
Abstract
Background:
The growth of Escherichia coli O157:H7 in contaminated dairy and other refrigerated food products due to temperature fluctuation poses a major food safety threat. Effective control or inhibition of E. coli O157:H7 growth depends on our understanding of mechanisms that regulate its growth at low temperature. We hypothesized that polynucleotide phosphorylase (PNPase) plays a critical role in E. coli O157:H7 low-temperature growth.
Methods:
To test this hypothesis, the pnp deletion mutant of E. coli O157:H7 was generated using the λ Red recombinase system, and the growth and survival of wild-type and pnp deletion mutant strains were compared at low temperatures.
Results:
The growth of pnp deletion mutant strains in Luria Broth (LB) and agar plate at 37°C was similar to their corresponding wild-type strains, while the deletion of pnp impaired E. coli O157:H7 growth in LB at 10°C and 22°C; growth impairment could be partially recovered in the mutant strains by ectopic expression of the pnp complementation plasmid, demonstrating that growth impairment was PNPase-specific. During 14 days of 10°C storage in both LB and milk, wild type strain EDL933 grew and reached >8 log10 colony-forming units per milliliter after 4 days of 10°C storage, while EDL933Δpnp gradually died off with effects more pronounced in milk, which were again mitigated by pnp overexpression. In addition, pnp deletion impaired the motility of E. coli O157:H7 but did not affect its susceptibility to H2O2.
Conclusion:
PNPase is required for the growth of E. coli O157:H7 at low temperature; PNPase thus provides a molecular target to control the growth of E. coli O157:H7, which may have important practical applications in dairy and other food industry.
Introduction
A
Besides pasteurization, refrigerated storage is the primary method for controlling bacterial growth in milk (Okamura et al., 2010). Though E. coli O157:H7 cannot grow at refrigerated temperature (5°C), it can grow at a temperature as low as 8°C (Palumbo et al., 1995; Wang et al., 1997). However, in reality, the temperature fluctuation is routine for dairy products during its storage, transport, retail display, and at home, often bringing milk temperatures close to 10°C or higher. Therefore, it is critical to control E. coli O157:H7 growth in milk and dairy products above refrigeration temperature, which depends on our understanding of mechanisms regulating its cold adaption and growth.
Polynucleotide phosphorylase (PNPase) has three identical subunits and is a component of multiprotein complex called the RNA degradosome (Carpousis, 2007). Each subunit contains two RNase PH-like domains linked by an α helical domain, a KH domain, and an S1 domain (Awano et al., 2008). RNase PH-like domains and the α helical domain are responsible for the PNPase enzymatic activity, while the KH and S1 domains are mainly responsible for RNA binding (Briani et al., 2007). In generic E. coli, PNPase plays a critical role in its cold acclimation at 15°C (Zangrossi et al., 2000; Beran et al., 2001), and the RNase activity of PNPase is important for its growth at a low temperature (Awano et al., 2008). Despite its positive role in cold adaptation of generic E. coli as low as 15°C, the roles of PNPase in the survival and proliferation of E. coli O157:H7 have not been tested. Neither the survival of generic E. coli nor E. coli O157:H7 at a cold temperature relevant to refrigerated foods (4°C and 10°C) has been evaluated.
To elucidate the function of PNPase in E. coli O157:H7 growth at low temperatures, we deleted the pnp gene from E. coli O157:H7 genomic DNA, which was then complemented with PNPase overexpression. The growth and survival of the resulting mutants were compared to those of wild-type E. coli O157:H7 in different environments and storage temperatures.
Materials and Methods
Bacterial strains and plasmids
The E. coli O157:H7 strains and plasmids used in this study are listed in Table 1. Bacteria were routinely grown in Luria broth (LB) at 30°C or 37°C.
MSU, Michigan State University; STEC,
Construction of E. coli O157:H7 pnp deletion mutants and pnp complementary plasmid
E. coli O157:H7 pnp deletion mutant was generated per the published method (Datsenko et al., 2000). Briefly, using the forward primer GGCTTTACCCACATAGAGCTGGGTTAGGGTTGTCATTAGTCGCGAGGATGattccggggatccgtcgacc, and the reverse primer: 5' CCGCCGCAGCGGAYGGCAAATGGCAACCTTACTCGCCCTGTTCAGCAGC tgtaggctggagctgcttcg. Kanamycin resistance cassette with pnp homology flanking sequence (uppercase) was polymerase chain reaction (PCR)-amplified from pKD13 and electroporated into E. coli O157:H7 containing the λ-Red recombinase plasmid. The mutation of pnp was confirmed by PCR amplification using primers external to the disrupted gene (forward: TGTCATTAGTCGCGAGGATG; reverse: GCGGAYGGCAAATGGCAACC), and primers target to pnp gene region (forward: AGCGTGTTTCGCTTTCTCGCCAT; reverse: CGTGCCCTGACCGAAGAAACTGG, product size is 96 bp). Deletion of pnp was further confirmed by Western blotting (see immunoblotting section for details).
To construct the pnp complementary plasmid, the pnp was PCR-amplified from E. coli O157:H7 using primers AT
PNPase immunoblotting
Overnight cultures of E. coli O157:H7 strains were (1:100) inoculated to LB and grown at 25°C for 4 h, when arabinose was added to a final concentration of 0.2% (wt/vol) to induce pnp expression, because the promoter of the expression vector, pBAD, is inducible by arabinose. Bacteria were further cultured at 25°C for an additional 5 h and used for protein extraction. Protein extracts were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis, transferred to nitrocellulose membranes, and assayed with antibodies specific to PNPase (Thermal, Rockford, IL).
