
Abstract
Norovirus infection is a major cause of nonbacterial foodborne outbreaks worldwide, but no specific treatments are available yet. In this study, we investigated the antiviral activity of mulberry (Morus alba, Ma) juice and its fractions on murine norovirus-1 (MNV-1) and feline calicivirus-F9 (FCV-F9) as human norovirus surrogates using cytopathic effect inhibition, plaque reduction, and RNA expression assays. In time-of-addition experiments, Ma juice was found to be effective in reducing the infectivity of MNV-1 and FCV-F9 in the pre- and co-treatments. The effective concentration for 50% reduction was approximately 0.005% juice (relative to 100% natural juice) and 0.25% juice for MNV-1 and FCV-F9, respectively. Ma juice at 0.1% exhibited about 60% reduction of the MNV-1 polymerase gene expression, confirming the inhibition of viral replication. In an attempt to identify active components with antiviral activities, Ma-F1 (<3 kDa) and Ma-F2 (>3 kDa) were examined to show that Ma-F2 was more effective than Ma-F1 in all modes, except for pre-virus treatment. Nevertheless, two major polyphenolic compounds of Ma juice, cyanidin-3-glucoside and cyanidin-3-rutinoside, showed antiviral activity in the co-treatment mode. Our results suggest that Ma juice and its fractions may inhibit internalization and replication of MNV-1, whereas it may influence adherence or internalization of FCV-F9 virions. Ma juice may prove useful in the prevention of foodborne viral infection.
Introduction
H
Little is known about the biology of HuNoVs, which is mainly due to the absence of a cell culture system or animal model (Rocha-Pereira et al., 2010). Murine norovirus (MNV-1) and feline calicivirus (FCV) were successfully grown and have served as a surrogate model system for HuNoVs (Wobus et al., 2004, 2006). Despite the significant economic loss and threat to public health worldwide, there is no vaccine or antiviral drug available for prevention or treatment of HuNoVs infection.
It has been reported that natural food products and polyphenolic compounds from plants effectively inhibited the viral infection, including NoV. Cranberry juice and its proanthocyanidin (PAC) reduced the infectivity of MNV-1 and FCV-F9, showing virucidal effects (Su et al., 2010a, 2010b). Pomegranate juice and polyphenol, grape seed extract, and chitosan were reported to be effective against NoV surrogates (Su and D'Souza, 2011). In our previous report, black raspberry fruit juice was shown to effectively inhibit the MNV-1 when it was treated simultaneously with MNV-1 virus (Oh et al., 2012).
Mulberry (Morus alba L.) belongs to the Moraceae family and its leaves, barks, and branches have been used in Chinese medicine (Arfan et al., 2012). It is rich in flavonoids, including quercetin, rutin, and cyanidin-3-rutinoside and exhibits anti-oxidative, antihypotensive, and antiviral activities (Du et al., 2003). It was shown to possess anti-adherence activity against Streptococcus mutans biofilm (Islam et al., 2008) and anti-microbial activity against methicillin-resistant Staphylococcus aureus, Bacillus subtilis, and Escherichia coli (Fukai et al., 2005). In the present study, we examined antiviral effects of mulberry (Ma) juice and its fractions against MNV-1 and FCV-F9 to develop the antiviral agents for prevention or control of HuNoV infection.
Materials and Methods
Viruses and cell lines
FCV-F9 (ATCC VR-2047) and Crandell Reese Feline Kidney (CRFK, ATCC CCL-94) cells were obtained from ATCC (Manassas, VA). RAW 264.7 cells and MNV-1 were kindly provided by Dr. Jai Myung Yang at Sogang University, Korea. CRFK and RAW 264.7 cells were maintained in Dulbecco's Modified Eagle Medium (DMEM) (Invitrogen, Grand Island, NY) supplemented with 10% heat-inactivated fetal bovine serum (FBS) (Sigma-Aldrich, St. Louis, MO) and 1x penicillin streptomycin (PS) (Invitrogen, Grand Island, NY) at 37°C in an atmosphere containing 5% CO2. MNV-1 and FCV-F9 were propagated in RAW 264.7 and CRFK cells with DMEM-2% FBS-1% PS medium for 3–4 days, with the titers at 4×105 plaque-forming units (PFU)/mL and 4×106 PFU/mL, respectively, and stored at −80°C.
