
Abstract
Three strains of Vibrio parahaemolyticus (690, BCRC 13023, and BCRC 13025), involved in foodborne outbreaks in Taiwan, were subjected to acid adaptation at pH 5.5 for 90 min. The effects of acid adaptation on the tolerance of V. parahaemolyticus to various environmental stresses, including heat (47°C), cold (4°C and −20°C), ethanol (8%), high salt (20% NaCl), and hydrogen peroxide (20 ppm) were examined. Results showed that acid adaptation increased the thermal tolerance of the three test strains of V. parahaemolyticus, while it did not affect their cold tolerance. Acid adaptation also increased the ethanol tolerance in V. parahaemolyticus 690 and BCRC 13025, but not in BCRC 13023. Differences in the tolerance to high salts were noted among the three strains after prior acid adaptation. However, these acid-adapted V. parahaemolyticus strains were more susceptible to hydrogen peroxide than their nonadapted controls. These findings demonstrated that acid adaption responses of V. parahaemolyticus varied among strains and types of stress challenge.
Introduction
A
Vibrio parahaemolyticus is a Gram-negative and moderately halophilic bacterium that causes human gastroenteritis associated with seafood consumption. In addition to contamination within seafood, cross-contamination of other food products can occur through the use of contaminated seafood-processing utensils. It has been recognized as a major cause of foodborne illness in many coastal countries, including China, India, Japan, Korea, Malaysia, Taiwan, Thailand, and the United States (Wong et al., 1999; DePaola et al., 2003; Nair et al., 2007; Su and Liu, 2007). In Taiwan, V. parahaemolyticus accounted for 61.1% of bacterial foodborne outbreaks between 1981 and 2012 (Taiwan Food and Drug Administration, 2013). In addition, the Centers for Disease Control and Prevention (CDC) also indicated an 85% increase in the incidence of Vibrio infections in 2009 compared with 1996–1998 baseline and calculated that 4500 cases of V. parahaemolyticus infection occur each year in the United States (CDC, 2010, 2013).
The tolerance of V. parahaemolyticus to various stresses is an important factor for its survival and persistence in seafood and in food-processing systems. Several studies have reported that stress adaptation induced cross-protection against environmental stresses in V. parahaemolyticus, V. cholerae, and V. vulnificus (Koga and Takumi, 1995a, b; Koga et al., 1999; Wong et al., 1998, 2002; Alvarez et al., 2003; Chang et al., 2004; Yeung and Boor, 2004; Bang and Drake, 2005; Chiang et al., 2006; Koo and Jahncke, 2006; Chiang and Chou, 2009; Huang and Wong, 2012).
Our previous studies revealed that the induced acid-adapted acid tolerance response in V. parahaemolyticus can be affected by acid-adaptation conditions, growth phase of the cells, and varies among bacterial strains. Among the test strains, V. parahaemolyticus 690, BCRC 13023, and BCRC 13025, involved in foodborne outbreaks in Taiwan, in late-exponential phase exhibited a greater degree of acid tolerance after acid adaptation at pH 5.5 for 90 min (Chiang et al., 2012b). Furthermore, two-dimensional electrophoresis analysis showed that protein expression also differed between acid-adapted and nonadapted strains of V. parahaemolyticus (Chiang et al., 2012a). These acid-adaptive proteins have been consistently reported to be involved in stress response (Jyot and Ghosh, 1995; Flahaut et al., 1997; Merrell et al., 2001; Wang et al., 2003), pH homeostasis (Hamilton and Buckley, 1991; Bearson et al., 1997; van de Guchte et al., 2002; Sánchez et al., 2007), energy and carbohydrate metabolism (Welin et al., 2003; Fernandez et al., 2008; Lee et al., 2008; Koponen et al., 2012), and may play important regulatory roles in acid or other stress-tolerance responses among V. parahaemolyticus strains. In this study, we further examined the effect of acid adaptation on the tolerance of three strains of V. parahaemolyticus to various environmental stresses.