Growth of E. coli O157:H7
First, E. coli O157:H7 strains were streaked on LB plates, then incubated at 37°C, 22°C, and 10°C for 12 h, 24 h, and 168 h, respectively. Next, the growth curves of E. coli O157:H7 of different strains were further analyzed. Briefly, E. coli O157:H7 strains (1.0×105 colony-forming units [CFU]/mL) were inoculated into the 96-well microplates (Costar, Tewksbury, MA) and incubated at 37°C or room temperature (22°C). Optical density (OD) values were read hourly at 600 nm using an SM5 spectrometer (Molecular Device, Sunnyvale, CA). Growth curves were constructed using OD600nm versus incubation time. Each strain had six replicates.
Growth and survival of E. coli O157:H7 strains in LB during storage at 10°C or 4°C
One hundred microliters of E. coli O157:H7 cultures (1.0×107 CFU/mL) were inoculated into 10 mL of LB and subsequently stored at 10°C or 4°C for 14 days. Survival and growth of E. coli O157:H7 strains were measured every other day, with four replicates per strain per storage temperature. On each sampling day, 0.5-mL samples were aseptically transferred, serially diluted in 1×phosphate-buffered saline and plated on LB agar to enumerate the E. coli O157:H7 colonies, which were expressed as log10CFU/mL.
Growth and survival of E. coli O157:H7 strains in commercial milk during 10°C or 4°C storage
On the day of inoculation, fresh stocking commercial whole milk was purchased from a local store and transported to the lab in a cooler within 10 min. One hundred microliters of E. coli O157:H7 cultures (1.0×107 CFU/mL) were inoculated into 10 mL milk and subsequently stored at 10°C and 4°C for 14 days. Survival and growth of E. coli O157:H7 strains were analyzed as described above.
Swimming motility analysis