Preparation of Ma juice and phenolic compound and determination of total polyphenols
Cyanidin-3-glucoside (C-3-G), cyanidin-3-rutinoside (C-3-R), gallic acid, quercetin-3-glucoside (Q-3-G), quercetin, and caffeic acid were purchased from Sigma-Aldrich (St. Louis, MO). Ma fruits (Sununsan Nonghyup, Korea) were purchased from a local market and identified by staff members of Natural Academy of Agricultural Science, Rural Development Administration, Korea. One hundred percent juice was prepared by 0.22 μm sterilized filtration after enzyme inactivation at 63°C of squeezed juice as described previously (Oh et al., 2012). It was further fractionated using an Amicon® Centriprep® filter (Millipore, MA) with molecular weight cutoff of 3 kDa into Ma-F1 and Ma-F2 containing components less than and greater than 3 kDa, respectively, which were lyophilized. Total polyphenol contents of juice were determined by the Folin-Ciocalteu method at 750 nm using microplate reader (SpectraMax M2, Molecular Devices Corp.), using gallic acid as a standard (Oh et al., 2012).
Cytotoxicity of Ma juice on CRFK and RAW 264.7 cell lines
Cytotoxicity was measured by an MTT (3-[4,5-dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide) assay as described previously (Oh et al., 2012). Briefly, after 12–24 h of cell seeding, 90 μL of DMEM-10% FBS-1% PS and 10 μL of 0.001%–1% (relative to 100%) of Ma juice were added to the cells, which were further incubated for 1–48 h or 72 h at 37°C. The percentage of cell viability after treatment with the juice was calculated as follows: % cell viability=[Abstreatment/Abscontrol]×100.
Cytopathic effect (CPE) reduction
RAW 264.7 or CRFK cells were seeded in 96-well plates (2×105 cells or 0.5×104 cells per well, respectively) for 12 h. After removing of medium, 10 μL of Ma juice or sterilized distilled water (control) and 10 μL of virus were mixed in cells containing 80 μL of media for 48–72 h. CPE by virus infection was measured using MTT assay as described above, and CPE reduction (%) was calculated as (Abs – Absvirus)/(Absnoninfected cell – Absvirus)×100. Selectivity index (SI) was calculated as the ratio of 50% cytotoxic concentration (CC50) to 50% effective concentration (EC50). All experiments were performed in triplicate.
Plaque reduction assay
To understand the inhibitory mechanisms of Ma juice and samples on NoVs surrogate infections, “pretreatment,” “co-treatment,” and “post-treatment” conditions were analyzed as time-of-addition plaque reduction assay as described previously (Oh et al., 2012). Briefly, sample was pretreated with cell or virus before viral infection (pretreatment of cells or previrus treatment, respectively) or co-treated during viral absorption for 1 h (co-treatment) or post-treated after virus adsorption for 1 h (post-treatment). Five and/or 2∼3 log10 PFU/mL virus was used for previrus or/and other treatments, respectively (final 50–100 PFU/well of MNV-1 and FCV-F9). Sterilized distilled water and 2-thiouridine (2TU) were used as negative and positive controls, respectively (Alam et al., 2012). Antiviral activities were also calculated as relative plaque formation % for control group based on different PFU and titer reduction (log10 PFU/mL) from recovered viruses (Oh et al., 2012).
Reverse transcriptase-polymerase chain reaction (RT-PCR) analysis
To examine the ability of Ma juice to prevent the replication of virus, the gene level of MNV-1 RNA-dependent RNA polymerase (RdRp) was measured by Applied Biosystems StepOne™ Real-Time PCR System and PCR. Juice was simultaneously or post-treated with MNV-1 inoculation (multiplicity of infection [MOI]=0.01) onto RAW 264.7 cells for 1 h. After removing the juice, the cells were incubated with fresh media. After 24 h, viral RNA was extracted from cells by RNeasy kit (QIAGEN). After cDNA synthesis using high-capacity cDNA reverse transcription kits (Applied Biosystems) as a recommended protocol, PCR or real-time PCR was performed with methods as described previously (Alam et al., 2012) or by manufacturer's protocol using Power SYBR green PCR Master Mix (Applied Biosystems). RT cDNA (12.5 μL) was added to Master Mix (12.5 μL) and amplified using the following thermal cycles: 50°Cx2m, 95°Cx10m, and [95°Cx15s, 60°Cx1m] 35 cycles. ΔCt and ΔΔCt were calculated as an average of duplicates in two independent experiments. RdRp mRNA expression levels were represented as a relative quantitation (RQ=2-ΔΔCT) to control.
