
Abstract
Our findings show that temperature plays a significant role in the induction of acid-stress adaptation in Listeria monocytogenes, and two distinct patterns were observed: (1) Presence of sublethal acid at 37°C or 22°C significantly induced acid-stress adaptation; and (2) Presence of sublethal acid at 4°C did not induce any acid-stress adaptation. Both patterns were confirmed by two experimental models: (1) L. monocytogenes cells were first grown at 37°C and then exposed to sublethal acid at 37°C, 22°C, and 4°C prior to lethal acid challenge; (2) Alternatively, L. monocytogenes cells were first grown at 4°C for 20 days before pre-exposure to sublethal acid and then challenged with lethal acid. Regardless of whether L. monocytogenes cells were simultaneously exposed with both cold stress and sublethal acid stress, or subjected to cold growth first before exposure to sublethal acid, no acid-stress adaptation was induced at 4°C. We also found that acid-stress adaptation in L. monocytogenes did not occur in acidic whey at 4°C. Bead beating treatment prior to mild acid pre-exposure at 4°C partially induced acid adaptation in L. monocytogenes. Our findings suggest that cold temperature can prevent the risk of acid-stress adaptation in L. monocytogenes.
Introduction
I
L. monocytogenes exhibits sophisticated adaptive mechanisms to counteract higher levels of lethal acid, heat, salt, or oxidative stresses after pre-exposure to sublethal concentrations of the same stress or different stress environments (O'Driscoll et al., 1996; Lou and Yousef, 1997; Bolton and Frank, 1999). Of all the stress adaptations, acid-stress adaptation in L. monocytogenes has received extensive attention. Acid-stress adaptation in L. monocytogenes, where cells exhibit enhanced acid tolerance at a lethal pH of 3.5 after pre-exposure to mild acid stress (pH 5.0), was first reported by Hill (1995). Further studies revealed that the activation of acid-stress adaptation required protein de novo synthesis (Davis et al., 1996; O'Driscoll et al., 1996). Two enzymatic systems, glutamate decarboxylase (GAD) system and arginine deiminase (ADI) systems, are critical in conferring acid-stress adaptation to L. monocytogenes (Cotter and Hill, 2003; Ryan et al., 2009).
Several factors play critical roles in controlling the induction of acid-stress adaptation in L. monocytogenes. Extensive studies have determined the influence of sublethal acid concentration, exposure time, type of acidulant, and bacterial growth stage on acid-stress adaptation in L. monocytogenes. Davis et al. (1996) determined that pre-exposure at pH 5.0 for 1 h was optimal to confer L. monocytogenes with acid-stress adaptation. With respect to growth stage, it is well known that stationary phase cells of L. monocytogenes have better acid tolerance compared to log phase cells (O'Driscoll et al., 1996; Datta and Benjamin, 1997). All these studies were conducted at the optimum growth temperatures. Under food processing conditions, L. monocytogenes cells may be exposed to suboptimal temperatures (4°C–22°C), which are much lower than that of laboratory conditions. The effect of refrigeration temperatures on acid-stress adaptation in L. monocytogenes is not known. Therefore, the objective of our study is to determine the effect of temperature on acid-stress adaptation in L. monocytogenes serotypes 4b and 1/2a.
Materials and Methods
Bacterial strains
The bacterial strains used in this study included L. monocytogenes Scott A (serotype 4b, human clinical, FDA) and L. monocytogenes EGD strain (BUG 600) (serotype 1/2a, clinical, Institute Pasteur, France). Working stock cultures were prepared by streaking frozen stock culture (stored at −80°C) onto tryptic soy agar (TSA) slants and maintained at 4°C. Overnight cultures were prepared by inoculating one loop of working stock cultures in 10 mL tryptic soy broth supplemented with 0.6% yeast extract (TSB-YE) and incubating at 37°C for 18–20 h. Early log-phase culture was prepared by transferring 200 μL overnight culture into 20 mL TSB-YE and incubating at 37°C until OD600nm reached ∼0.15 (Thermon Electron Corporation). Experiments were performed using 37°C grown early log-phase cells unless otherwise noted.
Effect of temperature on induction of acid-stress adaptation
The temperature-dependent acid adaptation was evaluated by initially pre-exposing the L. monocytogenes cells to pH 5.0 (1 M lactic acid) or 7.2 TSB-YE at three different temperatures (37°C, 22°C, and 4°C) for 1 h and subsequently performing postlethal acid challenge in pH 3.5 TSB-YE (3M lactic acid) at 37°C.
