
Abstract
Different structures related to biofilm formation by Shiga toxin–producing Escherichia coli (STEC), particularly O157 strains, have been described, but there are few data regarding their involvement in non-O157 strains. The aim of this study was to determine the ability of 14 O157 and 8 non-O157 strains isolated from bovine hide and carcass to interact with biotic and abiotic surfaces and also to evaluate the role of different adhesins. Biofilm formation assays showed that four O157 and two non-O157 strains were able to adhere to glass, and that only one O157 strain adhered to polystyrene. Reverse transcriptase–polymerase chain reaction was carried out using biofilm-forming strains to determine the expression of antigen 43 (Ag43), curli, type 1 fimbriae, STEC autotransporter contributing to biofilm formation (Sab), calcium-binding antigen 43 homologue (Cah), and autotransporter protein of enterohemorrhagic E. coli (EhaA). Most of these structures were expressed under biofilm conditions. However, the lack of Ag43 in one non-O157 strain, as well as Cah and EhaA in two O157 strains, suggests that other adhesins are involved in biofilm formation in these strains. Despite the fact that adherence to HeLa cells was detected in 20 strains (91%), it was not possible to correlate biofilm formation with adherence patterns. Invasiveness in T84 and Caco-2 cells was observed in four and three O157 strains, respectively. Altogether, we showed that there are different sets of genes involved in the interactions of STEC with biotic and abiotic surfaces. Interestingly, one O157 strain that was able to form biofilm on both glass and polystyrene also adhered to and invaded human cells, indicating an important route for its persistence in the environment and interaction with the host. Additionally, the ability of non-O157 strains not carrying the LEE pathogenicity island to form biofilm highlights an industrial and health problem that cannot be neglected.
Introduction
S
Materials and Methods
Bacterial strains
Eleven O157:H7 and six non-O157 STEC strains were isolated from 10 healthy cattle over a period of 2 years as described by Lascowski et al. (2012). Strains from the same animal were isolated from both the hide and the carcass. Three O157:H7 strains devoid of stx genes isolated from the carcass of animals carrying O157:H7 STEC strains in hides were also included in the study. There was no clonal relation between the strains studied (unpublished data). For all assays, strains were statically grown overnight at 37°C on T Medium (Triptose, 10 g; beef extract, 3 g; NaCl, 5 g/L) broth, and bacterial cultures were standardized to an OD600nm of 0.4.
Detection of genes associated with biofilm formation by polymerase chain reaction (PCR) assays
All isolates were analyzed by PCR for the genes of curli structural subunit and curli regulator (csgA and crl, respectively) (Maurer et al., 1998), type 1 fimbriae (fimH) (Johnson and Stell, 2000), antigen 43 (flu) and calcium-binding antigen 43 homologue (cah) (Restieri et al., 2007), Eha passenger and translocation domain (ehaAα and ehaAβ, respectively) (Wells et al., 2008), and STEC autotransporter contributing to biofilm formation (sab) (Herold et al., 2009). DNA was extracted from bacterial cells by heat lysis.
Detection of type 1 fimbriae and curli by phenotypical assays
Expression of type 1 fimbriae (T1F) and curli was identified as previously described (Biscola et al., 2011). Two non-O157 STEC strains formed both curli-expressing (red colony, curli+) and nonexpressing (white colony, curli−) colonies on colonization factor antigen (CFA)–Congo Red (CR) agar, and these two variants were purified and used in the study as separate cultures. E. coli Ec473/01 (Biscola et al., 2011) and E. coli HB101 were used as curli-positive and -negative controls, respectively.
Biofilm formation
The assays for quantifying biofilm formation were performed in two systems: 96-well polystyrene microtiter plates (Corning Costar Corporation, Cambridge, MA) by the method of Uhlich et al. (2006) with some modifications, and brand-new glass tubes (16×100 mm, Pyrex, Corning Costar Corporation, Cambridge, MA) by the method of Wang et al. (2012) with modifications. Both assays were carried out using standardized cultures, and incubation was performed at 28°C for 48 h. After incubation, polystyrene plates were washed three times with de-ionized water, fixed by heating at 70°C for 60 min, and stained with 0.1% crystal violet (Merck SA, Rio de Janeiro, Brazil). In glass assays, bacterial cultures were discarded after incubation, and tubes were washed and stained with 1% crystal violet after incubation. In both methods, the dye was solubilized with 95% ethanol (Merck) and absorbance was read at 540 nm in an enzyme-linked immunosorbent assay plate reader (Labsystems Multiskan MS). Each data set obtained in the polystyrene and glass assays was the average of results from three replicate wells. The experiments were repeated three times using independent cultures. Strain E. coli Ec473/01 (Biscola et al., 2011) was used as positive control.