Differential scanning fluorimetry (DSF)
In order to check the binding of Ma juice to MNV-1 RdRp protein, we used the DSF method, which measures the shift in melting temperature (Tm) upon ligand binding (Niesen et al., 2007). RdRp at a concentration of 0.5 mg/mL (8.25 μM) was incubated with a juice or each compound and mixed with 0.5 μL (500-fold) SYPRO Orange (Invitrogen, Grand Island, NY). The thermostability (Tm) value of RdRp was measured as described previously (Alam et al., 2012) using Mx3005P DSF (Stratagene, CA).
Statistical analysis
The experimental data were statistically analyzed using analysis of variance with SPSS software (version 9.2, SPSS Institute, Cary, NC), and the means were evaluated with Duncan's multiple-range test. Statistics with a value of p<0.05 were considered significant. Experimental results are expressed as mean±standard deviation. All measurements were performed in triplicate.
Results
Antiviral activity of Ma juice against MNV-1 and FCV-F9 infection
Ma juice at 0.1% reduced approximately 40% of RAW 264.7 cell viability for 3 days (data not shown). In contrast, up to 2.5% of the juice did not affect the CRFK cell viability significantly. Antiviral assays including CPE and plaque assays were performed within 50% cytotoxic concentration (CC50) of Ma juice, which was less than 0.1% for RAW 264.7 cells and 2.5% for CRFK cells (Table 1).
The effective dose for reducing 50% (EC50) was measured in cytopathic effect (CPE) and plaque reduction assay (PA). EC50 of PA was calculated approximate average value.
The CPE and 50% cytotoxic concentration (CC50) was measured by MTT assay.
The selective index (SI) was calculated as CC50/EC50.
MNV-1, murine norovirus-1; FCV-F9, feline calicivirus-F9.
When antiviral activity of Ma juice was measured in virus-infected cells, 0.004% Ma juice inhibited 60% of the CPE induced by MNV-1 and 0.01% juice showed 100% complete inhibition (data not shown). The activity of Ma juice was also effective in FCV-infected CRFK cells, showing 40% inhibition at concentrations of 0.3%–0.6%. Ma juice at concentrations of 0.005% and 0.25% reduced the plaque formation of MNV-1 and FCV-F9, respectively, to above 50% effectively. Selectivity indexes (SI=CC50/EC50) were 20 and 8.3–10 for MNV-1 and FCV-F9 by CPE and plaque assay, respectively (Table 1).