Materials and Methods
Microorganisms
V. parahaemolyticus strains 690, BCRC 13023, and BCRC 13025, which were involved in a number of foodborne outbreaks in Taiwan, were used as test strains. V. parahaemolyticus 690 was obtained from Department of Microbiology, Soochow University (Taipei, Taiwan). V. parahaemolyticus BCRC 13023 and BCRC 13025 were purchased from Bioresource Collection and Research Center (BCRC), Food Industry Research and Development Institute (Hsinchu, Taiwan). These three strains of V. parahaemolyticus were subcultured twice for 6 h and then incubated for 4 h at 37°C in tryptic soy broth supplemented with 3% NaCl (TSB-3% NaCl) to reach the late-exponential phase before each experiment.
Acid adaptation treatment
To perform the acid-adaptation treatment, the late-exponential phase cultures (50 mL) of V. parahaemolyticus were centrifuged (3000×g, 10 min) and washed twice with phosphate-buffered saline containing 3% NaCl (PBS-3% NaCl, 0.1 M Na2HPO4/NaH2PO4, pH 7). Acid-adapted V. parahaemolyticus was prepared by suspending in 50 mL of TSB-3% NaCl acidified to pH 5.5 with 10 N HCl and held at 37°C for 90 min. Nonadapted V. parahaemolyticus was taken by suspending in the same volume of fresh TSB-3% NaCl without acidification. Both acid-adapted and nonadapted V. parahaemolyticus cultures were used in the following stress challenge studies described below.
Stress challenge studies
To determine the tolerance of V. parahaemolyticus to various stresses, 1 mL of acid-adapted or nonadapted cultures was inoculated into 50 mL of PBS-3% NaCl to give an initial population of ca. 106 CFU/mL. For heat-challenge experiments, cell suspensions were subjected to a 47°C water bath. For cold-challenge experiments, cell suspensions were incubated at 4°C and −20°C, respectively. For other stress challenge experiments, cell suspensions were exposed to the following conditions at room temperature (ca. 25°C): 8% ethanol; 20% NaCl; and 20 ppm hydrogen peroxide. After different periods of exposure, V. parahaemolyticus samples were taken and serially diluted in PBS-3% NaCl and then pour plated (1 mL) with tryptic soy agar supplemented with 3% NaCl (TSA-3% NaCl). Viable colonies were counted after 18 h of incubation at 37°C to determine the survival percentage of V. parahaemolyticus.