Migration in semisolid agar, an indicator of swimming motility, was assayed as previously described (Ryjenkov et al., 2006). Briefly, overnight cultures were inoculated 1:100 in LB and grown to log phase, when 0.2% arabinose was added to induce pnp expression. After 4-h incubation at 37°C, 2-μL culture was spotted onto 0.25% swimming soft agar plates containing 0.2% arabinose. Swimming zones were measured in millimeters after 5 h of incubation at 37°C.
Cell viability test in LB plate with various H2O2 concentrations
Overnight E. coli O157:H7 cultures were 1:100 diluted in LB and incubated at 37°C until the log phase. The exponentially growing cultures were diluted 1:10; then they were further serially diluted fivefold. Two microliters of serially diluted cultures were spotted on the surface of LB agar plates containing 0%, 0.00125%, 0.0025%, or 0.005% of H2O2 (Sigma). Cell growth on the spots was recorded after overnight incubation at 37°C (Wu et al., 2009).
Statistical analysis
Data were analyzed using the General Linear Model procedure (SAS, 2000). Means±standard deviation were reported. Means were separated by least significant difference test multiple comparison with significance at p<0.05. The enumerated E. coli O157:H7 CFU was log-transformed.
Results
Confirmation of pnp knockout in E. coli O157:H7
An E. coli O157:H7 pnp mutant was confirmed by PCR, resulting in 2202-bp and 1303-bp PCR products in wild-type and pnp deletion mutant strains, respectively, which were consistent with the expected sizes (Fig. 1). The pnp deletions were further verified by PCR targeting the pnp gene region (Fig. 1B) and PNPase Western blotting (Fig. 1C). Clearly, pnp is absent at both transcriptional and translational levels in EDL933Δpnp. Deletion of pnp in E. coli O157:H7 decreased its swimming motility, which was complemented by overexpressing pBAD::pnp (Fig. 2A). Transformation of pBAD::pnp to EDL933Δpnp resulted in a high level of PNPase (Fig. 2B).
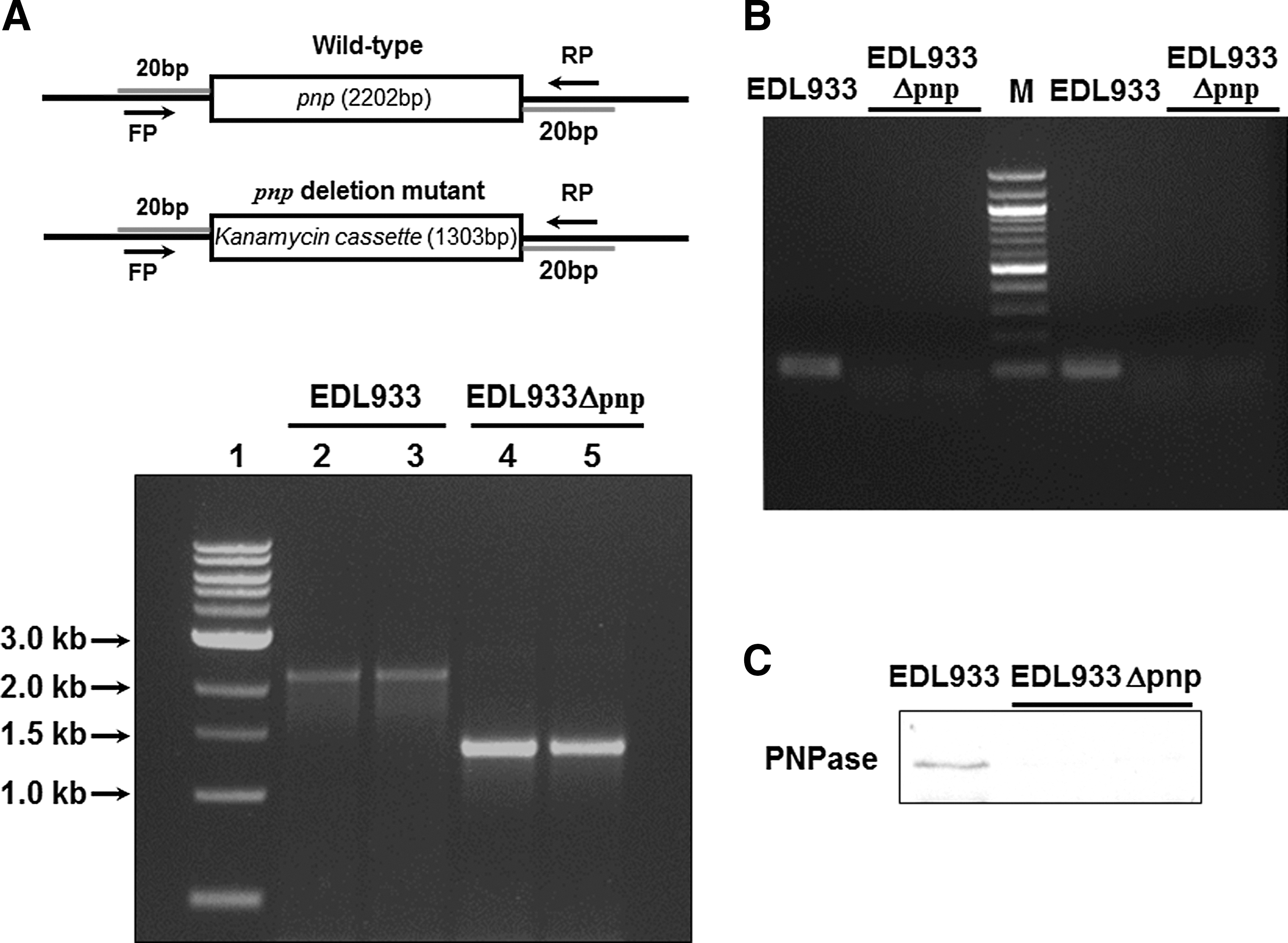
The confirmation of pnp mutant in Escherichia coli O157:H7 EDL933 strain.
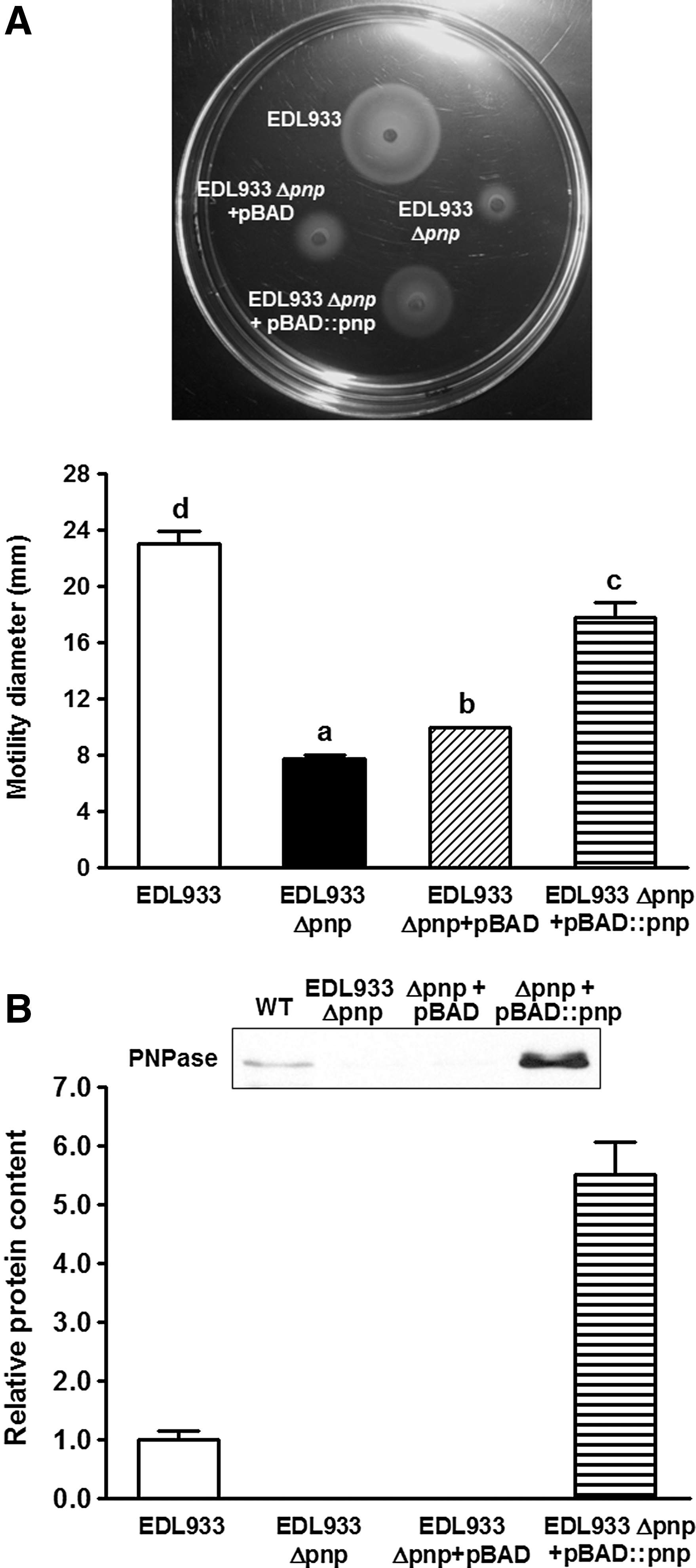
The swimming motility of Escherichia coli O157:H7 EDL933 and its derivative strains.