In the precell treatment, 0.1% Ma juice (Fig. 1A) reduced the plaque formation to 29.4%, and serially diluted concentrations also inhibited the plaque formation (Fig. 1A). In the previrus treatment, 0.1% Ma juice reduced the plaque formation to 38.9%. It was also reduced to 35.5% and 29.1% in the co- and post-treatment by 0.1% juice, respectively. Similar results were observed for the other tested concentrations at 0.001%–0.1% Ma juice, showing broad inhibition effects in various time-of-addition modes.
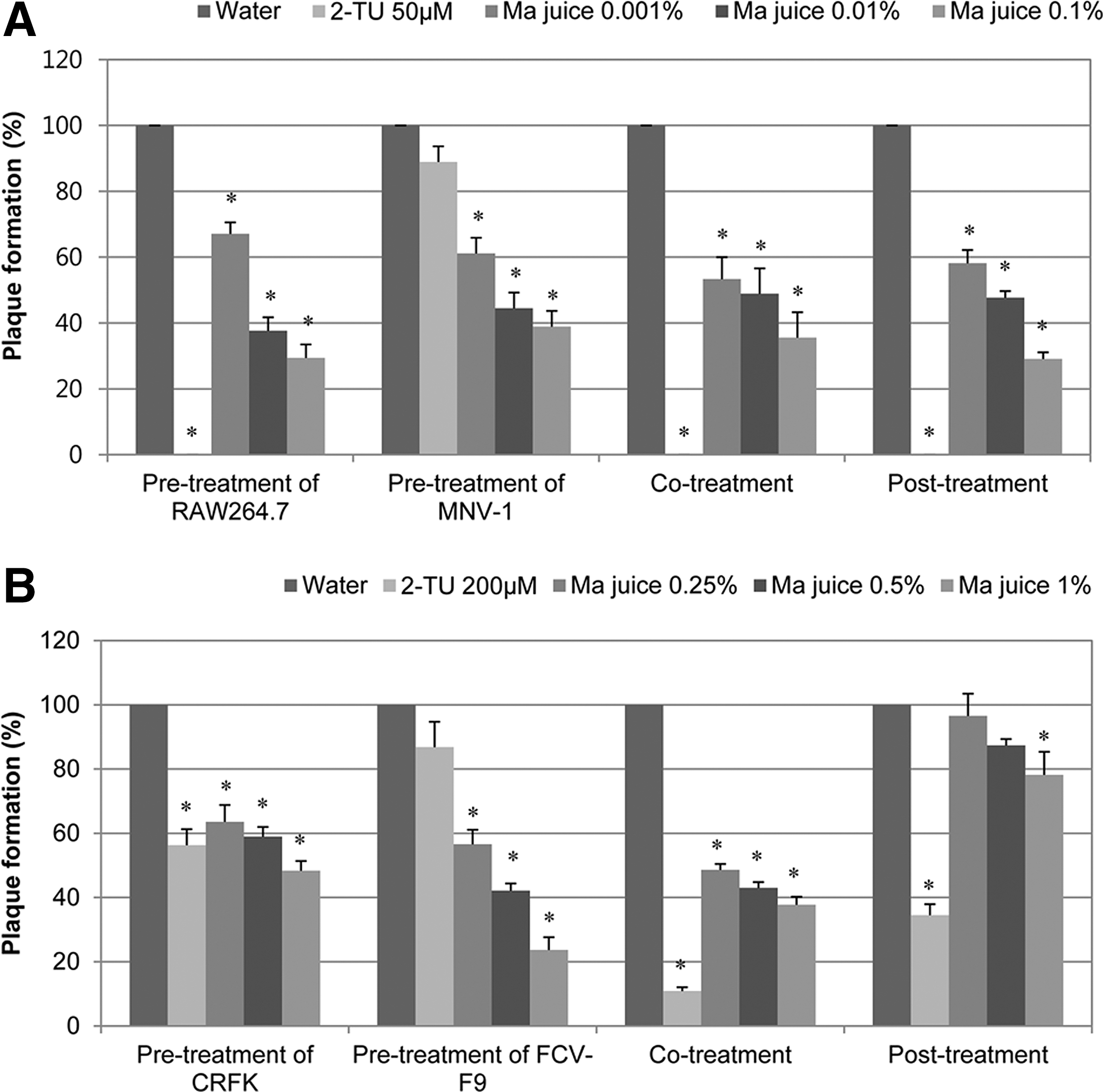
Anti-murine norovirus (MNV-1) or anti-feline calicivirus (FCV-F9) effect of Ma juice by plaque reduction assay in time-of-addition modes.
In the case of FCV, 1% Ma juice exhibited plaque reduction to 23.7% and 37.8% in the pretreatment and co-treatment of viruses, respectively (Fig. 1B), whereas plaque reduction was observed to be mild in the post-treatment. When we compared the inhibitory activity with 2TU used as a positive control, 0.1% Ma juice had similar activity to that of 1∼10 μM 2TU (data not shown).
Influence of Ma juice on MNV-1 RNA expression
In the co-treated cells, 0.1% Ma juice treatment led to significant reduction of the MNV-1 RdRp gene, showing much lower gene level than that of 20 μM 2TU (Fig. 2A). This result suggests that viral internalization was inhibited by Ma juice, resulting in reduction of RNA expression. Ma juice (0.1%) in the post-treatment exhibited about 60% reduction of the MNV-1 RdRp gene expression, showing comparable activities to that of 20 μM 2TU. Ma juice (1%) abolished the mRNA expression almost completely (Fig. 2B). Agarose gel electrophoresis also showed decreased mRNA expression levels (Fig. 2C). However, Ma juice and its major polyphenolic components did not alter the Tm value of MNV-1 polymerase, RdRp, suggesting that they do not directly bind to the MNV-1 viral enzyme (data not shown).