Statistical analysis
The data obtained from the three independent experiments were expressed as mean values±standard deviations and analyzed by analysis of variance with SAS statistical software (SAS Institute, Cary, NC). The statistically significant differences (p<0.05) among the means were compared by the Duncan's multiple-range test.
Results and Discussion
Effect of acid adaptation on the tolerance of V. parahaemolyticus to heat stress
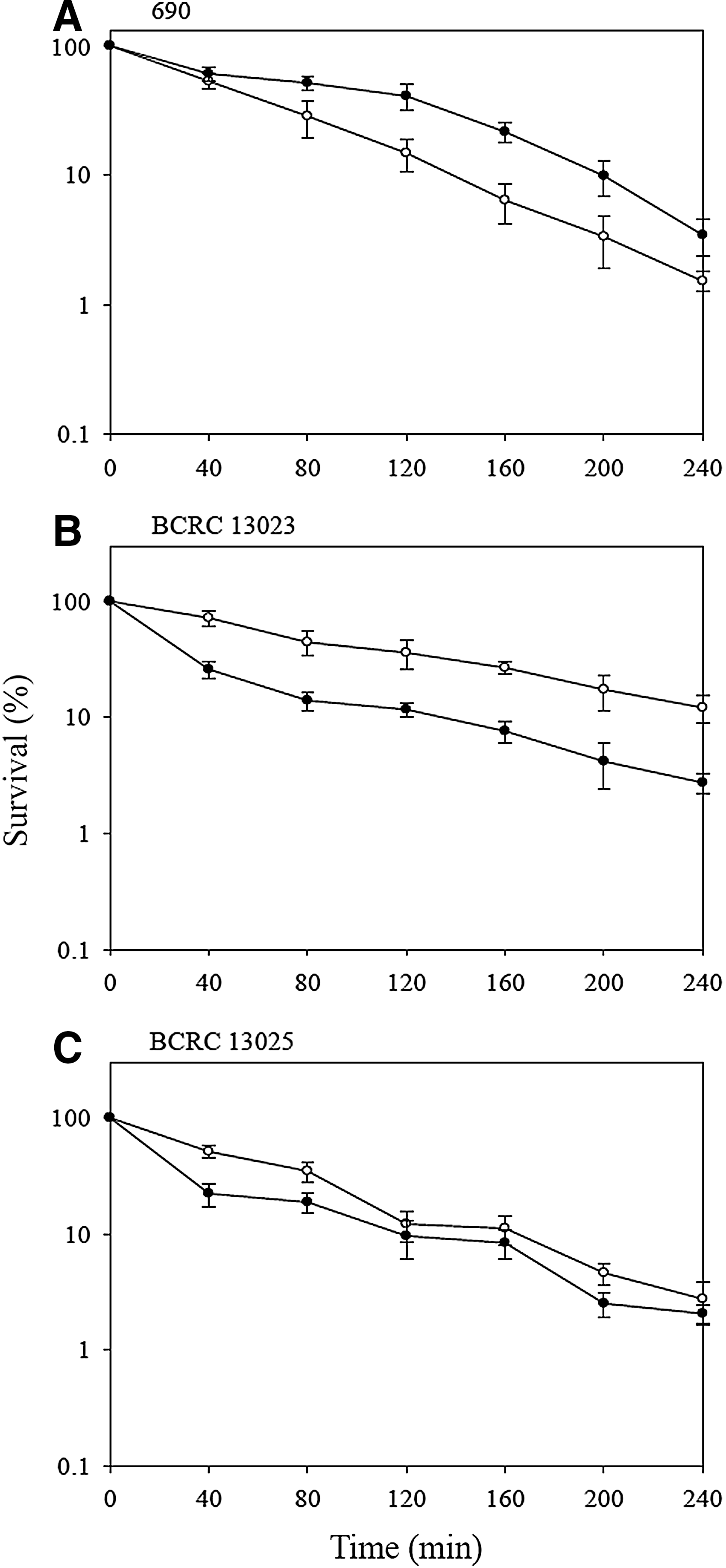
In this study, the survival of V. parahaemolyticus strains at 47°C was examined. As shown in Figure 1, the survival of V. parahaemolyticus decreased with the exposure period. Nonadapted 690, BCRC 13023, and BCRC 13025 showed a survival of 0.2%, 1.5%, and 0.2%, respectively, after exposure at 47°C for 50 min. Higher heat resistance was noted with BCRC 13023 among the three nonadapted strains. Although the magnitude of thermal tolerance varied with strains, an increase in the tolerance of V. parahaemolyticus to heat stress was noted after prior acid adaptation. The survival of acid-adapted 690, BCRC 13023, and BCRC 13025 (4.2%, 3.1%, and 3.0%) was significantly (p<0.05) higher than that of the respective nonadapted strains with the same exposure condition indicating that acid adaptation increased the thermotolerance of V. parahaemolyticus. The induction of some specific heat shock proteins, such as DnaK and GroEL in stress-adapted V. parahaemolyticus cells (Koga et al., 1996; Wong et al., 2002; Chifiriuc et al., 2011; Chiang et al., 2008, 2012a) might be responsible for the increased tolerance to high temperatures. Our results are in agreement with the results of studies on Bacillus cereus, Listeria monocytogenes, Escherichia coli, Salmonella serovar Typhimurium, and V. parahaemolyticus (Leyer and Johnson, 1993; Lou and Yousef, 1996; Koga et al., 1999; Ryu and Beuchat, 1999; Leenanon and Drake, 2001; Browne and Dowds, 2002; Cheng et al., 2002; Periago et al., 2002; Skandamis et al., 2008), indicating that acid-adapted bacteria were more heat tolerant than their nonadapted cells.
