
Abstract
Salmonella enterica subsp. enterica serovar 4,[5],12:i:- DT193 is recognized as an emerging monophasic variant of Salmonella Typhimurium in many European countries. Resistance to ampicillin, streptomycin, sulphonamides, and tetracycline (R-type ASSuT) is described as one of the most common profiles of resistance within this clone. Recently, strains presenting such features were isolated from two unrelated outbreaks in Italy. Strains were characterized by pulsed-field gel electrophoresis (PFGE), performed with XbaI, BlnI, and SpeI, and multiple-locus variable-number tandem repeat analysis (MLVA). XbaI-PFGE showed strains related to the two outbreaks as indistinguishable. Conversely, both BlnI-PFGE and MLVA characterized the strains related the two outbreaks as different. XbaI-PFGE identified two profiles, differing by one band, within strains isolated from one of the two outbreaks. Also BlnI-PFGE and MLVA generated different profiles among the strains related to that outbreak. Combining the PFGE profiles obtained by XbaI and BlnI and comparing them with the MLVA profiles, the two methods grouped the same isolates based on identity. Moreover, genomic deletions of the genes included in the operon fljAB, the flanking iroB gene, and the closely located STM2757 gene were investigated. For all strains, the same profile of deletion characterized by the absence of fljA, fljB, and hin genes and the presence of STM2757 and iroB genes was identified. This profile of deletion represents a mixture between two profiles of Salmonella 4,[5],12:i:- described as the “Spanish” and the “U.S.” clones. This study demonstrated that although strains of Salmonella 4,[5],12:i:- DT193 ASSuT are highly clonal, minor differences between strains may be seen during the same outbreak by using in parallel PFGE with different restriction enzymes, MLVA, and the analysis of molecular markers related to the operon fljAB. The combination of these different molecular approaches was essential to clarify the epidemiological relationship among the strains.
Introduction
S
Salmonella 4,[5],12:i:- has been responsible for a remarkable number of human outbreaks worldwide (Echeita et al., 1999; Tavechio et al., 2004; Mossong et al., 2007; Switt et al., 2009; Bone et al., 2010; Gossner et al., 2010; CDC, 2011, 2012; Raguenaud et al., 2012). These episodes clearly demonstrated that monophasic variants were as virulent as Salmonella Typhimurium (EFSA, 2010).
In Italy, the proportion of Salmonella 4,[5],12:i-, among all strains of veterinary origin notified to the National Reference Laboratory for Salmonella in the framework of the Enter-vet network between 2002 and 2010, increased from 3.5% (2002–2004) to 10.3% (2010) and, in this last year, Salmonella 4,[5],12:i:- was the most common serovar isolated from such samples (Barco et al., 2012a). The vast majority of these monophasic strains were from pigs and pork products, followed, with a lower prevalence, by isolates obtained from poultry and cattle, whereas isolation from other sources was rather rare (Barco et al., 2012b). In humans, at a national level, Salmonella 4,[5],12:i:- has been the third most frequently isolated serovar from 2004 to 2008 (Dionisi et al., 2009), and since 2009 it even has exceeded Salmonella Enteritidis and has reached the second position after Salmonella Typhimurium (
Isolates of Salmonella 4,[5],12:i:-, belonging to phage-type DT 193 and expressing resistances to ampicillin, streptomycin, sulphonamides, and tetracycline (R-type ASSuT), have emerged rapidly in several European countries, and this profile has become one of the most dominant clones within this serovar (Hopkins et al., 2012). Salmonella 4,[5],12:i:- DT 193 ASSuT presenting the PFGE type STYMXB.0131 has caused great concern, since it was responsible for two outbreaks in Luxemburg in 2006 with 133 confirmed cases, 24 hospitalizations, and 1 death (Mossong et al., 2007). It was also the most common profile among Salmonella 4,[5],12:i:- strains isolated in Germany from pig, pork, and humans (Hauser et al., 2010), as well as from swine and human strains in England-Wales, France, Germany, Italy, Spain, and The Netherlands (Hopkins et al., 2010), and it was also described among strains isolated from humans and foodstuffs in Switzerland (Gallati et al., 2013).