Growth at different temperatures in LB medium
The growth of wild-type and pnp deletion mutant strains was compared in both solid and liquid LB medium at different temperatures. The growth of EDL933Δpnp in LB and agar plates at 37°C was similar to that of EDL933 (Fig. 3A and C), but was impaired at 22°C (Fig. 3B). The growth of EDL933Δpnp at 22°C was partially restored by complementation with pBAD::pnp, but not the empty vector, pBAD (Fig. 3B). Similarly, EDL933Δpnp was unable to grow on LB agar plates at 10°C, which was again restored by overexpression of pBAD::pnp plasmid (Fig. 3). The deletion of pnp in E. coli O157:H7 strains sakai, 86-24, and MIO335 (Table 1) resulted in similar growth behaviors in LB medium (Fig. 4), further demonstrating a profound effect of PNPase in E. coli O157:H7 low-temperature growth (Fig. 4).
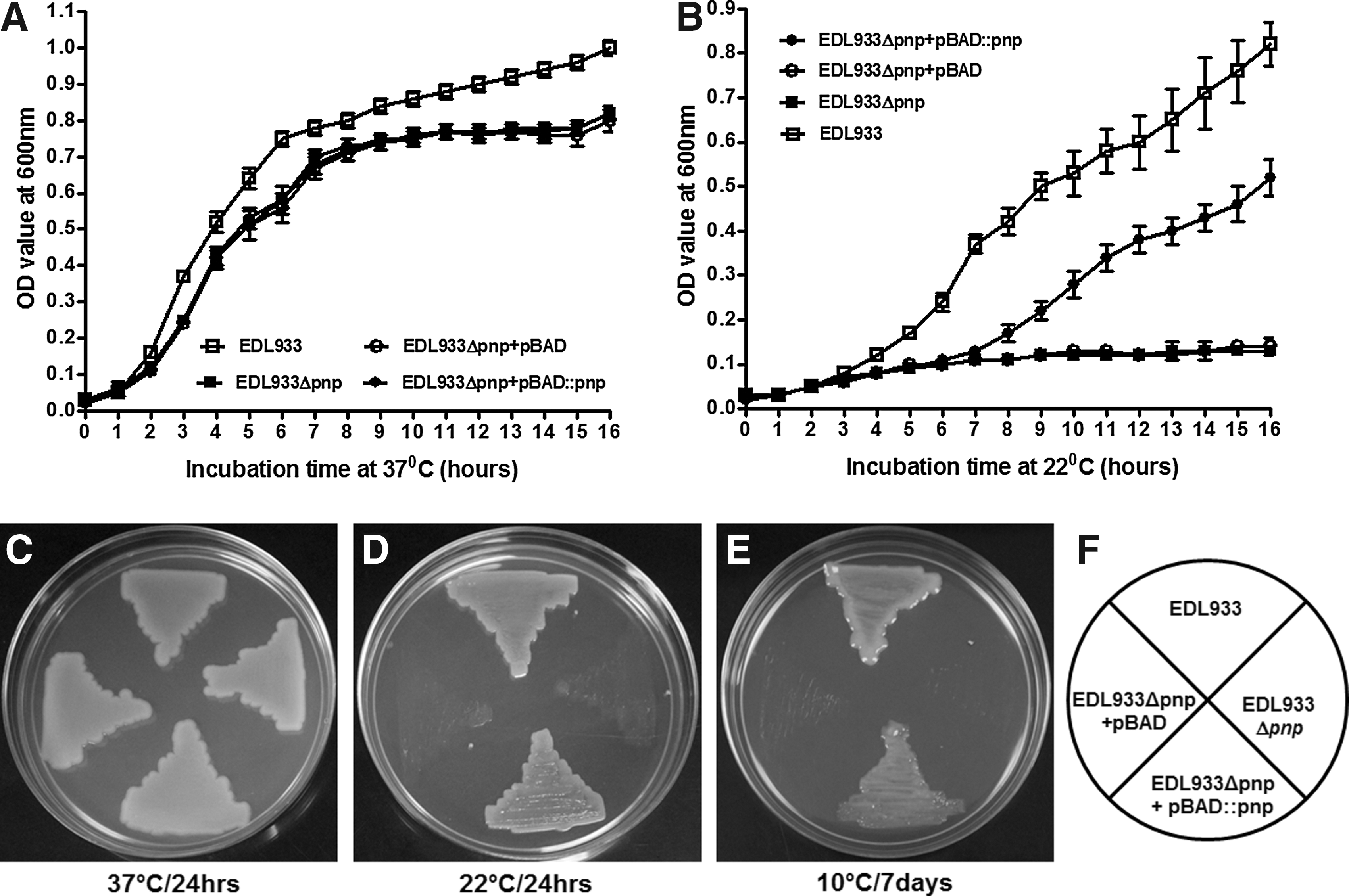