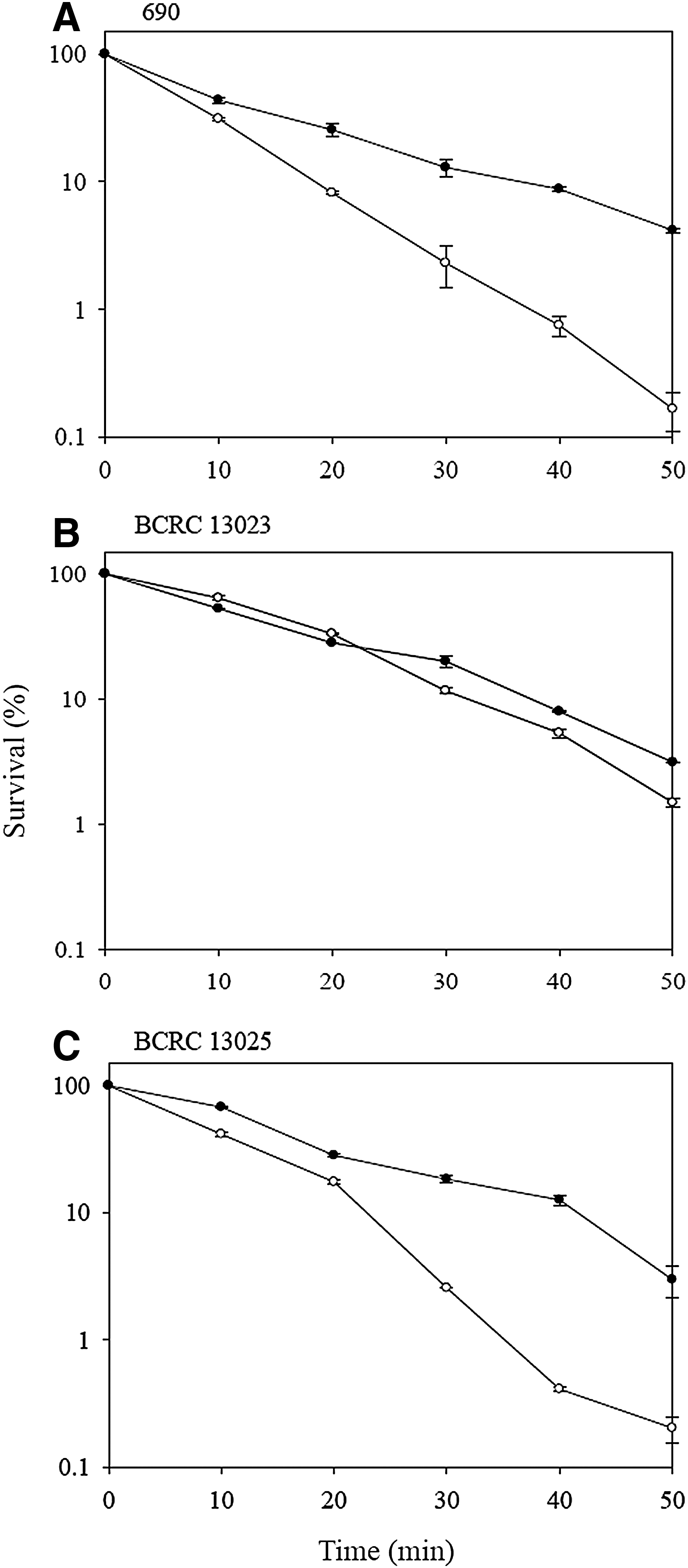
Survival of Vibrio parahaemolyticus strains (
Effect of acid adaptation on the tolerance of V. parahaemolyticus to cold stress
The survival of V. parahaemolyticus strains at 4°C is shown in Figure 2. Most V. parahaemolyticus are sensitive to low temperatures. Some strains can survive at 4°C, and their population numbers declined slightly (Miles et al., 1997; Magalhães et al., 2000). Similarly, our results showed that the survival of the three strains decreased to 42.6%–53.1% after exposure to refrigeration temperature for 10 days. Additionally, the survival between acid-adapted and nonadapted strains of V. parahaemolyticus was not significantly different (p>0.05) during the exposure period. The same result, reported by Tosun and Gönül (2003), also found no differences in survival between acid-adapted and nonadapted cells of Salmonella Typhimurium at 3°C.