This study reports the isolation of Salmonella 4,[5],12:i:-, ASSuT, DT193 STYMXB.0131 strains from two epidemiologically unrelated outbreaks in Italy. The strains were subtyped by PFGE, using three different restrictions enzymes (XbaI, BlnI, SpeI), and by multiple-locus variable-number tandem repeat analysis (MLVA), to confirm the outbreaks and to investigate whether they were caused by the same clone. Moreover, since different profiles of deletion of the genes responsible for the expression of phase 2 flagellar antigen have been described, the presence/absence of the genes of the operon fljAB as well as other closely located genes (iroB and STM2757) were mapped as additional molecular markers to characterize the strains related to the outbreaks.
Materials and Methods
Salmonella isolates and description of outbreaks
The study panel consisted of 27 Salmonella 4,[5],12:i:-strains, isolated from two unrelated outbreaks (hereafter called outbreak A and outbreak B), that occurred in 2009 and 2010 in two regions of Northeast and Northwest Italy, respectively.
Strains associated with outbreak A and analyzed in the present study consisted of four isolates from a cat and its owners (an adult and a child). All strains were obtained from fecal material. The infection likely occurred as a result of the close contact between the householders and the companion animal.
Outbreak B occurred among guests who had eaten at a rural guesthouse-restaurant. As already demonstrated (Mürmann et al., 2009), many opportunities for contamination during cutting, preparation, and handling of pork meat products are present, and outbreak B was probably due to the consumption of contaminated pork products produced in the restaurant and obtained from infected animals.
Salmonella was isolated from 70 persons among the 153 guests, who ate at that restaurant in the period of the outbreak and showed clinical symptoms of salmonellosis. Moreover, Salmonella was isolated from an additional nine people, including the owners of the restaurant, some relatives, and employees.
The strains related to outbreak B investigated in the present study were obtained from cases (four strains isolated from four patients), from animals kept in facilities close to the restaurant and reared to produce foodstuffs served there (one strain from poultry feces and 10 strains from swine feces) and from seasoned and raw pork products collected in the restaurant and produced using meat obtained from animals reared in the close facilities (eight strains from fresh and seasoned pork products). The strains were isolated from animal and food samples collected at the restaurant immediately after the notification of the initial human cases. In Table 1, the number and the origin of the strains studied in each outbreak are reported.
STTR 9-5-6-10-3.
Laboratory methods
Phenotypic characterization
The strains were serotyped according to the Kauffman-White scheme combining the traditional slide agglutination method and polymerase chain reaction (PCR) (Barco et al., 2011). All strains were tested for phage susceptibility using the protocol reported by Anderson et al. (1977) and following the interpretative guidelines defined for Salmonella Typhimurium by the Health Protection Agency (Colindale, UK). Antimicrobial susceptibility was evaluated using the disk diffusion method according to the criteria established by the CLSI (2010). The antimicrobials (Becton Dickinson) tested were: ampicillin (10 mg), cefotaxime (30 mg), chloramphenicol (30 mg), ciprofloxacin (5 mg), gentamicin (10 mg), kanamycin (30 mg), nalidixic acid (30 mg), streptomycin (10 mg), sulfonamides (250 mg), tetracycline (30 mg), and trimethoprim–sulfamethoxazole (1.25–23.75 mg). Escherichia coli ATCC 25922 was used as control strain.