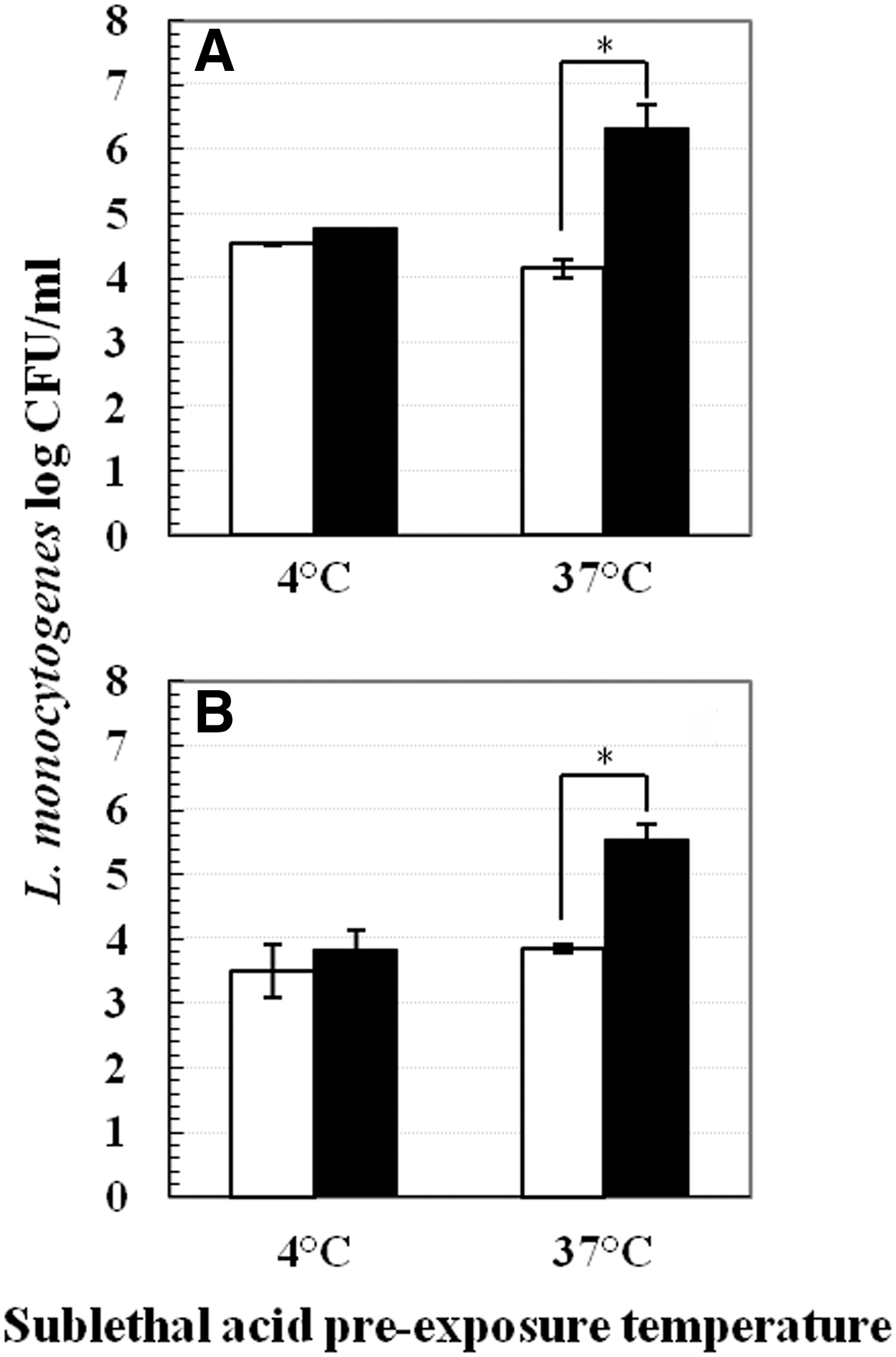
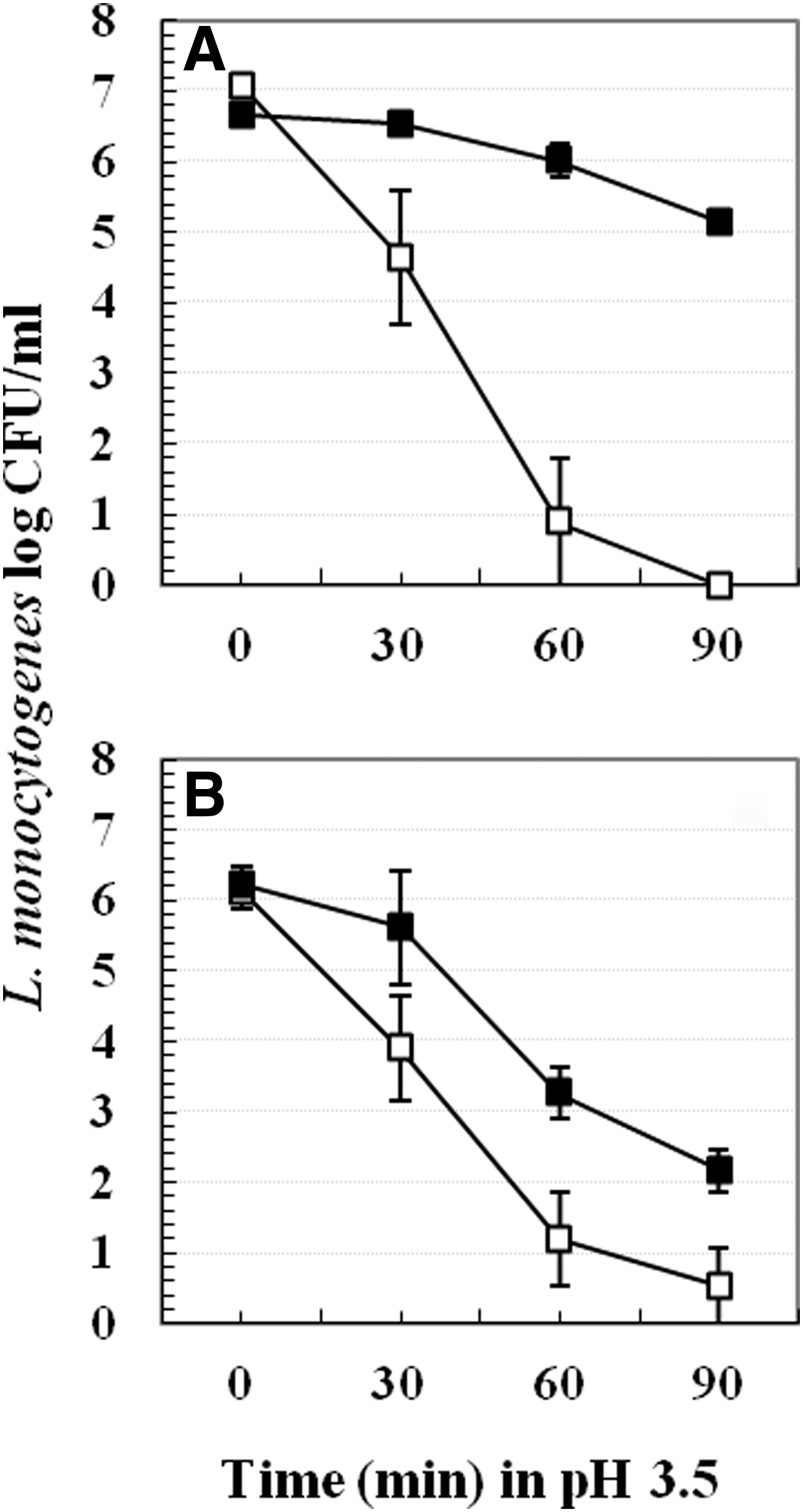
One milliliter of 37°C grown early log phase cultures were transferred into 9 mL pre-acidified (mild acid-stressed cells) or neutral TSB-YE (nonstressed cells) for pre-exposure at 37°C, 22°C, or 4°C. Tubes containing 9 mL pre-acidified or neutral broth were previously equilibrated to 37°C, 22°C, or 4°C. After 1 h pre-exposure, mild acid-stressed and non-stressed samples were centrifuged (Sorvall Biofuge Fresco, 16,060×g). While centrifuging, samples pre-exposed at 4°C were centrifuged at 4°C whereas samples pre-exposed at 37°C or 22°C were centrifuged at room temperature. After centrifugation, cell pellets were resuspended in pH 3.5 TSB-YE at 37°C for up to 180 min. L. monocytogenes survivors were spread plated at different time intervals on TSA-EF (TSA containing 1.0% esculin and 0.05% ferric ammonium citrate) and incubated at 37°C for 48 h (Fig. 1).