Growth of Escherichia coli O157:H7 in EDL933 and its derivative strains at different temperatures. Growth of E. coli O157:H7 strains in Luria Broth (LB) at 37°C
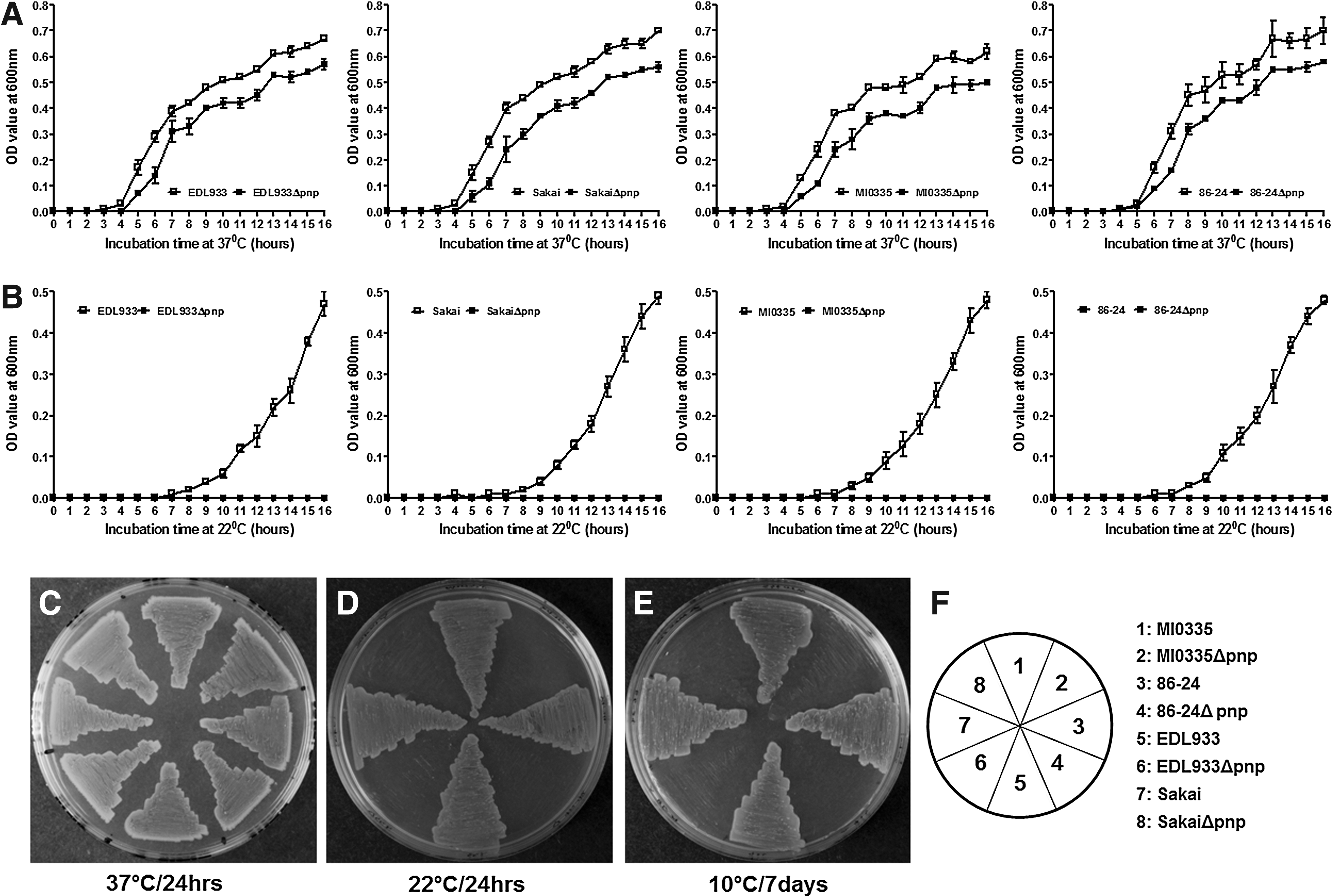
Growth of Escherichia coli O157:H7 strains Sakai, MI0335, and 86-24 and their corresponding pnp deletion mutant strains at different temperatures. Growth of E. coli O157:H7 strains in Luria Broth (LB) at 37°C
The growth and survival of EDL933 and EDL933Δpnp strains in LB broth were further studied during long-term storage at 4°C and 10°C. The EDL933 strain grew during 10°C storage and reached >8 log10CFU/mL after 4 days (Fig. 5A). No growth was observed in the EDL933Δpnp strain during 14 days of storage at 10°C (Fig. 5A), while complementation with pBAD::pnp restored the growth of EDL933Δpnp after 10 days at 10°C (Fig. 5A). Both EDL933and EDL933Δpnp did not grow in LB during 4°C storage; instead, both showed slight loss of viability (Fig. 5B), indicating that PNPase did not affect the survival of E. coli O157:H7 at 4°C.