PFGE
PFGE was carried out according to the PulseNet standardized protocol (Ribot et al., 2006). Genomic DNA was obtained and subjected to macrorestriction analysis with XbaI, SpeI, and BlnI restriction enzymes (Roche, Maylan, France). The generated fragments were separated by using the CHEF MAPPER XA Pulsed Field Electrophoresis System (BioRad, Hercules, CA). Agarose plugs and PFGE conditions using XbaI and BlnI were performed according to the PulseNet protocol. Macrorestriction using SpeI was carried out according to the conditions suggested by Thong and colleagues (1994). XbaI-digested DNA from Salmonella Braenderup H9812 was used as a molecular reference marker. PFGE profiles differing by one or more fragments were considered as distinct. Dendrogram and cluster analysis were produced using the InfoQuest™ FP version 4.5 software package (BioRad, Hercules, CA). Dice coefficient analysis and dendrograms were obtained by using the unweighted pair group method with arithmetic means (UPGMA), with a position tolerance limit of 2% and optimization of 1%. One dendrogram was generated by combining the PFGE-XbaI, PFGE-BlnI, and PFGE-SpeI patterns. The pulsotypes of the strains obtained by XbaI restriction were uploaded to the database established at the Health Protection Agency (Colindale, London, UK), compared with the profiles in the international database and named according to PulseNet Europe standards.
MLVA
MLVA was performed according to the protocol described by Lindstedt and colleagues (2004). MLVA results were reported as a string of five numbers (STTR9-STTR5-STTR6-STTR10pl-STTR3) using nomenclature suggested by Larsson and colleagues (2009).
Gene-mapping in the fljAB genomic region
The presence of the genes fljA, fljB, and hin as well as the iroB gene flanking the fljAB operon and the closely located STM2757 gene were analyzed by using the gene-specific primers and PCR conditions as previously described by Soyer and colleagues (2009). These PCR targets were respectively located at the 3’ end (genes fljA, fljB, hin [STM2772], and iroB [STM2773]) and flanked the 5‘ end (gene STM2757) of cluster V. This is a region spanning 16 genes (from gene STM2758 to gene STM2773) that were absent in a selection of Spanish Salmonella 4,5,12:i:- isolates, whose gene content was compared to Salmonella Typhimurium LT2 by Garaizar and colleagues (2002). The presence/absence of these five genes were mapped according to the approach previously described by Soyer et al. (2009).
Results
All 27 strains were classified as Salmonella 4,[5],12:i:-, and the absence of phase-2 flagellar antigen was confirmed both by traditional serotyping and by PCR. Moreover, all isolates were phage-typed as DT 193 and they all expressed the resistance profile: ampicillin, streptomycin, sulphonamides, and tetracycline (R-type ASSuT).
PFGE analysis using XbaI generated two patterns differing by one band. The most common pattern, corresponding to pulsotype STYMXB.0131, included the four strains isolated from outbreak A and 19 strains from outbreak B (one strain from poultry feces, seven from swine feces, seven from pork products, and four from humans). The remaining four strains from this last outbreak (three obtained from swine feces and one from a pork product) showed a second pattern, which differed from STYMXB.0131 by the presence of an additional band of 78 kb. The similarity between the two XbaI profiles was 96.6% (Dice and UPGMA analysis).
PFGE analysis performed with SpeI generated an indistinguishable pattern for all but one strain, differing from the dominant profile by the presence of one additional band of about 55 kb. This strain was isolated from a swine fecal sample in outbreak B. The similarity between the two SpeI profiles was 97.9%.
PFGE performed using BlnI generated three patterns and allowed the strains related to the two outbreaks to be distinguished. Twenty-three of 24 isolates from outbreak B showed the same BlnI profile; the remaining strain obtained from a human fecal sample differed by the absence of one band of about 90 kb. These two profiles showed a similarity of 96.0%. All isolates from outbreak A showed the same BlnI profile, differing from the dominant outbreak B profile by the presence of two additional bands of about 60 and 450 bp, respectively. The profile specific for outbreak A showed a similarity of 92.8% with the ones from outbreak B.
When the PFGE patterns generated by all three enzymes were considered, five different patterns were generated (Fig. 1). Four profiles were obtained for strains isolated from outbreak B (PFGE-B1, PFGE-B2, PFGE-B3, and PFGE-B4) and a unique profile was obtained from the strains isolated from outbreak A (PFGE-A1). As far as outbreak B is concerned, the dominant profile (PFGE-B1) included 17 isolates (3 from humans, 1 from poultry feces, 6 from swine feces, and 7 from pork products), whereas the remaining 6 isolates generated 3 very similar profiles (PFGE-B2, PFGE-B3, and PFGE-B4).
