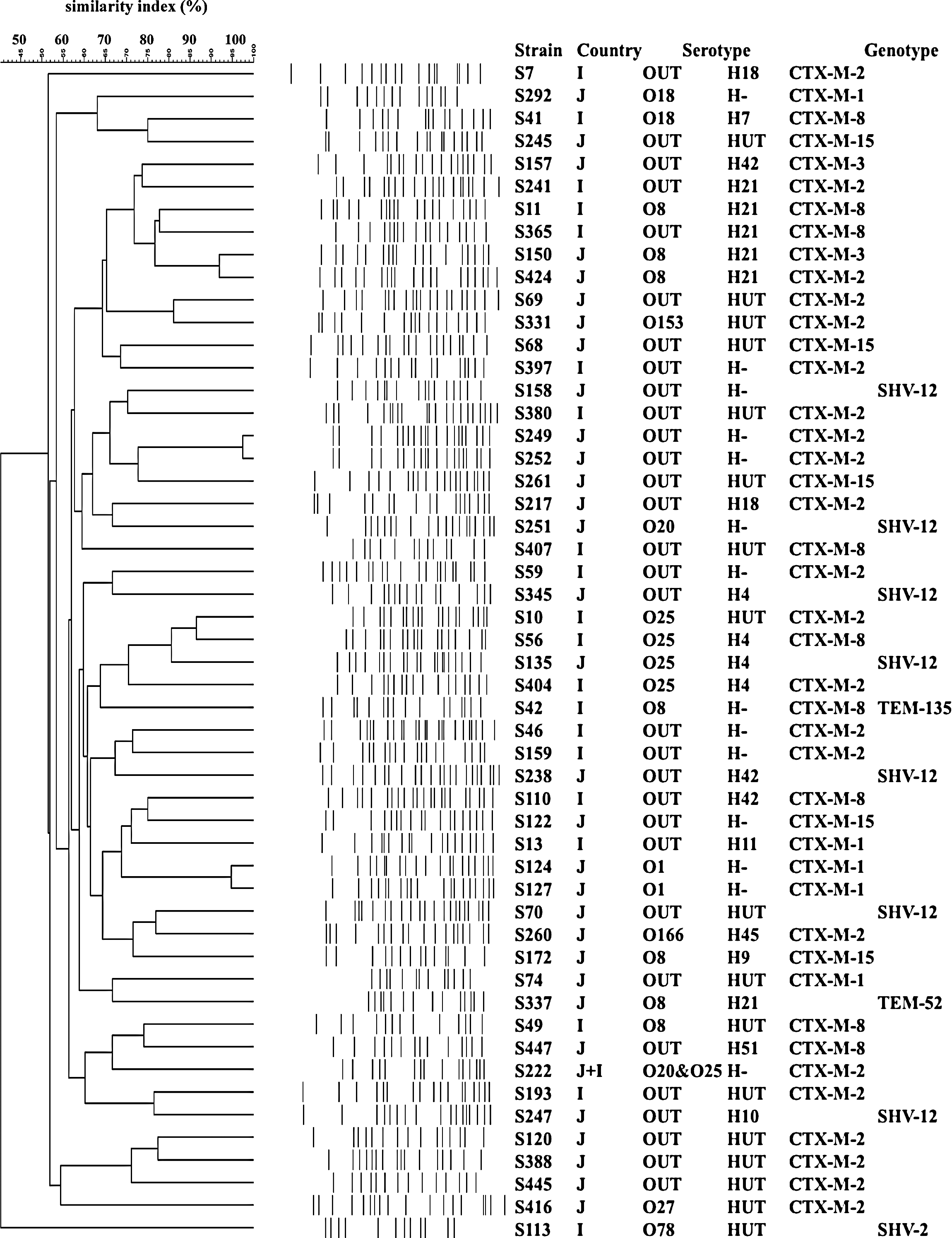
Dendrogram of pulsed-field gel electrophoresis (PFGE) pattern among bla CTX-M-harboring Escherichia coli (based on unweighted pair group method with arithmetic averages cluster analysis). Ladder patterns were analyzed by Fingerprinting II software (Bio-Rad Laboratories) to calculate the Dice similarity index. Isolates were considered to belong to a group of clonally related strains if the dice similarity index of the PFGE pattern was ≥85% (Carrico et al., 2005). When two isolates were recovered from separate parts such as leg and chest purchased from the same supermarket at intervals of more than 3 weeks, the isolates were subjected to further studies as clonally unrelated isolates in the present study, even if they demonstrated 100% similarity by PFGE. J, domestic chicken meat samples; I, imported chicken meat samples; OUT, O-antigen untypeable; HUT, H-antigen untypeable.