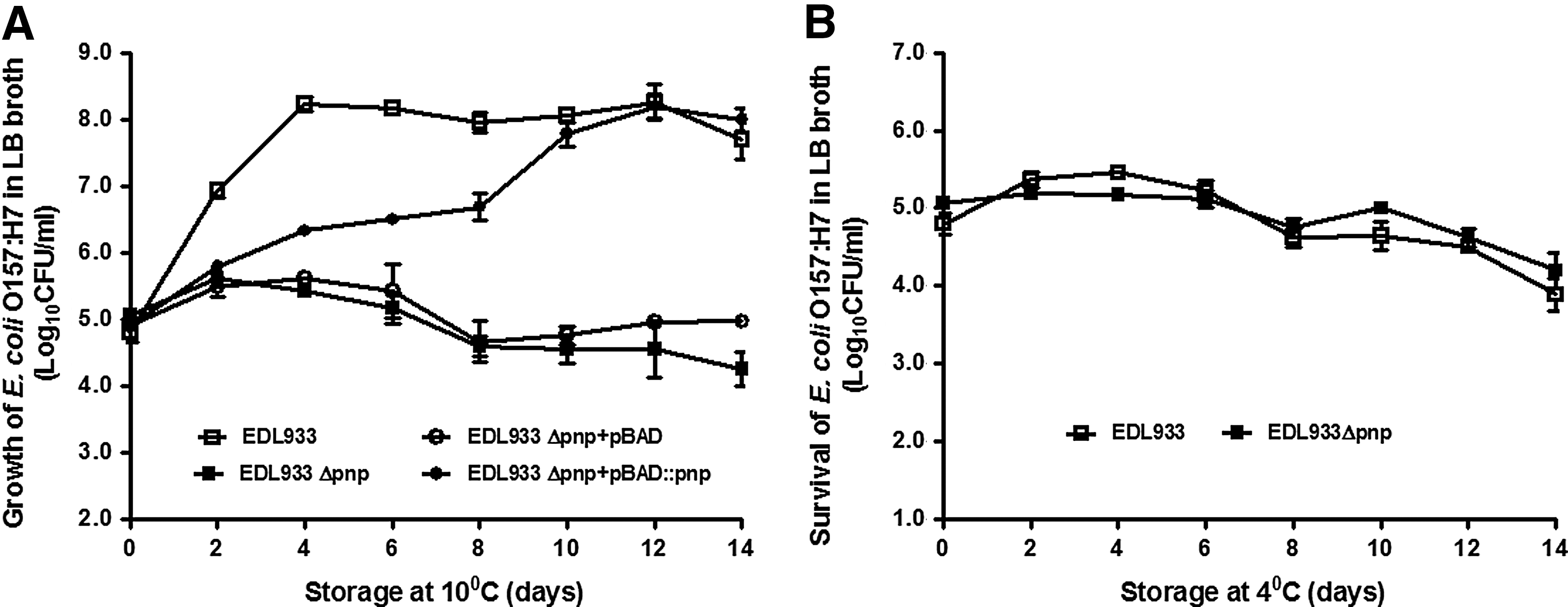
Growth and survival of Escherichia coli O157:H7 strains in Luria Broth (LB) during 14 days of storage.
Growth in milk at different temperatures
EDL933 grew well in milk when stored at 10°C, and increased more than 3 logs during 14 days of storage. On the other hand, there was more than 3-log reduction of the EDL933Δpnp in milk during 14 days of 10°C storage (Fig. 6A). As in LB, the growth of EDL933Δpnp with pBAD::pnp complementation was restored gradually during storage at 10°C. Both EDL933 and EDL933Δpnp were unable to grow but able to survive during 14 days of 4°C storage (Fig. 6B).
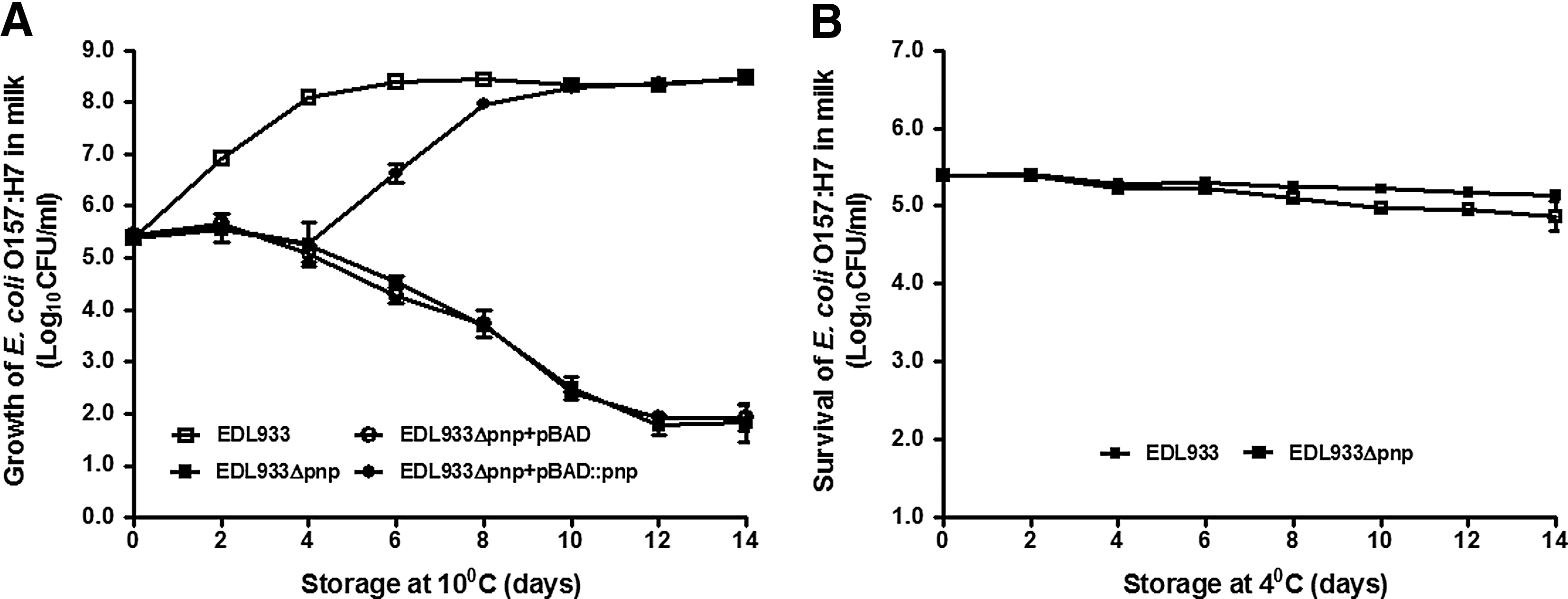
Growth and survival of Escherichia coli O157:H7 strains in commercial whole milk during 14 days of storage.