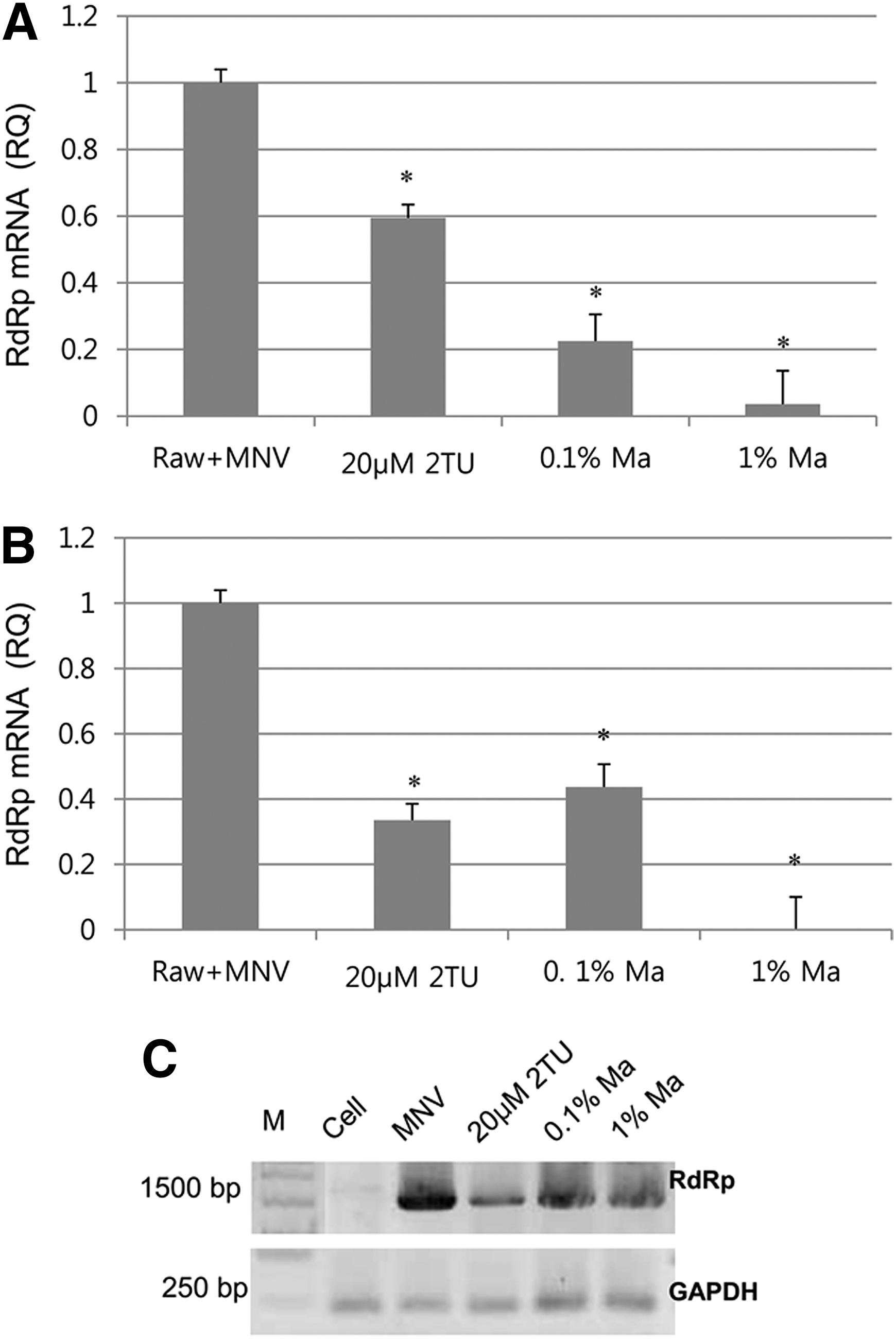
Influence of Ma juice on RNA expression analyzed by real-time quantitative reverse transcriptase polymerase chain reaction (qRT-PCR) and conventional PCR. Murine norovirus (MNV)-1 infection in RAW 264.7 cells was treated with Ma juice during (
Antiviral activities of Ma juice fractions Ma-F1 and Ma-F2
In an attempt to identify active components with antiviral activities from the juice, Ma juice was further purified into two factions by molecular size, Ma-F1 (<3 kDa) and Ma-F2 (>3 kDa). In the pre-MNV-1 treatment, Ma-F1 and Ma-F2 reduced the plaque formation to 41.5% and 66.0% at a concentration of 2 mg/mL, respectively (Figs. 3A and B). In terms of the high-molecular-weight fraction, Ma-F2 was more effective than Ma-F1, showing 4%–17% plaque formation