Survival of Listeria monocytogenes Scott A (serotype 4b)
Similar experiments were also performed using early log-phase cold (4°C) grown cells (OD600nm ∼0.15), but only two pre-exposure temperatures (37°C and 4°C) were used and survivors following 22°C postlethal acid treatments were enumerated at 90 min for Scott A and at 180 min for BUG 600 (Fig. 2).
Survival of Listeria monocytogenes Scott A
Determining factors affecting lack of acid adaptation at 4°C
Increase of mild acid pre-exposure time, various mild acid concentrations, change of acidulants, and addition of salt were analyzed to determine the acid adaptation of 4°C mild acid-stressed cells. As a control, 37°C mild acid-stressed and nonstressed cells were used in all experiments. All experiments were performed using L. monocytogenes Scott A grown to early log phase at 37°C. Postlethal acid inactivation treatment was performed in pH 3.5 TSB-YE (3M lactic acid) at 22°C for 60 or 90 min and survivors were enumerated on TSA-EF. Throughout all experiments, cells pre-exposed at 4°C were centrifuged at 4°C while 37°C pre-exposed cells were centrifuged at room temperature. Lactic acid (1M) was used for mild-acid pre-exposure unless otherwise noted.
The following series of experiments were conducted for the proof of concept at 4°C: 1. Increasing mild-acid pre-exposure time. Pre-exposure was conducted in pH 5.0 or 7.2 TSB-YE at 37°C or 4°C. Pre-exposure time periods tested at 37°C were 15, 30, 45, and 60 min, whereas time periods tested at 4°C were 1, 4, 8, and 24 h. At each time point, mild acid-stressed and nonstressed cells were centrifuged and subjected to lethal inactivation (Fig. 3). 2. Varying mild-acid concentrations. Cells were pre-exposed to different pHs (4.0, 4.5, 5.0, 5.5, 6.0, and 7.2) for 1 h at 37°C or 4°C. After pre-exposure, samples were centrifuged and cell pellets were subjected to lethal acid inactivation (Fig. 4). 3. Effect of various pre-exposure acidulants. Instead of lactic acid, 1M of hydrochloric acid and acetic acid were used to acidify TSB-YE to pH 5.0. Pre-exposure was performed in pH 5.0 or 7.2 TSB-YE for 1 h at 37°C or 4°C. Thereafter, cell pellets were obtained and subjected to lethal inactivation (Fig. 5). 4. Addition of sodium chloride. TSB-YE used in this study contained 0.5% sodium chloride. Additional salt concentrations tested include 2%, 4%, 6%, 8%, and 10%. Mild acid-stressed and nonstressed cells were prepared by pre-exposure