The susceptibility of EDL933 and EDL933Δpnp strains in response to different levels of H2O2 was further explored, and there was no difference in H2O2 susceptibility between EDL933 and EDL933Δpnp (Fig. 7).
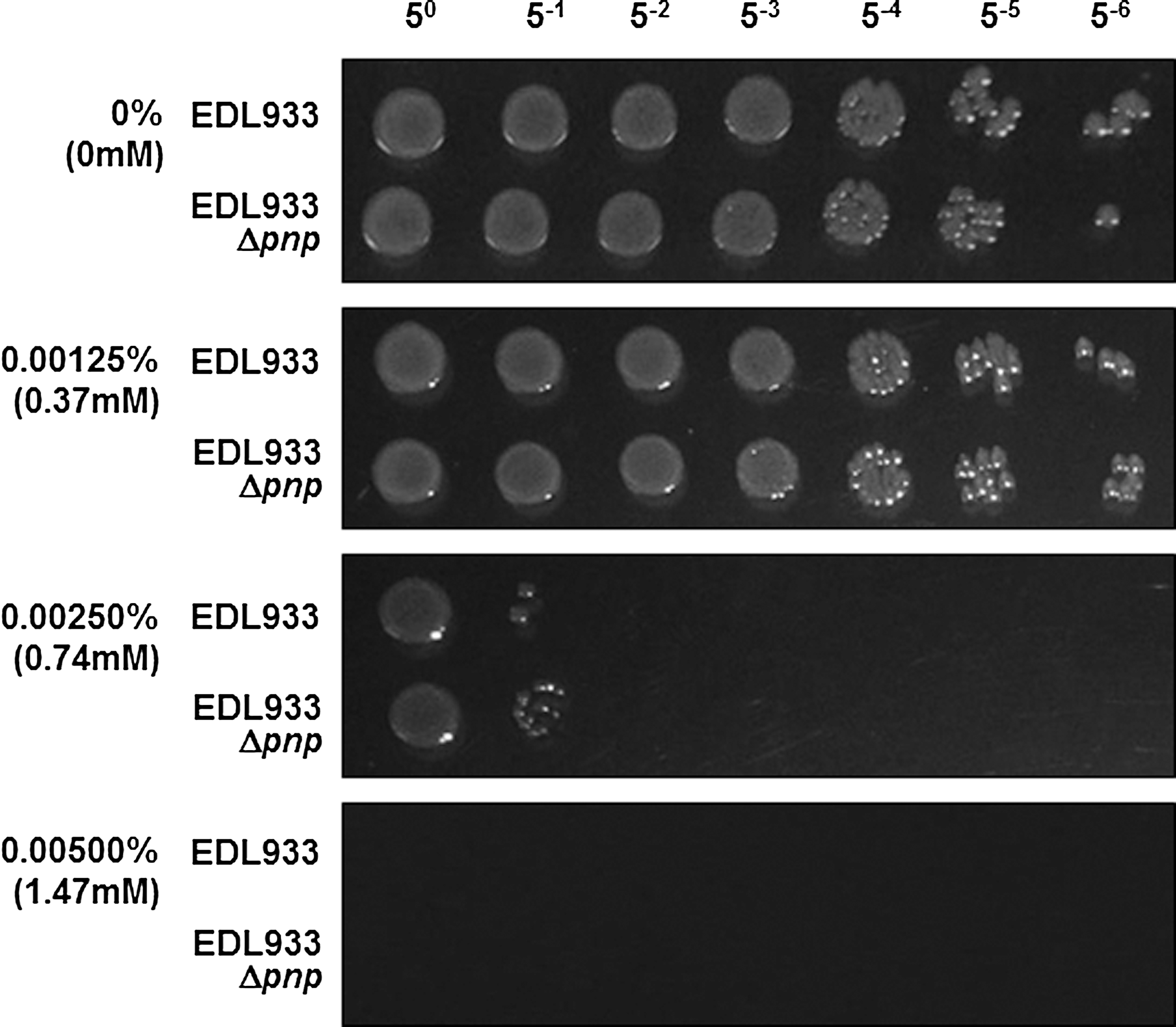
Cell viability of EDL933 and EDL933Δpnp on Luria Broth (LB) agar plates containing various concentrations of H2O2. Exponentially grown cultures were normalized to optical density (OD)550nm=0.04 and then serially diluted at 1:5, and then inoculated on plates.