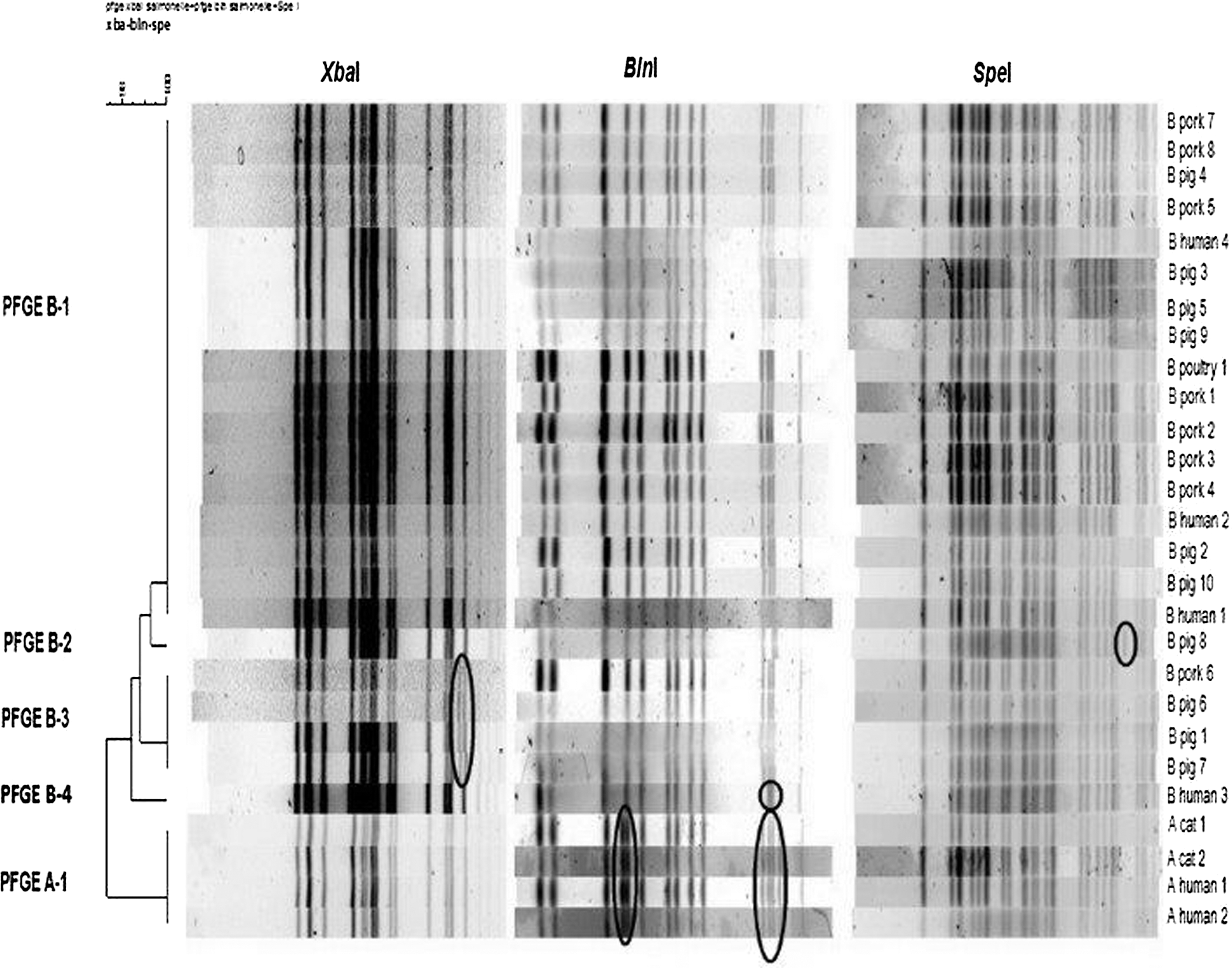
Dendrogram generated combining the pulsed-field gel electrophoresis (PFGE)-XbaI, PFGE-BlnI, and PFGE-SpeI patterns of the strains isolated from the two outbreaks (A and B).