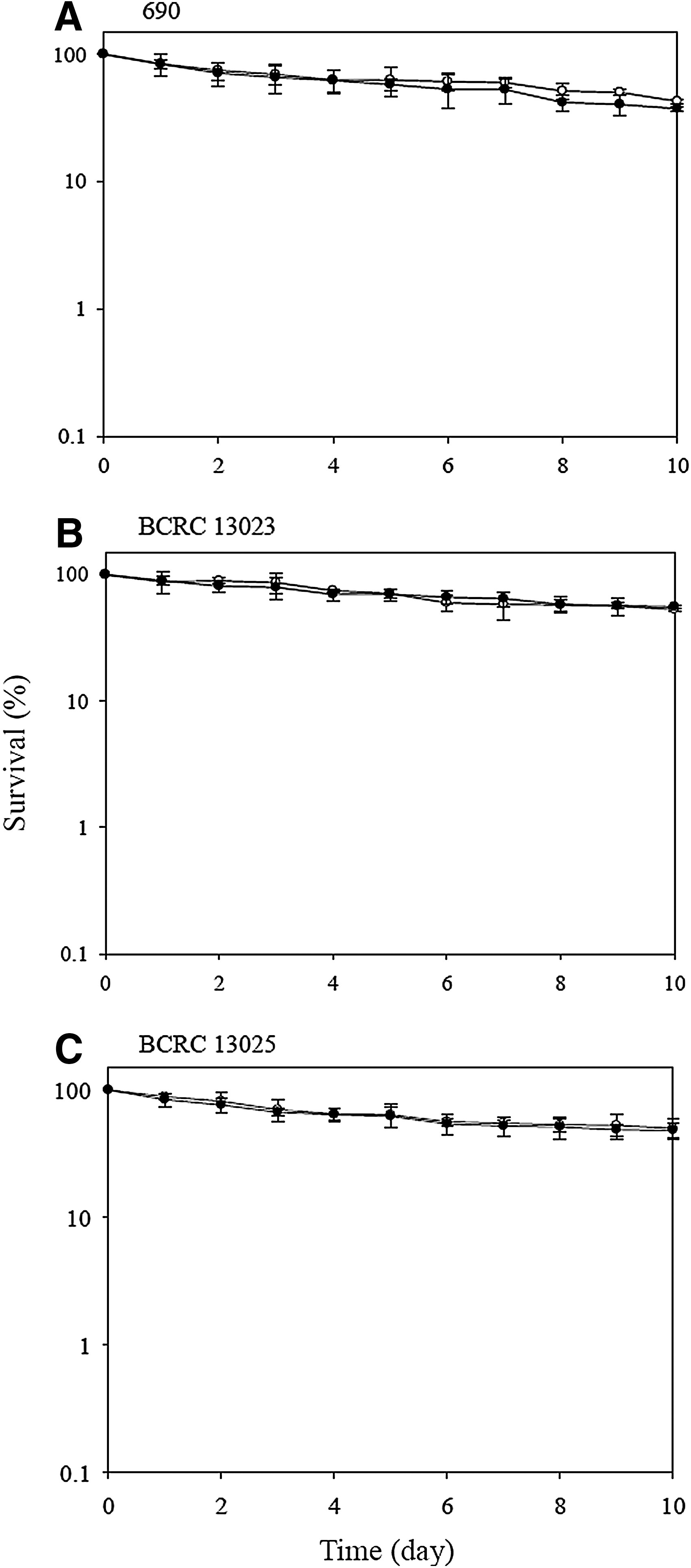
Survival of Vibrio parahaemolyticus strains (
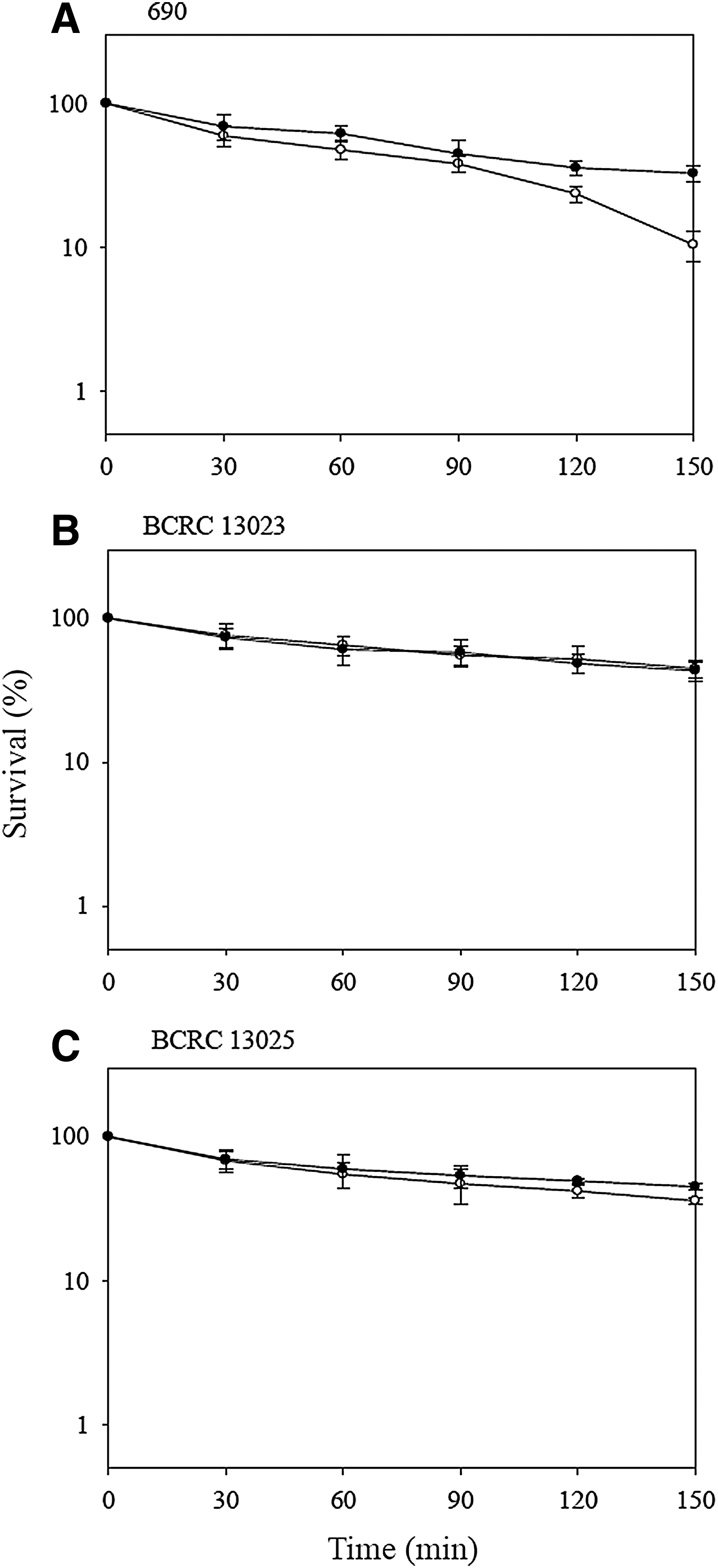
The survival of V. parahaemolyticus strains at −20°C is shown in Figure 3. Compared with the refrigeration temperature, V. parahaemolyticus showed a sharp decrease in survival during exposure to freezing temperatures. Additionally, the survival of acid-adapted V. parahaemolyticus was similar to that of its nonadapted controls in three strains after exposure at −20°C for 5 days. Nevertheless, Leenanon and Drake (2001) and Bang and Drake (2005) indicated that acid-adapted E. coli and V. vulnificus had better freeze–thaw resistance than their nonadapted cells. The diverse results may be due to the differences in bacterial species and conditions of adaptation and challenge used in our and other studies. According to our results, acid adaptation did not affect the cold tolerance of V. parahaemolyticus at 4°C and −20°C.