Discussion
Milk is nutrient-rich and, thus, provides a favorable medium for the growth of many pathogenic bacteria. Major milkborne bacterial pathogens include Salmonella spp., Listeria monocytogenes, Staphylococcus aureus, Campylobacter spp., and pathogenic E. coli (Farrokh et al., 2013). Among them, E. coli O157:H7 is one of the major concerns to the dairy industry due to its low infectious dose and its ability to cause severe disease symptoms, which can grow significantly during handling or whenever there is an improper storage. Furthermore, vegetables and ground beef associated with E. coli O157:H7 outbreaks are often stored at low temperatures, where temperature fluctuation is common. Therefore, it is vitally important to control or prevent the growth of pathogens in milk and other foods at temperatures above refrigeration temperature. Many studies assessed the growth and survival of E. coli O157:H7 at low temperature (Wang et al., 1998; Elhanafi et al., 2004; Mutaku et al., 2005; Lu et al., 2011), but mechanisms regulating its growth at low temperature remain poorly understood. Previous studies have demonstrated that PNPase is required for cold adaptation of several examined pathogens and generic E. coli (Goverde et al., 1998; Haddad et al., 2009), through selectively degrading cold shock mRNAs at the end of the acclimation phase, further promoting the growth of generic E. coli (Yamanaka et al., 2001). However, the role of PNPase in the survival of generic E. coli or E. coli O157:H7 at a temperature relevant to refrigerated foods (4°C and 10°C) has not been evaluated.
During 14 days of storage, EDL933 and EDL933Δpnp strains behaved similarly at 4°C in both LB as well as milk, where they could not grow at 4°C but survived during 14 days of storage, indicating that PNPase is not required for the survival of E. coli O157:H7 at 4°C. This result differs from a study of Campylobacter jejuni in which it was shown that PNPase is indispensable for its long-term survival at both 4°C and 10°C (Haddad et al., 2009). On the other hand, our results demonstrate once again that the milk contaminated with E. coli O157:H7 remains hazardous during refrigerated storage, a finding in agreement with studies showing that E. coli O157:H7 survives in BHI broth (Palumbo et al., 1995), pasteurized and unpasteurized milk (Wang et al., 1997), or post-fermented African yogurt (Ogwaro et al., 2002) during 4°C–5°C storage. Similarly, PNPase had negligible effects on the growth of E. coli O157:H7 at 37°C in both LB and agar, which is consistent with studies on the generic E. coli K12 strain (Awano et al., 2008).
However, PNPase becomes crucial for E. coli O157:H7 growth when temperature decreases. After 14 days of 10°C storage, there was about 3.5 log10CFU and 6.5 log10CFU difference in LB and milk respectively, consistent with the study in Yersinia enterocolitica (Goverde et al., 1998), where deletion of PNPase resulted in defective growth at 5°C. In contrast, PNPase was not required for Salmonella enterica (Clements et al., 2002) and Pseudomonas putida (Favaro et al., 2003) growth at cold temperature, indicating that PNPase regulates cold temperature growth in a species-specific manner. Studies on the K12 strain indicate that PNPase mRNA increases in response to a temperature reduction from 37°C to 15°C, which involves post-transcriptional-level regulation (Zangrossi et al., 2000; Beran et al., 2001).
Because different E. coli O157:H7 strains exhibit considerable variation in stress resistance and growth abilities, we further tested the role of PNPase in the growth of different E. coli O157:H7 strains at cold temperature. Similar to EDL933, we found that PNPase is critical for the growth of Sakai, MI0335, and 86-24 at low temperatures.
Furthermore, we observed the decline of viable populations of EDL933Δpnp in milk during 10°C storage but not 4°C storage. We speculated that this was due to the antagonistic activity of milkborne microflora including lactic acid bacteria during storage (He et al., 2009). The pnp deletion impaired EDL933 growth at 10°C, thus making it less competitive compared with other strains. It is also possible that the deletion of pnp in E. coli O157:H7 may render it more sensitive to antimicrobials. Consistently, pnp disruption sensitizes the E. coli K-12 strain to antibiotics (McMurry et al., 1987). Therefore, the reduction of EDL933Δpnp populations during 10°C storage could also be due to the antimicrobial effect of natural antimicrobials in milk such as lactoferrin and lactoperoxidase (Al-Nabulsi et al., 2007), which are not destroyed by pasteurization (Paulsson et al., 1993) or to the small amount of antimicrobials produced by milkborne lactic acid bacteria during 10°C storage. In support of our results, a previous report demonstrated that the viability of the E. coli O157:H7 strain inoculated to pasteurized milk was reduced to undetectable levels within 4 days storage at 22°C (Wang et al., 1997).
In C. jejuni, PNPase positively regulated motility. In line with this, we found that the swimming motility of E. coli O157:H7 was significantly decreased in EDL933Δpnp. Motility is important for the initial attachment of bacteria. Thus, deletion of pnp might attenuate both growth at low temperature and E. coli O157:H7 adherence to the surface. Hydrogen peroxide is a powerful oxidant that is used to wash udders, milking facilities, and utensils. It is also allowed for use in raw milk for cheese production at 0.005% (vol/vol). Deletion of pnp did not affect the survival of EDL933 in response to different levels of H2O2, indicating that PNPase is probably not a major factor in the response of E. coli O157:H7 to oxidative stress.
Conclusions
In summary, pnp deletion dramatically impaired E. coli O157:H7 growth at 10°C and 22°C. Such impairment was largely recovered by ectopic expression of the pnp, demonstrating that growth impairment was PNPase-specific. In addition, pnp deletion impaired the motility of E. coli O157:H7 but did not affect its susceptibility to H2O2. Our finding that PNPase is necessary for the cold growth of E. coli O157:H7 at 10°C likely has important practical applications, since milk is routinely exposed to temperature fluctuations during production, processing, transportation, and handling in consumers' homes.
Footnotes
Acknowledgments
We thank Mr. Mark David Hanna for his technical support. This work was supported by USDA-AFRI 2010-65201-20599, the Agricultural Experiment Station at University of Wyoming, and Agricultural Research Center at Washington State University.
Disclosure Statement
No competing financial interests exist.