MLVA analysis generated four different profiles. The strains from outbreak A showed the same MLVA profile: 3-13-11-0-211 (MLVA-A1). Among isolates from outbreak B, the dominant profile was 3-12-10-0-221 (MLVA-B1) and included 18 isolates. The profile 3-12-9-0-211 (MLVA-B2) was demonstrated in one strain isolated from a human fecal sample and the last profile, 3-12-8-0-211 (MLVA-B3), was demonstrated in four isolates (three strains from pig feces and one strain from pork). The profile associated with outbreak A differed in two loci (STTR5 and STTR6) from the profiles associated with outbreak B, whereas the three profiles identified for the later differed only at locus STTR6.
A close correlation was noted between the grouping by MLVA and PFGE (Table 1). The 18 isolates from outbreak B with the dominant MLVA profile (MLVA-B1) corresponded to the isolates (17) presenting the dominant PFGE profile (PFGE-B1) and the unique isolate presenting the PFGE-B2 profile. The isolates (four) presenting the MLVA-B3 profile corresponded to the ones showing the PFGE-B3 profile, and the human isolate (one) presenting the MLVA-B2 profile corresponded to the one showing the PFGE-B4 profile. The isolates from outbreak A showed the same PFGE and MLVA profiles (PFGE-A1 and MLVA-A1).
All the isolates related to the two outbreaks showed the same profile of gene deletion of cluster V. For all strains, the absence of fljA, fljB, and hin genes was detected, whereas the gene iroB was always present. Moreover, the gene STM2757, flanking the cluster V, was conserved in all strains.
Discussion
This study reported for the first time the involvement of Salmonella. 4,[5],12:i:- DT193 ASSuT pulsotype STYMXB.0131 strains in a foodborne outbreak (outbreak B) that occurred in Italy, and it reported at least one other closely related type (outbreak A) that caused disease in this country.
It also demonstrated that Salmonella 4,[5],12:i:- DT193 ASSuT strains are highly clonal, but minor differences may be seen also between the isolates involved in the same outbreak.
As a consequence of the high clonality, PFGE using XbaI did not highlight their full diversity. However, the strains investigated were clearly allocated into the two different outbreaks when an additional restriction enzyme (BlnI) was used, indicating that following this approach, PFGE remains a good choice for subtyping Salmonella 4,[5],12:i:- DT193 ASSuT strains. BlnI has already been described as a highly discriminatory enzyme to subtype clonal subtypes of Salmonella (Baggesen et al., 2000; Zamperini et al., 2007). According to the data obtained in the present study, MLVA gave equal discrimination between strains of Salmonella 4,[5],12:i:- DT193 ASSuT as PFGE. As shown for BlnI-PFGE, MLVA also identified different profiles for the isolates within the two outbreaks.
When comparing MLVA profiles from the two outbreaks, differences were identified at two (STTR5 and STTR6) of the five loci targeted. Similarly, a study characterized by MLVA isolates related to two outbreaks of Salmonella Typhimurium that occurred in England and Denmark concluded that the strains investigated were not epidemiologically linked since some differences at both of these loci were detected (Best et al., 2009).
A great deal of variability within STTR5 and STTR6 loci was also described in a study characterizing epidemiologically unrelated Salmonella Typhimurium strains obtained from humans and from different sources in the United Kingdom (Best et al., 2007), as well as a study considering Salmonella 4,[5],12:i:- strains from pig, pork, and human samples in Germany (Hauser et al., 2010) and a study investigating a selection of Spanish strains of Salmonella 4,[5],12:i:- U302 (Laorden et al., 2010). Other studies, using the same MLVA method, also described STTR10 as a variable locus (Chiou et al., 2010; Prendergast et al., 2011; Sintchenko et al., 2012).