in all the treatments in a dose-dependent manner (Fig. 3B). When it was calculated as recovered viral titer, MNV-1 titer was reduced by 1.19 log10 PFU/mL with 2 mg/mL of Ma-F2 in the co-treatment, followed by pre-cell (1.05 log10 PFU/mL) and post-treatment (0.79 log10 PFU/mL) (Supplementary Table S1; Supplementary Data are available online at
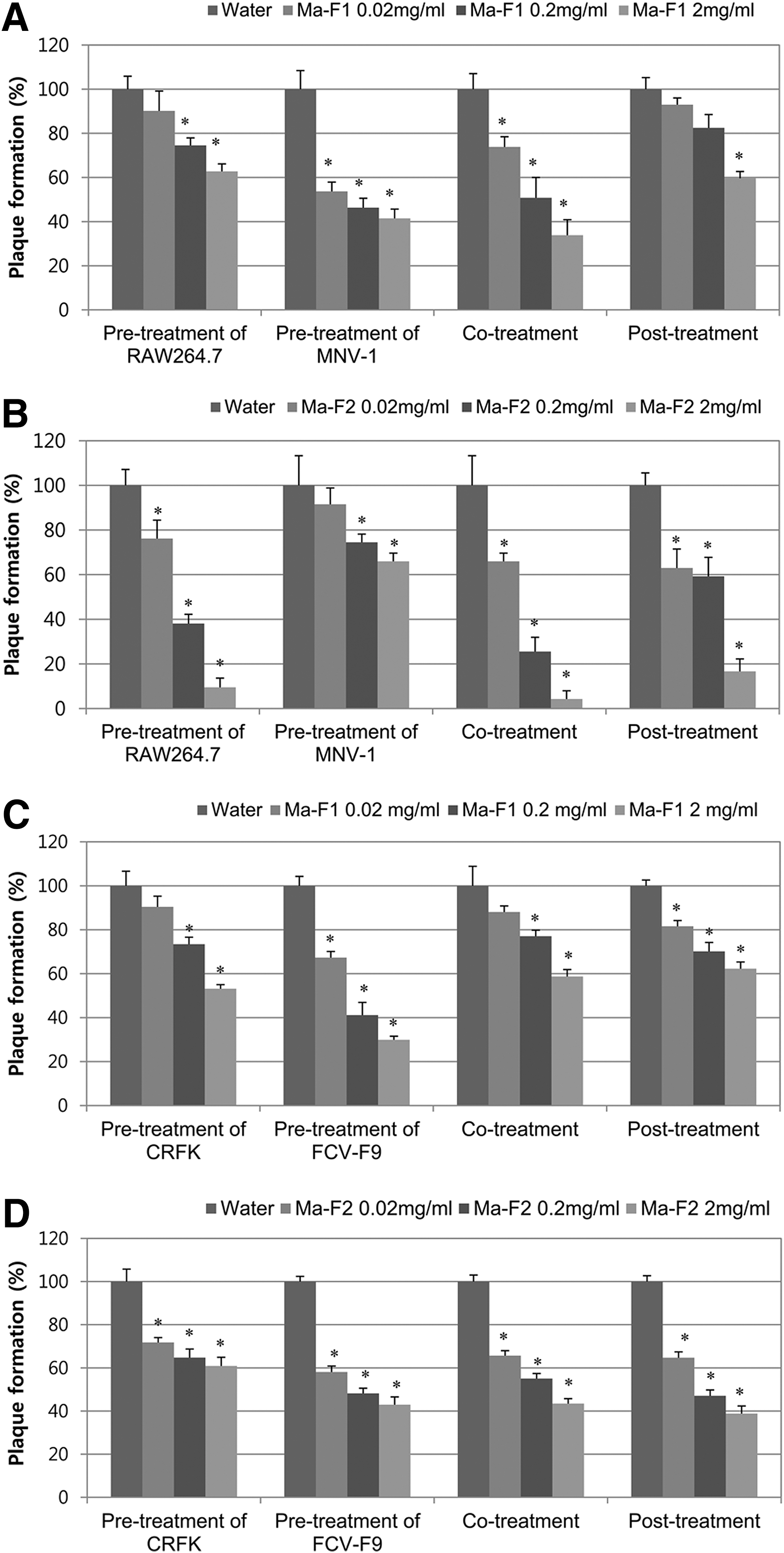
The antiviral effect of Ma fractions against murine norovirus (MNV)-1 and feline calicivirus-F9 (FCV-F9) by plaque reduction assays.
Effects of cyanidin-3-glucoside and cyanidin-3-rutinoside
The total polyphenolic content of Ma juice (100%) was found to be 2.54 mg gallic acid equivalent (GAE)/mL by Folin-Ciocalteu method (Oh et al., 2012). Ma-F1 and Ma-F2 contained 0.02 mg and 0.01 mg GAE per mg, respectively.