to pH 5.0 or 7.2 TSB-YE supplemented with various sodium chloride concentrations (0.5%, 2%, 4%, 6%, 8%, and 10%) at 37°C or 4°C for 1 h. After pre-exposure, cells were pelleted and subjected to lethal acid inactivation (Fig. 6).
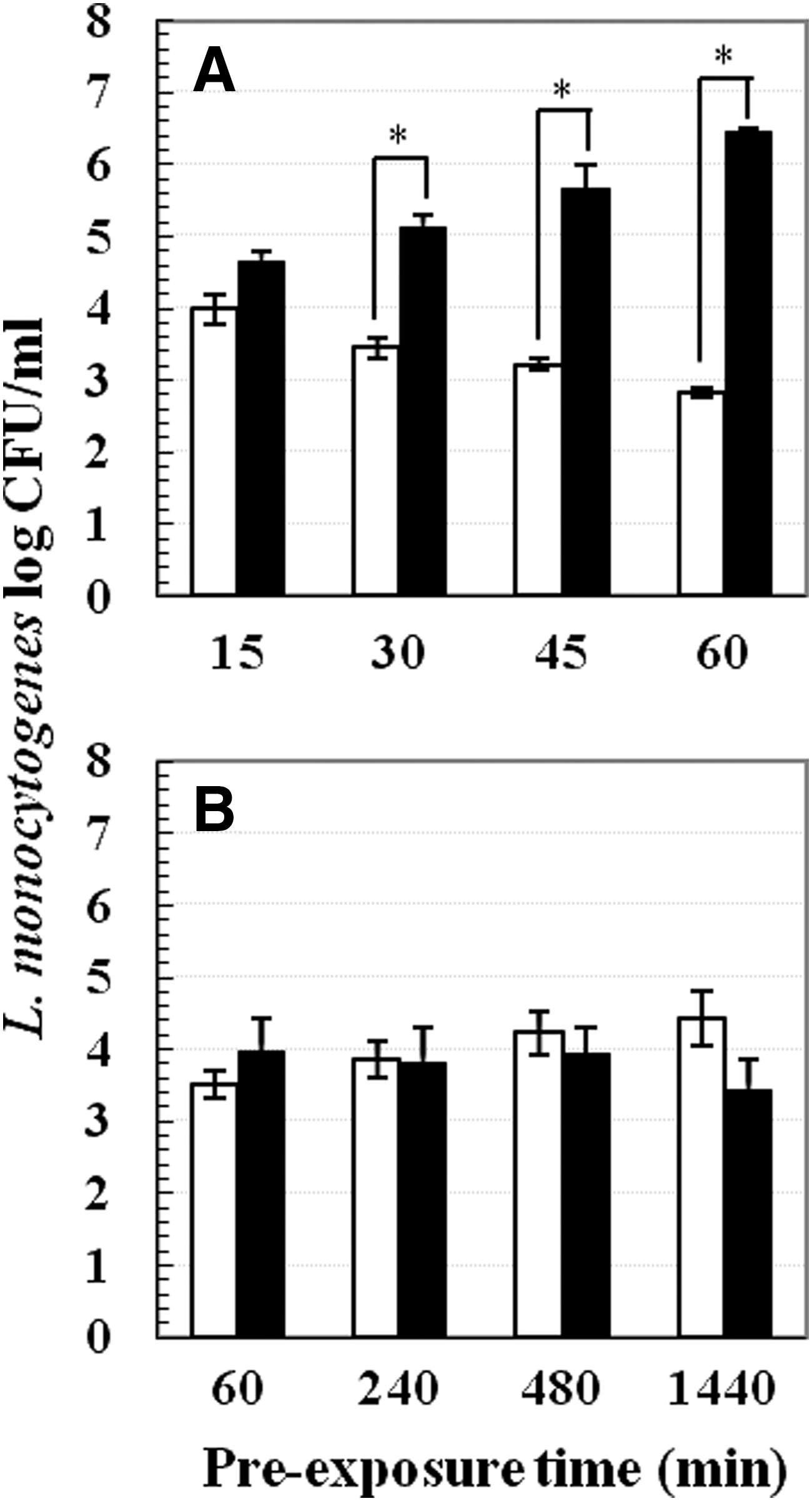
Survival of Listeria monocytogenes Scott A in lethal pH 3.5 tryptic soy broth supplemented with 0.6% yeast extract at 22°C for 60 min after pre-exposure to sublethal pH 5.0 (■) or pH 7.2 (□) at two temperatures:
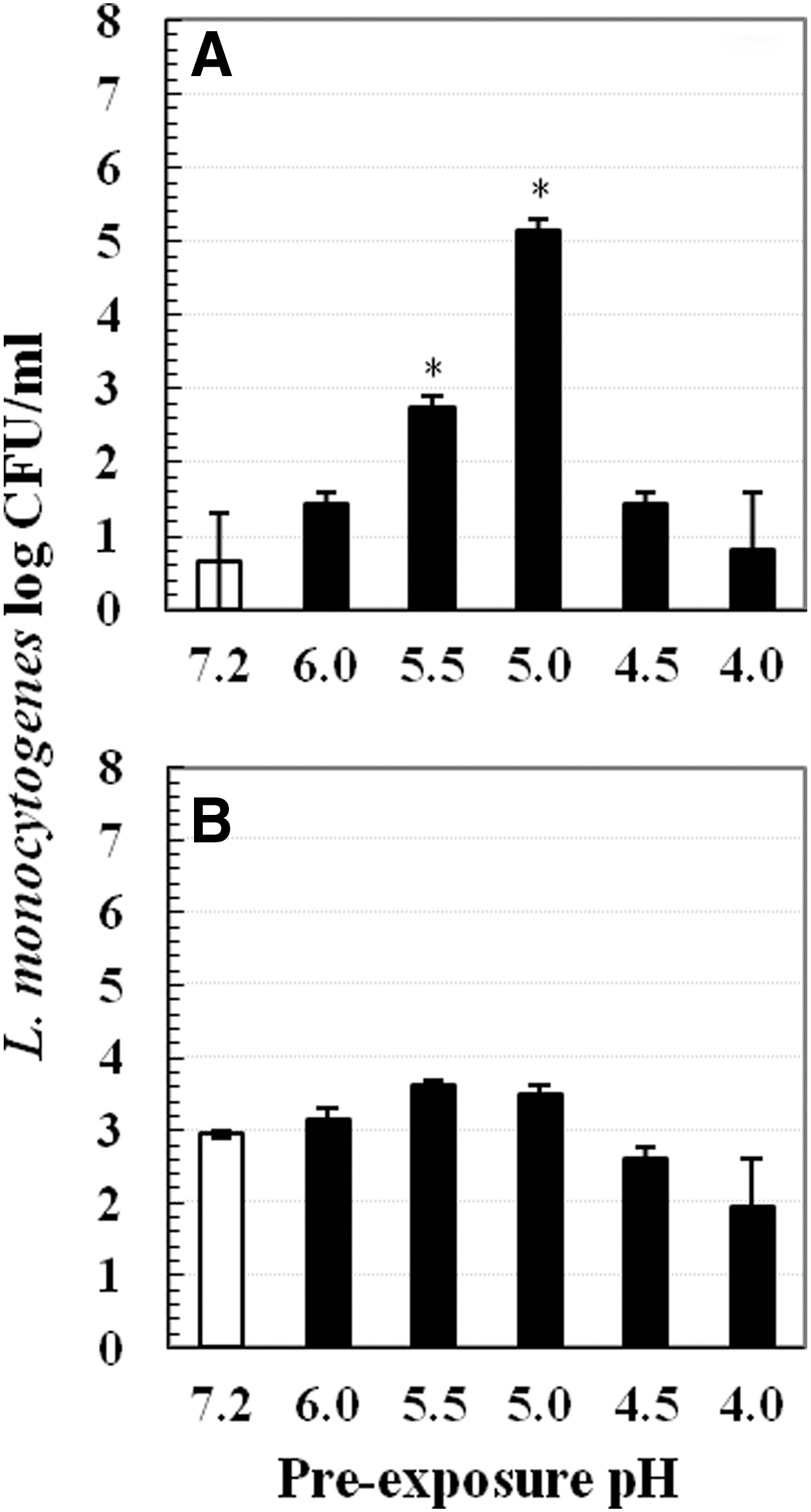
Survival of Listeria monocytogenes Scott A in lethal pH 3.5 tryptic soy broth supplemented with 0.6% yeast extract at 22°C for 90 min after 1 h pre-exposure to different sublethal pH at two temperatures:
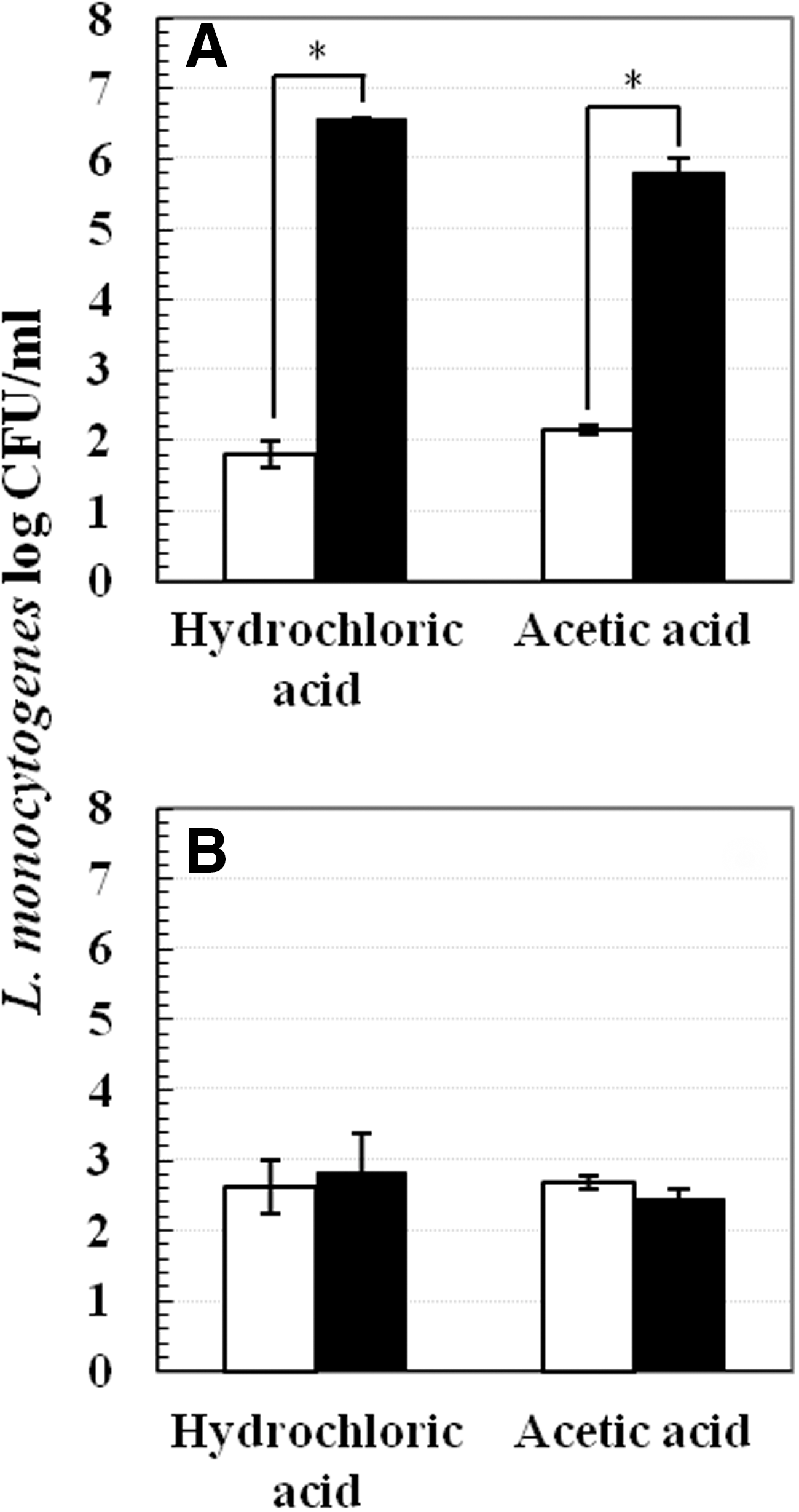
Survival of Listeria monocytogenes Scott A in lethal pH 3.5 tryptic soy broth supplemented with 0.6% yeast extract at 22°C for 90 min after 1 h pre-exposure to sublethal pH 5.0 (■) or pH 7.2 (□) at two temperatures:
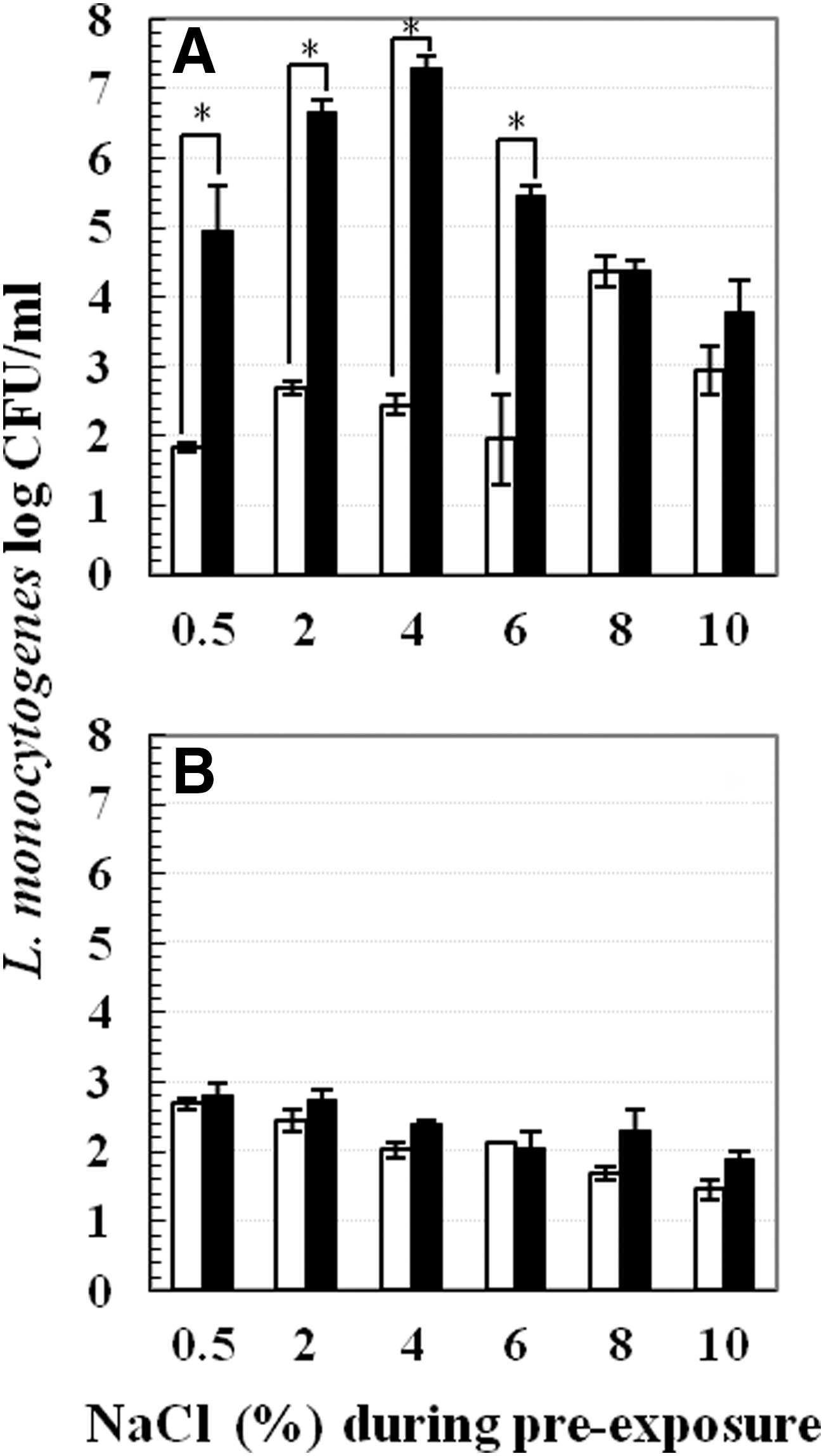
Survival of Listeria monocytogenes Scott A in lethal pH 3.5 tryptic soy broth supplemented with 0.6% yeast extract at 22°C for 90 min after 1 h pre-exposure to sublethal pH 5.0 (■) or pH 7.2 (□) in the presence of different concentrations of sodium chloride at two temperatures:
Acid-stress adaptation in acidic whey at different temperatures
Fat reduced milk was heated to 85°C and coagulated by 3M lactic acid. Coagulant was strained and acidic whey (pH ∼5.1) was collected. L. monocytogenes Scott A early log-phase cells were diluted in TSB-YE with 1:10 and centrifuged to collect cell pellets. Thereafter, cell pellets were resuspended in temperature equilibrated acidic whey for 1 h at 37°C, 22°C, or 4°C. After pre-exposure, calculated 3M lactic acid quantity was directly added into those whey samples to attain final pH of 3.5. Control cells were directly inactivated in pH 3.5 acidic whey. The lethal acid challenge was performed at 22°C and survivors were measured on PALCAM agar plates supplemented with 6 mg/L ceftazidime (Difco, Detroit, MI) every 30 min up to 2 h (Fig. 7).
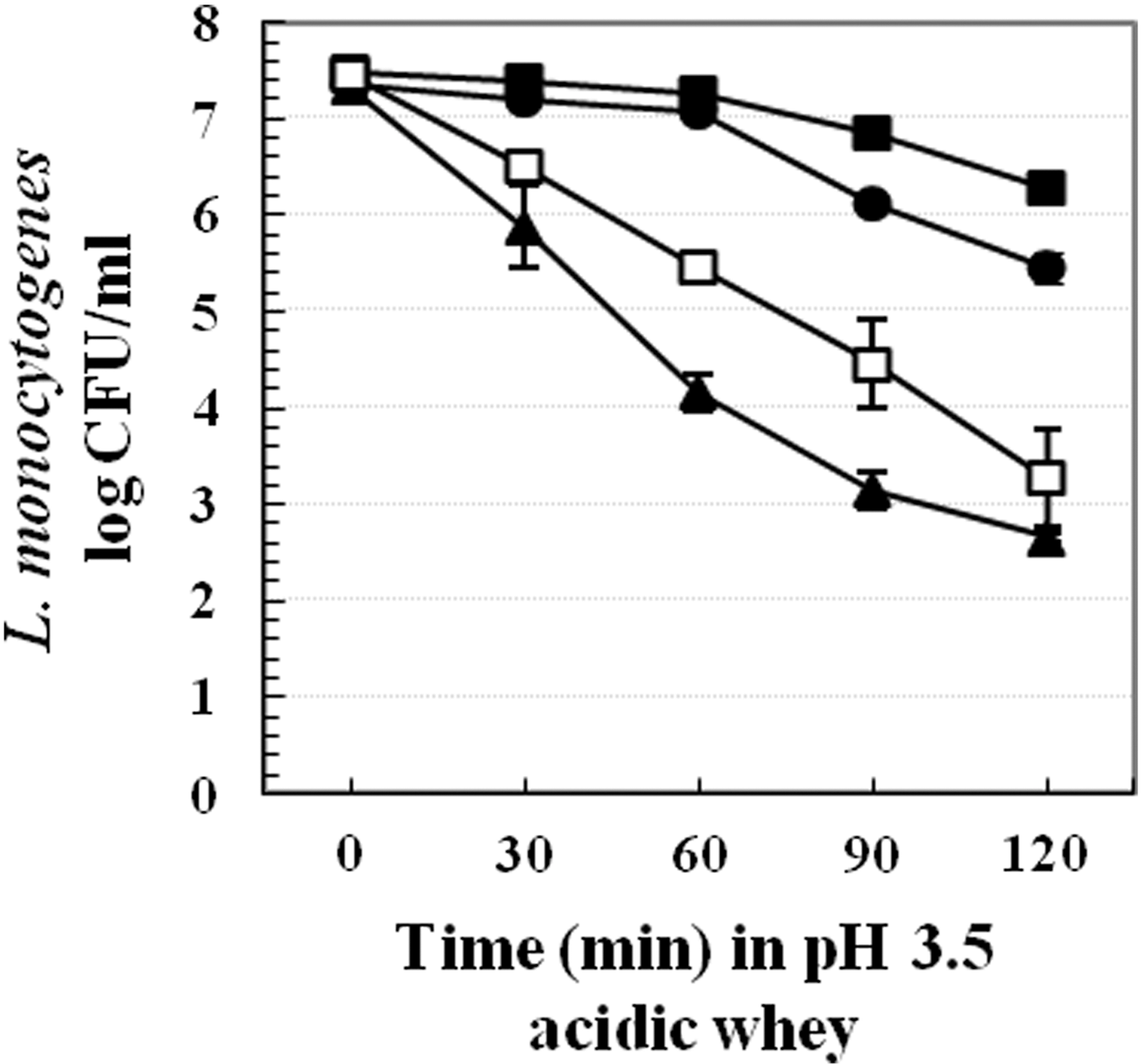
Survival of Listeria monocytogenes Scott A in lethal pH 3.5 acidic whey at 22°C. Cells were pre-exposed to acidic whey (pH 5.1) for 1 h at 37°C (■), 22°C (•), or 4°C (■). Control cells (□) were added to acidic whey and immediately inactivated by lethal pH 3.5 at 22°C. CFU, colony-forming units.