As described in the present study, Hopkins and colleagues (2007) also reported that since MLVA is a highly discriminatory method, some minor changes in targeted loci can be evidenced among outbreak-related strains. These authors, as already indicated for Eschericha coli O157:H7 (Noller et al., 2003), proposed a difference of no more than two repeats at one single locus as cut-off to identify isolates that are part of the same outbreak. Using this cut-off also for the present study, a classification of the strains consistent with the epidemiological situation of the outbreaks described was obtained. Petersen and colleagues (2011) confirmed that among Salmonella Typhimurium strains related to a specific outbreak, changes in terms of gain or loss of a single repeat unit occurred and involved one specific locus (mainly STTR5, STTR6, and STTR10), whereas only occasionally changes involving more repeat units were described. These data indicate that MLVA yields satisfactory discriminatory power to differentiate epidemiologically unrelated strains also when highly clonal strains, like the ones described in the present study, are investigated, and it provides sufficient epidemiologic concordance to be useful in outbreak investigations.
Intriguingly, although MLVA and PFGE determine genomic diversity by very different methods (Hopkins et al., 2007), MLVA produced exactly the same subtyping picture, in terms of the correlation among the strains, as PFGE when it was performed combining different restriction enzymes. Therefore, since MLVA is easier and more rapid than PFGE (Kurosawa et al., 2012), MLVA might be the preferable subtyping method. However, it presents a main drawback when compared to PFGE. It is a relatively new method and the historical data available are quite limited. Conversely, PFGE datasets containing data in existence for many years and from different geographic areas have been available, and they provide a valuable tool to interpret the profiles and define an acceptable level of variability for detecting possible clustered cases (Barrett et al., 2006).
As far as the fljAB operon is concerned, the strains investigated in the present study lacked the fljA, fljB, and hin genes. The same profile of deletion was also identified as the most prominent for a selection of German S. 4,[5],12:i:- strains obtained from swine and human samples (Hauser et al., 2010) and French Salmonella 4,[5],12:i:- strains (Bugarel et al., 2012).
In a selection of Spanish Salmonella 4,[5],12:i:- U302 strains, the genes of the fljAB operon, as well as a part or the entire gene flanking the operon (iroB), were lacking (Laorden et al., 2010). In the strains described in the present study, the gene iroB was always conserved. A study investigating American Salmonella 4,[5],12:i:- isolates reported different profiles of deletion for the fljAB operon, but the great majority of the strains investigated presented complete and partial deletions of fljB, whereas the hin gene was conserved in almost all isolates (Zamperini et al., 2007). This finding was confirmed later by Soyer and colleagues (2009), who compared the profiles of deletion of a selection of genes included in the genomic region defined by Garaizar and colleagues (2002) as cluster V (from gene STM2758 to gene STM2773 of Salmonella Typhimurium LT2 and including the fljAB operon). These authors, who investigated a collection of Salmonella 4,[5],12:i:- strains obtained in United States and Spain, confirmed that the Spanish isolates lacked the fljA, fljB, hin, and iroB genes and conserved the STM2757 gene, whereas the American isolates lacked the fljA, fljB, and STM2757 genes, but conserved hin and iroB genes.
The strains described in the present article showed a profile of deletion of cluster V that is a mixture between the “Spanish” and the “U.S.” clones. The Italian deletion profile is characterized by the absence of the hin gene and the presence of the STM2757 gene, as for Spanish strains, but conserved the iroB gene as for the American clone.
This study highlights that pathogenic strains of Salmonella 4,[5],12:i:- DT193 ASSuT also circulate in Italy, as in other European countries. Moreover, the study demonstrated that, despite strains presenting this profile are highly clonal, some molecular differences can also be evidenced among epidemiologically related strains. Moreover, as already demonstrated, the combination of different molecular methods (Bessa et al., 2007) can be valuable to characterize highly clonal strains and to investigate their epidemiological relationship.
Footnotes
Acknowledgment
The authors are grateful to Dr. Gabrialla Conedera for providing part of the strains characterized in the present study.
Disclosure Statement
No competing financial interests exist.