Antimicrobial susceptibility profiles of ESBL-producing E. coli
As shown in Table 2, ESBL-producing E. coli isolates were susceptible to cefmetazole, flomoxef, imipenem, amikacin, and fosfomycin. On the other hand, these isolates tended to be resistant to ceftazidime, aztreonam, and minocycline as well as cefotaxime. Among antimicrobial agents including β-lactams tested, susceptibility profiles to chloramphenicol and fluoroquinolones of isolates from domestic products were similar to those from imported products, although the resistance rate to gentamicin in bacterial isolates from imported products was significantly higher than those from domestic ones (p<0.05) (Table 2). In addition, they showed resistance to β-lactams such as piperacillin, cefotaxime, and/or ceftazidime, whereas they were not resistant to fluoroquinolones such as ciprofloxacin and levofloxacin.
Susceptibilities of 52 ESBL-producing E. coli to each antimicrobial agent were categorized into susceptible, intermediate, and resistant in accordance with Clinical and Laboratory Standards Institute criteria.
Proportions of R+I were compared between imported meat samples and domestic meat samples.
P-value was not obtained.
S, susceptible; I, intermediate; R, resistant; PIPC, piperacillin; CMZ, cefmetazole; CTX, cefotaxime; CAZ, ceftazidime; FMOX, flomoxef; IPM, imipenem; AZT, aztreonam; GM, gentamicin; AMK, amikacin; MINO, minocycline; CP, chloramphenicol; FOM, fosfomycin; CPFX, ciprofloxacin; LVFX, levofloxacin.
Plasmid replicon types
CTX-M-15-producing E. coli belonging to the O25b:H4-ST131 is a global epidemic clone, and their isolation rate has been increased recently in Japan. Therefore, conjugation experiments were performed for five isolates (non-O25) harboring bla CTX-M-15 and four O25b:H4-ST131 isolates. The conjugal transfer of the bla CTX-M-carrying plasmid to recipient cells was successful in four non-O25 and two O25b isolates. Replicon types of the plasmid were mainly I1-γ (4 of 6) followed by IncF, and IncP (Table 3).
OUT, O-antigen untypeable; ND, conjugal transfer of bla CTX-M to recipient cells did not succeed.
Discussion
Contamination of foods with ESBL-producing E. coli has recently become a worldwide concern. ESBL-producing E. coli isolates were recovered from 62 (29.5%) among 210 batches in raw chicken meat imported by the United Kingdom from South America (Dhanji et al., 2010). In Tunisia, the distribution of isolation rates of ESBL-producing E. coli were between 12.6% and 26% among the food samples including chicken and turkey (Jouini et al., 2007). In the present study, the isolation rate of ESBL producers was 50.7% (35 of 69) in chicken meat samples. The high prevalence of ESBL producers in chicken meat in the present study was consistent with Japanese previous reports and was higher than reports from the United Kingdom and Tunisia (Kojima et al., 2005; Jouini et al., 2007; Dhanji et al., 2010; Asai et al., 2011; Hiroi et al., 2012). In Japan, the oxyimino-cephalosporins such as ceftiofur and cefquinome are approved to treat bacterial diseases in cattle and pigs, but are not allowed for use in broilers and chickens. At present, the reason for the high prevalence of ESBL producers in chicken meat samples remains unclear.
Some investigators have reported that CTX-M-type ESBLs detected from chicken meat consisted of CTX-M-1, −2, −14, and −15 (Belgium), CTX-M-2 and −8 (United Kingdom), and CTX-M-1, −8, and −14 (Tunisia), respectively, but the CTX-M-types were somewhat different from those isolated from patients admitted to clinical settings (Jouini et al., 2007; Smet et al., 2008; Dhanji et al., 2010). By contrast, in the Netherlands, a probable relationship between the contamination of chicken meat with drug-resistant bacteria and the appearance of ESBL-producers in humans has been reported (Leverstein-van Hall et al., 2011; Overdevest et al., 2011). In the present study, 43 bla CTX-M-harboring E. coli isolates were recovered from 35 chicken meat samples, and their CTX-M-types were mainly CTX-M-2 in both domestic and imported meat samples, followed by CTX-M-8 in imported meat samples. These findings are consistent with those reported from the United Kingdom, suggesting that the genotypes of ESBLs detected from chicken meat samples were very similar to those from chicken meat in the United Kingdom, which would reflect the fact that both countries import chicken meat from South America. In Japan, the predominant CTX-M-types were CTX-M-14 and CTX-M-9 belonging to the CTX-M-9-group enzymes in clinical isolates (Suzuki et al., 2009). Thus, the CTX-M-types of ESBL-producing E. coli recovered from chicken meat samples in the present study were inconsistent with those of clinical isolates detected more recently in Japanese hospital settings. The most probable reason for the increasing isolation of CTX-M-9-group ESBL producers from humans in Japan would be the acquisition of E. coli that produce CTX-M-9-group enzymes from different routes, such as human-to-human or pets-to-human transmissions (Harada et al., 2012).