Effect of bead beating on the induction of acid-stress adaptation at 4°C
Early log phase culture of L. monocytogenes Scott A (1.2 mL) was transferred into screw-cap tubes containing 2 g of 0.1 mm zirconium (BioSpec Products, Inc., OK) beads. After 2 min of bead beating (4800 oscillations/min) at room temperature, 100 μL cultures were transferred into 900 μL pre-acidified (mild acid-stress) or neutral broth (nonstress) that was initially kept at 37°C or 4°C for temperature equilibration. Pre-exposure was allowed for 1 h at 37°C or 4°C, after which pellets were recovered and resuspended in pH 3.5 TSB-YE (3M lactic acid) at 22°C. Survivors were measured every 30 min up to 90 min (Fig. 8).
Survival of 2-min bead-beated Listeria monocytogenes Scott A cells in lethal pH 3.5 tryptic soy broth supplemented with 0.6% yeast extract at 22°C after 1 h pre-exposure to sublethal pH 5.0 (■) or pH 7.2 (□) at two temperatures:
Statistical analysis
All experiments were performed in three replicates with a completely randomized design structure. Data shown here are mean average (±SE) of three individual trials. Student t-test (p<0.05) was performed to determine significant mean separation between survival of mild acid-stressed and nonstressed cells (SPSS version 12.0, Chicago, IL).
Results
Mild acid-stress at 4°C did not induce acid-stress adaptation in L. monocytogenes
Figure 1 shows the acid adaptation of 37°C grown cells of L. monocytogenes Scott A and BUG 600, respectively, after 1 h mild acid pre-exposure at 37°C, 22°C, and 4°C. Two distinct patterns of acid adaptation were observed: (1) conditions where acid adaptation occurred; and (2) conditions where acid adaptation did not occur. For both strains, cells pre-exposed to mild acid stress at 22°C or 37°C for 1 h exhibited about 2–5 log colony-forming units (CFU)/mL better survival than nonstressed cells (Fig. 1A, 1B, 1D, 1E). In contrast, no difference in survival between mild acid-stressed and nonstressed cells was observed if pre-exposure was performed at 4°C (Fig. 1C, 1F). For Scott A, 4°C mild acid-stressed had 2–3 log CFU/mL lower survival compared to 22°C or 37°C mild acid-stressed cells after 90 min in pH 3.5 TSB-YE. For BUG 600, this difference was about 2 log CFU/mL. This conceptual finding was also true if the lethal acid challenge was performed at 4°C or 22°C (data not shown).
Lack of mild acid adaptation at 4°C was also observed in L. monocytogenes cells grown at 4°C (Fig. 2A and 2B). For both strains, no difference existed between survival of mild acid-stressed and non-stressed cells if mild acid pre-exposure was performed at 4°C. However, at 37°C, mild acid-stressed cells had about 2 log CFU/mL higher survival compared to nonstressed cells.
Increase in pre-exposure time, varying pH of mild acid exposure, change of acidulants, and addition of salt did not induce acid-stress adaptation in L. monocytogenes at 4°C
Increasing mild acid pre-exposure (pH 5.0) time from 1 h to 4, 8, or 24 h also failed to induce acid adaptation at 4°C (Fig. 3B). Survival of both mild acid-stressed and non-stressed cells was approximately 4 log CFU/mL. In contrast, 30 min of pre-exposure to pH 5.0 at 37°C induced acid adaptation as mild acid-stressed cells exhibited 1.5 log CFU/mL higher survival than nonstressed cells (Fig. 3A).
Change of mild acid pre-exposure pH also did not induce acid adaptation at 4°C. Survival of pH 4.0, 4.5, 5.0, 5.5, and 6.0 mild acid-stressed cells were 2–3.5 log CFU/mL, which was not higher than that of nonstressed cells (3 log CFU/mL) (Fig. 4B). However, pH 5.0 and pH 5.5 pre-exposure at 37°C conferred L. monocytogenes with 2–4 log CFU/mL greater survival than nonstressed cells (Fig. 4A).
No acid adaptation was observed after pre-exposure to hydrochloric acid or acetic acid at pH 5.0 at 4°C. There was about 3 log CFU/mL survival for 4°C mild acid-stressed cells, which was similar to that of 4°C nonstressed cells (Fig. 5B). However, at 37°C, mild acid-stressed cells exhibited approximately 4 log CFU/mL higher survival than nonstressed cells (Fig. 5A).
Addition of sodium chloride during pre-exposure did not induce acid adaptation at 4°C, as survival of non-acid-stressed cells was similar to that of mild acid-stressed cells (Fig. 6B). In contrast, pronounced acid adaptation was observed when the cells were pre-exposed to pH 5.0 supplemented with sodium chloride at 37°C. In the presence of low concentrations (0.5%–6%) of sodium chloride, mild acid-stressed cells exhibited at least 3 log CFU/mL greater survival than non-acid-stressed cells (Fig. 6A).