Survival of Vibrio parahaemolyticus strains (
Effect of acid adaptation on the tolerance of V. parahaemolyticus to ethanol stress
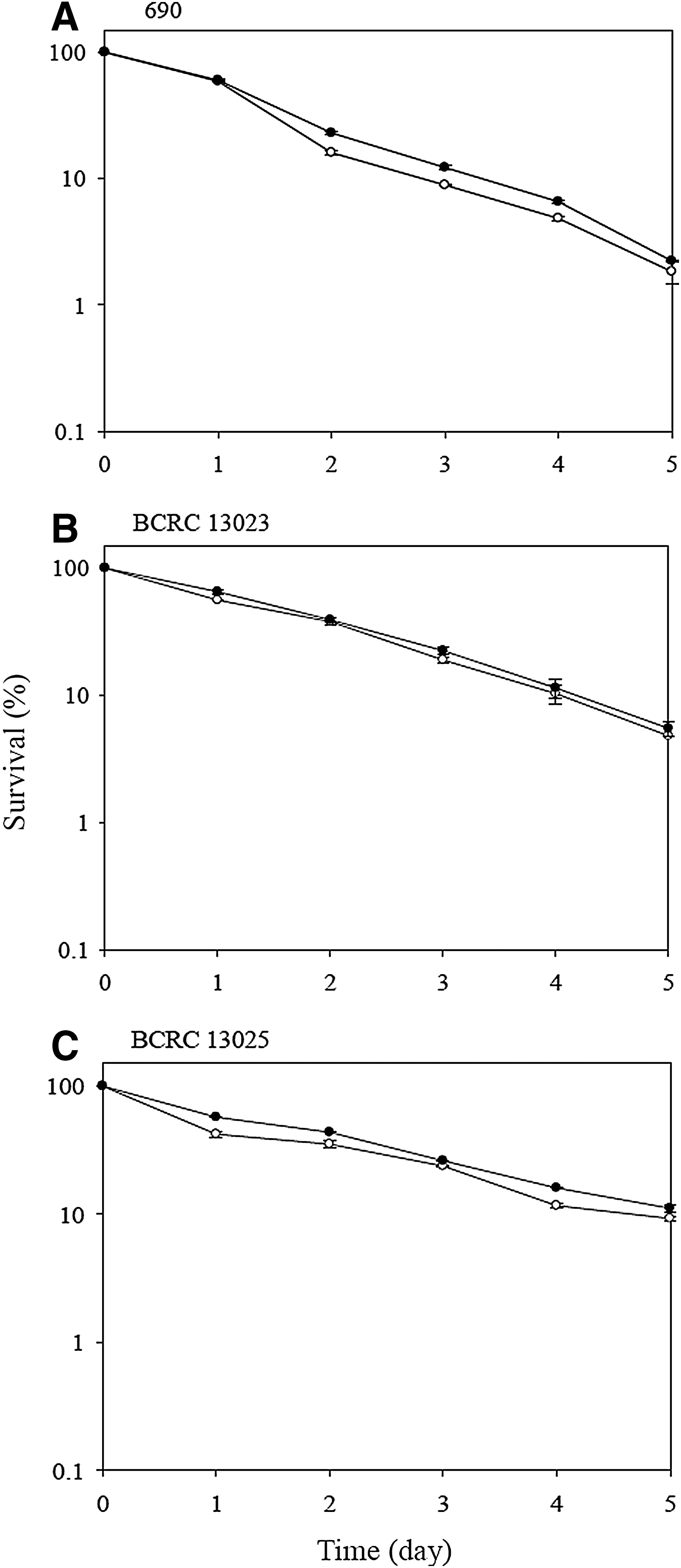
The survival of V. parahaemolyticus strains during exposure to 8% ethanol is shown in Figure 4. The survival of the three test strains declined as the exposure period extended. At the end of 150-min exposure, the survival of nonadapted strains 690, BCRC 13023, and BCRC 13025 were 10.4%, 43.3%, and 35.9%, respectively. V. parahaemolyticus BCRC 13023 showed the highest tolerance to ethanol, followed by BCRC 13025 and 690. In addition, acid-adapted 690 showed a significantly higher (p<0.05) survival of 32.7%, which was 3.1-fold that of its nonadapted control after exposure to ethanol. Similar survival was observed in BCRC 13025. The findings observed in this study are consistent with those described by O'Driscoll et al. (1996), Lou and Yousef (1997), and Browne and Dowds (2002) on L. monocytogenes and B. cereus, respectively. Such an increase in ethanol tolerance may be involved in the induction of stress proteins and alteration of membrane fatty-acid composition in acid-adapted cells (Leyer and Johnson, 1993; van Schaik et al., 1999; Quivey et al., 2000). However, the survival between acid-adapted and nonadapted BCRC 13023 was not significantly (p>0.05) different. These results showed that acid adaptation increased the tolerance of V. parahaemolyticus to ethanol in strains 690 and BCRC 13025, but not in BCRC 13023.