Mulberry contains high amounts of flavonoids such as anthocyanins. Major phenolic components of Ma juice, cyanidin-3-glucoside and cyanidin-3-rutinoside, were used to examine the antiviral activity in the co-treatment mode. Cyanidin-3-glucoside at 100 μM exhibited 40% plaque formation against MNV-1 (Fig. 4A) and FCV-F9 (Fig. 4B) and 100 μM cyanidin-3-rutinoside showed 60% and 40% plaque formation against MNV-1 and FCV-F9, respectively. EC50 of these single compounds against MNV-1 in the co-treatment was 100 μM.
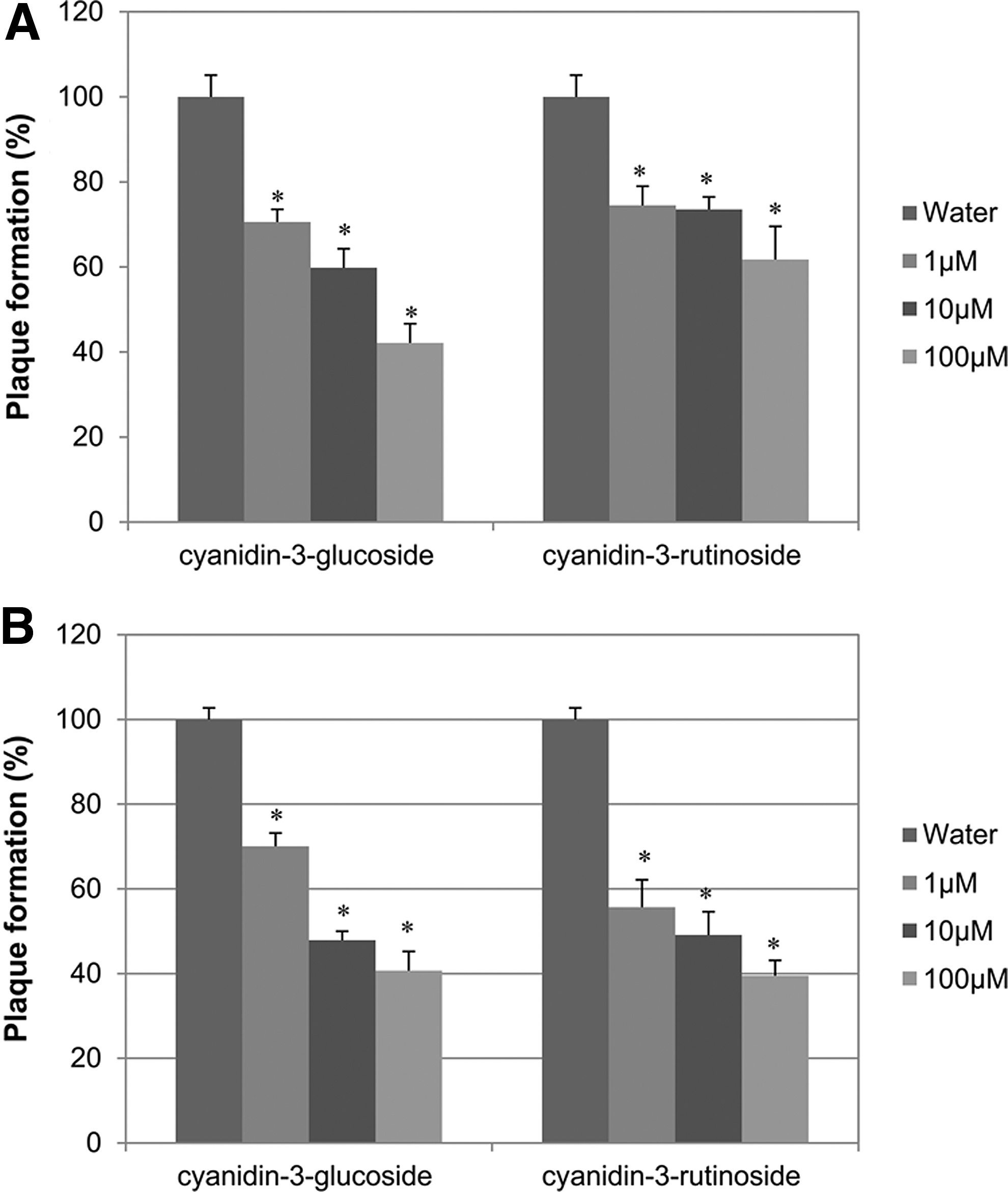
The antiviral effects of cyanidin-3-glucoside and cyanidin-3-rutinoside against
Discussion
The antiviral effects of cranberry juice, grape juice, and pomegranate juice on the infectivity of enteric viruses were previously reported (Su et al., 2010a, 2010b , 2010c; Su and D'Souza, 2011). Black raspberry juice was also shown to inhibit MNV-1 infection very effectively (Oh et al., 2012). In the present study, just 0.0001%–0.1% Ma juice significantly reduced the plaque formation of MNV-1, while cranberry juice and others were used at final 50% (Su et al., 2010a, 2010b, 2010c; Su and D'Souza, 2011). Given that HuNoV is highly stable and contagious, and less than 20 virions can cause illness, it is notable that the effective concentration of Ma juice is lower than that of other juice or extracts. Furthermore, Ma-F2 was also effective in the pretreatment, co-treatment (1.2 log10 PFU/mL reduction), and post-treatment, while Ma-F1 were slightly more effective in the previrus treatment. This suggests that high-molecular-weight (>3 kDa) components of Ma juice may play a role in inhibiting the binding of MNV-1 by blocking of the host cell receptors and internalization into cells as well as viral replication. Our results also demonstrate that Ma juice showed not only marked CPE and plaque reduction of MNV-1 but also down-modulation of RdRp mRNA expression. However, it did not cause any direct influence on viral polymerase protein itself.