No acid-stress adaptation of L. monocytogenes in acidic whey at 4°C
Cells pre-exposed to acidic whey at 22°C or 37°C showed at least 2 log CFU/mL greater survival compared to control cells. In contrast, cells pre-exposed in acidic whey at 4°C exhibited somewhat impaired survival compared to nonstressed cells (Fig. 7). This indicated that L. monocytogenes acid adaptation occurred in acidic whey at 37°C or 22°C but not at 4°C.
Bead-beating of L. monocytogenes cells induced partial acid-stress adaptation at 4°C
In this assay, we limited the bead-beating time for 2 min, which reduced the cell viability from the initial 7.2 to 6.2 log CFU/mL. Bead-beated cells induced acid adaptation after pre-exposure to pH 5.0 TSB-YE at 4°C since about 2 log CFU/mL survival difference between mild acid-stressed and nonstressed cells was observed (Fig. 8B). However, at 37°C, 4–5 log CFU/mL greater survival was observed in mild acid-stressed cells compared to nonstressed cells (Fig. 8A).
Discussion
In the present study, we used log-phase L. monocytogenes cells instead of stationary-phase cells. One reason is that stationary-phase cells become partially acid adapted due to the drop of pH during their growth into stationary stage (Davis et al., 1996). Another reason not to work with stationary-phase cells is that after entering stationary phase, cells become naturally resistant because of the activation of stringent response (mediated by (p)ppGpp) and general stress response (mediated by σB) (Hecker and Volker, 1998; Ferreira et al., 2001; Godfrey et al., 2002). Such responses occurring in stationary phase can make L. monocytogenes resistant to various types of physiological stresses including lethal acid stress.
Ivy et al. (2012) and Samelis et al. (2003) reported that L. monocytogenes stationary-phase cells propagated at 7°C or 10°C showed lower acid tolerance than optimal-temperature-grown stationary-phase cells. Both studies used L. monocytogenes cells that were grown to stationary phase in which growth medium pH drops to 5.6–6.0 due to glucose metabolism (Davis et al., 1996). Such drop of media pH can simulate the condition of mild acid pre-exposure in stationary phase. Therefore, such reduced acid tolerance in cold-grown stationary cells observed in these studies could be partially due to repressed acid adaptation under cold temperatures.
Several authors have proposed that the activation of bacterial acid adaptation requires de novo proteins synthesis (Davis et al., 1996; O'Driscoll et al., 1996). It is possible that acid shock proteins (i.e., GAD and ADI systems) are not synthesized during mild acid pre-exposure at 4°C. There are currently no specific studies that evaluate mild acid treatment modulated genes as temperature-dependent effect. In two relevant studies, Chan et al. (2007) and Ivy et al. (2012) investigated the differential expression of 4°C, 7°C, and 37°C grown stationary phase L. monocytogenes cells. Since stationary-phase cells can cause some inherent acid adaptation due to decrease in growth medium pH, changes in gene expression profile in such physiological state may provide some additional relevant clues of temperature-dependent behavior of acid shock genes/proteins. Interestingly, these studies suggested down-regulation of GAD and ADI genes in cold-grown stationary-phase cells (Chan et al., 2007; Ivy et al., 2012). In another study, it was observed that GAD and ADI genes were also not induced when cells were exposed to organic acids at 7°C (Stasiewicz et al., 2011). Therefore, there may be a lack of synthesis of some critical acid shock proteins when cells are exposed to mild acidic condition under cold temperatures.
Being exposed to pH 5.0 at 4°C, L. monocytogenes encounters two different types of physiological stress, namely, acid stress and cold stress. L. monocytogenes can adapt to a cold environment by expressing different protein patterns that contribute to its modulated metabolism pathway, nutrient uptake, protein folding, and lipid biosynthesis (Cacace et al., 2010). Hence, it is likely that between acid and cold adaptation, cold adaptation becomes the priority task for the bacterium to deal with. We hypothesized that L. monocytogenes could start to respond to acid stress after cells were fully cold adapted. We therefore tested whether cold-grown log phase cells that are fully cold acclimatized can show acid adaptation at 4°C. However, cold-grown log cells still failed to activate acid adaptation at 4°C (Fig. 2). Therefore, the actual cold adaptation event may not be the sole factor for repressed acid adaptation at 4°C.
Cold stress environment may block the penetration of acid into cytoplasm and subsequently fails to trigger intracellular responses to acid stress. To test this, we used a simple bead-beating approach as an alternative for sonication. Bead beating is routinely applied in proteomic or microarray work to break the bacterial cell envelope to yield more protein or mRNA extraction. Our data demonstrated that bead beating of L. monocytogenes cells could partially induce acid adaptation at 4°C (Fig. 8B), whereas the magnitude of acid adaptation at 4°C was still lower than what occurred at 37°C (Fig. 8A). It is possible that blockage of acid entry into cytoplasm of L. monocytogenes cells with an intact cell envelope could be one of the factors for repressed acid adaptation at 4°C. A proteomic analysis on cold-adapted L. monocytogenes showed that cell wall synthesis was up-regulated at 4°C (Cacace et al., 2010). Another study on Staphylococcus spp. revealed that 4°C cold-stressed phenotypes possessed a thicker cell wall compared to normal cells (Onyango et al., 2012). Such cell morphological modifications could build the physiological barriers to prevent acid from entering cytoplasm under cold environment and eventually impede the process of acid adaptation. On the other hand, since cell envelope blockage partially prevents the induction of acid adaptation, it is necessary to investigate other possible mechanisms to have a comprehensive understanding of the cold temperature effect on acid adaptation in L. monocytogenes.
Conclusions
Acid-stress adaptation that typically occurs in L. monocytogenes at 22°C or 37°C was not induced when cells were pre-exposed to sublethal acid at 4°C. Our findings suggest that cold processing or cold storage temperature can prevent the risk of creating acid-stress resistant phenotypes of L. monocytogenes.
Footnotes
Acknowledgments
This research was supported in part by the Strategic Research Initiative and Food Safety Initiative awards to RN by the Mississippi Agricultural and Forestry Experiment Station (MAFES) under project MIS-401160.
Disclosure Statement
No competing financial interests exist.