Survival of Vibrio parahaemolyticus strains (
Effect of acid adaptation on the tolerance of V. parahaemolyticus to high-salt stress
Figure 5 shows the survival of V. parahaemolyticus strains during exposure to 20% NaCl. The survival of the test strains decreased with the exposure period. At the end of 240-min exposure, nonadapted 690 showed a survival of 1.5%. At the same time, the survival of acid-adapted 690 (3.5%) was significantly higher (p<0.05) than its nonadapted control, indicating that acid adaptation increased the tolerance of V. parahaemolyticus 690 to high-salt condition. Several studies have also reported that acid-adapted L. monocytogenes and E. coli had better high-salt resistance than their nonadapted cells (Lou and Yousef, 1997; Cheng et al., 2002; Faleiro et al., 2003). In contrast to strain 690, acid adaptation increased the susceptibility of V. parahaemolyticus BCRC 13023 to high salts. The survival of acid-adapted V. parahaemolyticus BCRC 13023 (2.7%) was significantly lower (p<0.05) than that of its nonadapted control (12.1%) after 240-min exposure to 20% NaCl. This finding is in accordance with Chen et al. (2009b), who found that acid-adapted B. cereus was more susceptible to high concentrations of NaCl than its nonadapted cells. However, the survival between acid-adapted and nonadapted V. parahaemolyticus BCRC 13025 was not significantly different (p>0.05) in high-salt stress. In the results of this study, the tolerance of V. parahaemolyticus to high salts had significant differences between the strains after acid adaptation and seems to be strain dependent. Rowe and Kirk (1999) also reported that acid-adapted E. coli O157:H7 had various adaptive salt tolerances among strains. This phenomenon may be related to their different abilities to respond to severe osmotic stress such as accumulation of compatible solutes, regulation of membrane transport protein activity, and induction of σfactor RpoS (Gould, 1989; Gutierrez et al., 1995; Yousef and Courtney, 2003; Beales, 2004; Vasudevan and Venkitanarayanan, 2006). Differences in protein number and expression might be another cause of different survival between acid-adapted or nonadapted strains of V. parahaemolyticus with the tested environmental stresses (Chiang et al., 2012a).
Survival of Vibrio parahaemolyticus strains (
Effect of acid adaptation on the tolerance of V. parahaemolyticus to hydrogen peroxide stress
This study investigated the survival of V. parahaemolyticus strains during exposure to 20 ppm H2O2. As shown in Figure 6, the survival of V. parahaemolyticus decreased as the exposure time was extended. After exposure for 15 min, strain BCRC 13025 exhibited the highest survival (10.9%) and 690 had the lowest survival (1.0%) among the three nonadapted strains. Additionally, the survival of nonadapted strains of V. parahaemolyticus was significantly higher (p<0.05) than their acid-adapted controls. For example, nonadapted BCRC 13023 showed a survival of 4.92%, which was 4.03-fold that of its acid-adapted counterpart (1.2%), at the end of exposure. Results showed that acid adaptation decreased the tolerance of V. parahaemolyticus BCRC 13023 to hydrogen peroxide. Similar survival results were also observed in strains 690 and BCRC 13025. Some studies noted that acid adaptation did not affect the resistance of B. cereus to hydrogen peroxide (O'Driscoll et al., 1996; Browne and Dowds, 2002; Chen et al., 2009c). Nevertheless, Lin et al. (2004) found that cold-shocked V. parahaemolyticus was more susceptible to hydrogen peroxide than its control cells. The oxidative susceptibility of sublethal-stressed V. parahaemolyticus may be due to injury of cell membrane and change of cellular antioxidants and related genes expression (Lai and Wong, 2013).

Survival of Vibrio parahaemolyticus strains (
Conclusions
The results showed that the effect of acid adaptation on the survival of V. parahaemolyticus varied among strains and types of stress challenge. Acid adaptation increased the thermal tolerance of V. parahaemolyticus in the three test strains. The tolerance of V. parahaemolyticus to ethanol and high salts had significant differences between the strains after prior acid adaptation. However, the survival of acid-adapted V. parahaemolyticus strains was lower than their nonadapted controls after exposure to hydrogen peroxide. These findings are important in understanding the survival of V. parahaemolyticus exposed to acid and other stress treatments in food processing and preservation.
Footnotes
Acknowledgments
This research was funded by the project of National Science Council, Executive Yuan (Taipei, Taiwan) (NSC 102-2313-B-424-001-MY3).
Disclosure Statement
No competing financial interests exist.