Polyphenolic contents analysis shows that Ma juice has higher polyphenolic contents (2.54 mg GAE/mL) than that of black raspberry fruit (0.64 mg GAE/mL) (Oh et al., 2012) or cranberry juice (1.04 mg GAE /mL), which were known to be responsible for antiviral activity (Su et al., 2010a). Various actions of polyphenolics derived from plants have been proposed, including inhibiting the adsorption of the virus to the cell receptor (Haslam, 1996). PAC of cranberry or grape juice and seed extract indicated antiviral activity against human enteric virus surrogates at 0.3–0.6 mg/mL levels, showing virucidal effects ( Su et al., 2010b, 2010c; Su and D'Souza, 2011). PAC induced the structural change or damage of FCV-F9 (Su et al., 2010a), and its cocktails changed the structure of rotavirus SA-11 virus particles, including alteration of viral receptor sites (Lipson et al., 2007). Cranberry and pomegranate juices reduced the specific binding of HuNoV P particles (Li et al., 2012). Resveratrol from Geranium sanguineum, catechins from green tea, and polyphenols such as CYSTUS052 from Cistus incanus appear to act with anti-influenza viral action by blocking viral attachment and penetration (Ludwig, 2011). In this study, low-molecular-weight anthocyanins, cyanidin-3-glucoside and cyanidin-3-rutinoside, as well as gallic acid or quercetin (Oh et al., 2012) showed antiviral activity in the co-treatment (20%–30% activities at 1 μM), and Ma-F2 was most effective in the co-treatment (4% plaque formation at 2 mg/mL). Our results suggest that polyphenolic compounds of Ma juice may influence the inhibition of norovirus infection at the attachment or internalization of virions and at the early stage of replication. Further examination is necessary to confirm the active components and explain their antiviral mechanisms.
Virus attachment is mediated by MNV-1 capsid proteins via receptors such as ganglioside GD1a (Perry and Wobus, 2010). MNV-1 endocytosis could be significantly inhibited by nystatin, methyl-β-cyclodextrin, and the dynamin-specific inhibitor dynasore (Perry and Wobus, 2010). Small-molecule therapeutics for norovirus infection such as piperazine, cyclosulfamide, (E)-2-styrylchromones derivatives (Rocha-Pereira et al., 2010), favipiravir (Rocha-Pereira et al., 2012), ribavirin, and 2-TU also have been recently reported (Rocha-Pereira et al., 2010; Alam et al., 2012; Dou et al., 2012;). However, natural food products might be safe and ecofriendly without side-effects.
In conclusion, Ma juice and its components showed antiviral effects against foodborne enteric virus surrogates. It was found to be effective in reducing the infectivity of MNV-1 in various treatment modes including pre- and co-treatment by plaque reduction as well as reduction of CPE and MNV-1 RdRp gene expression. Ma juices and its components might be good adjuncts for prevention of NoV infection and disease. To maximize the antiviral effect, combination with other active components or treatments will be considered in a future study.
Footnotes
Acknowledgments
This work was supported by the National Research Foundation of Korea Grant funded by the Korean Government (Ministry of Education, Science and Technology) (NRF-2011-355-C00119).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.