In the present study, several genes belonging to the bla CTX-M-1-group, such as bla CTX-M-3 and bla CTX-M-15, were found from retail chicken meat samples, but the O serotypes of E. coli harboring the bla CTX-M-1-group gene were somewhat different from those of ESBL-producing E. coli isolated in Japanese clinical settings (Matsumura et al., 2012). PFGE analysis showed that strains carrying the same ESBL genes were genetically different, while closely related strains harbored different ESBL genes, suggesting that clonal spread is less important for the distribution of antimicrobial-resistant E. coli. Recently, Zheng et al. have reported that ISEcp1-like elements were found in the upstream of the bla CTX-M-9-group genes of ESBL producers obtained from food animals, and such genetic structures were very similar to those of humans (Zheng et al., 2012). Further comparative analyses of environmental genetic elements mediating bla CTX-M would be necessary to assess the probability of transmission of ESBL producers via retail foods in Japan.
The pandemic of CTX-M-15-producing E. coli in the past decade has been reported, especially in Europe (Woodford et al., 2004; Rogers et al., 2011). The epidemic E. coli O25b:H4-ST131 clone producing CTX-M-15 was consistently resistant to fluoroquinolones as well as various β-lactams (Nordmann and Poirel, 2005). In the present study, no CTX-M-15-producing O25b:H4 isolate was found, but four E. coli O25b:H4-ST131 isolates that produced CTX-M-2 or CTX-M-8 and five CTX-M-15 producers, serotypes of which were O8 or O-untypable, were recovered from retail chicken meat samples. Interestingly, these isolates were unexpectedly susceptible to fluoroquinolones. Although Inc types of plasmids encoding genes for CTX-M-15 in E. coli clinical isolates were usually IncFII, FIA, or FIB (Carattoli, 2009), the replicon types of three CTX-M-15-producing E. coli isolates were IncI1-γ plasmids in our study. Although no direct comparison between chicken meat and human isolates was performed in the present study, these results indicated that the CTX-M-15-producing E. coli isolates recovered from chicken meat samples have different genetic backgrounds.
Our study presents several limitations. The number of samples processed was small, and systematic surveillance was not used. However, we purchased food samples from 29 separate supermarkets between January and October 2010, and ESBL producers were excluded when the isolates were suggested to belong to the same clone by the cluster analysis of PFGE profile. This study seems to indicate the recent state of contamination with ESBL-producing microbes in retail foods. Indeed, it should be considered that plasmid and mobile genetic elements conferring resistance to third-generation cephalosporins might spread within E. coli via gene transfer, but the cluster analysis of PFGE profile could not exclude the possibility described above. It seems very important to investigate horizontal gene transfer such as exchanges of plasmids or mobile genetic elements carrying genes for ESBLs between bacteria isolated from foods. In the next step of our investigation, we will carefully check the genetic structures of the mobile elements carrying ESBL genes recovered from retail chicken meat and compare them to the relevant mobile elements from human isolates accumulated in the database.
Conclusions
We found a higher-than-expected prevalence of ESBL-producing E. coli in retail chicken meat samples in Japan. No clonal spread of ESBL-producing isolates contaminating food samples was observed in the present study, but the high prevalence rate of ESBL producers recovered from food samples might well depend on frequent horizontal gene transfer between bacterial isolates, through exchanges of plasmids and/or mobile elements carrying ESBL genes. Further surveillance and molecular epidemiological investigations conducted in an interdisciplinary way so as to consider not only the human but also the veterinary fields would be essential to predict the future spread of ESBL producers in both humans and animals.
Footnotes
Acknowledgments
We thank Dr. Takaaki Kondo for his helpful suggestion about statistical analyses. This study was supported by the Ministry of Health, Labour, and Welfare of Japan (Grant Nos. H21-Shinkou-Ippan-008 and H24-Shinkou-Ippan-010).
Disclosure Statement
No competing financial interests exist.