Expression of structures associated with biofilm formation by reverse-transcriptase PCR (RT-PCR)
Expression of genes associated with biofilm formation assessed by conventional PCR was indirectly determined by RT-PCR in strains able to form biofilm on polystyrene and glass surfaces. The primer sequences employed were as described above. Accordingly, after bacterial growth under biofilm formation conditions and prior to the staining step, RNA was extracted using the RNeasy Mini Kit (Qiagen, Valencia, CA) as per manufacturer's instructions. During extraction, RNase-Free DNase (Qiagen) was added to the column for RNA digestion for 60 min at room temperature. Primers 8F and 1493R for 16S ribosomal protein (Turner et al., 1999) were used to confirm total digestion. To obtain cDNA and amplification of targets, the SuperScript™ One-Step RT-PCR system (Invitrogen, Carlsbad, CA) was used according to the manufacturer's specifications.
Bacterial interactions with human cells
Adherence to HeLa (ATCC CCL2) cells was tested as previously described (Mora et al., 2009) using only the 6-h protocol. Invasion assays were performed as described by Robbins-Browne and Bennet-Wood (1992) with modifications. Initially, a screening assay with undifferentiated (growth in 5 days) Caco-2 (ATCC HTB-37) cells was used to determine the invasive ability of all strains. Strains showing significant invasion percentage when compared with control were submitted to new assays using differentiated (growth in 14 days) Caco-2 and T84 (ATCC 248) cells. These cell lines were maintained, respectively, in Dulbecco's Modified Eagle Medium (DMEM) and Dulbecco's Modified Eagle Medium/Ham's F-12 (DMEM/F12) (Gibco, Grand Island, NY) supplemented with 10% fetal bovine serum, and 1% penicillin–streptomycin–neomycin antibiotic mixture (Gibco). For the invasion assay, a 40-μL aliquot of the strain was added to four wells. Plates were incubated for 4 h at 37°C. After washing cells, fresh medium without antibiotic was added to two wells of each strain, and extracellular bacteria were killed by adding fresh medium containing gentamicin (100 mg/mL) in the other two wells of each strain. After further incubation for 2 h, the cells were washed and lysed with 1% Triton X-100. Appropriate bacterial dilutions were plated on MacConkey agar (Difco BD, Sparks, MD) to determine the number of viable internalized bacteria. Invasion indices (%) were calculated using the equation (ICB/AB)×100, where ICB is the number of intracellular bacteria and AB is the number of cell-associated bacteria. All tests were performed three times in duplicate. E. coli EH 41 (Doughty et al., 2002) was used as positive control and a strain of Salmonella enterica serovar Typhimurium was used as a control of the test.
Statistical analysis
Results of interactions with biotic and abiotic surfaces were compared with the positive control by Student t-test, and considered significant when p<0.05. Strains with no significant difference were classified as similar to the positive control.
Results
Genotypic and phenotypic detection of structures associated with biofilm formation
The detection of genes associated with biofilm formation and their expression were carried out in O157 and non-O157 STEC strains, and the results are presented in Table 1. The curli related genes csgA and crl were identified in all 22 strains, but only non-O157 (5/6) strains expressed this fimbriae in CFA-CR assays, and two of them exhibited curli variants. The gene sequences related to autotransporter proteins Cah (cah) and EhaA (ehaAα and ehaAβ) were detected in all strains as well as the sequence associated with T1F synthesis (fimH). However, T1F expression was detected only in seven (32%) strains, all of them being non-O157. Antigen 43 (flu) and Sab (sab) genes were only detected in non-O157 strains (100% and 12.5%, respectively).
Strains devoid of stx genes.
+, Curli-positive variant strains;−, curli-negative variant strains.
NT, nontypable with antisera against O1 to O181 serogroups.
H, hide; C, carcass; LAL, localized-adherence-like; AA, aggregative adherence; DA, diffuse adherence; NA, non-adherent.
Ability to form biofilm on abiotic surfaces
The ability to form biofilm on polystyrene surface was only identified in one O157 STEC strain (Fig. 1A). When examined in glass tubes, six strains (O157, n=4; and non-O157, n=2) were able to form biofilm (Fig. 1B). The only O157 STEC strain capable of forming biofilm on the polystyrene surface was also able to form biofilm on the glass surface.
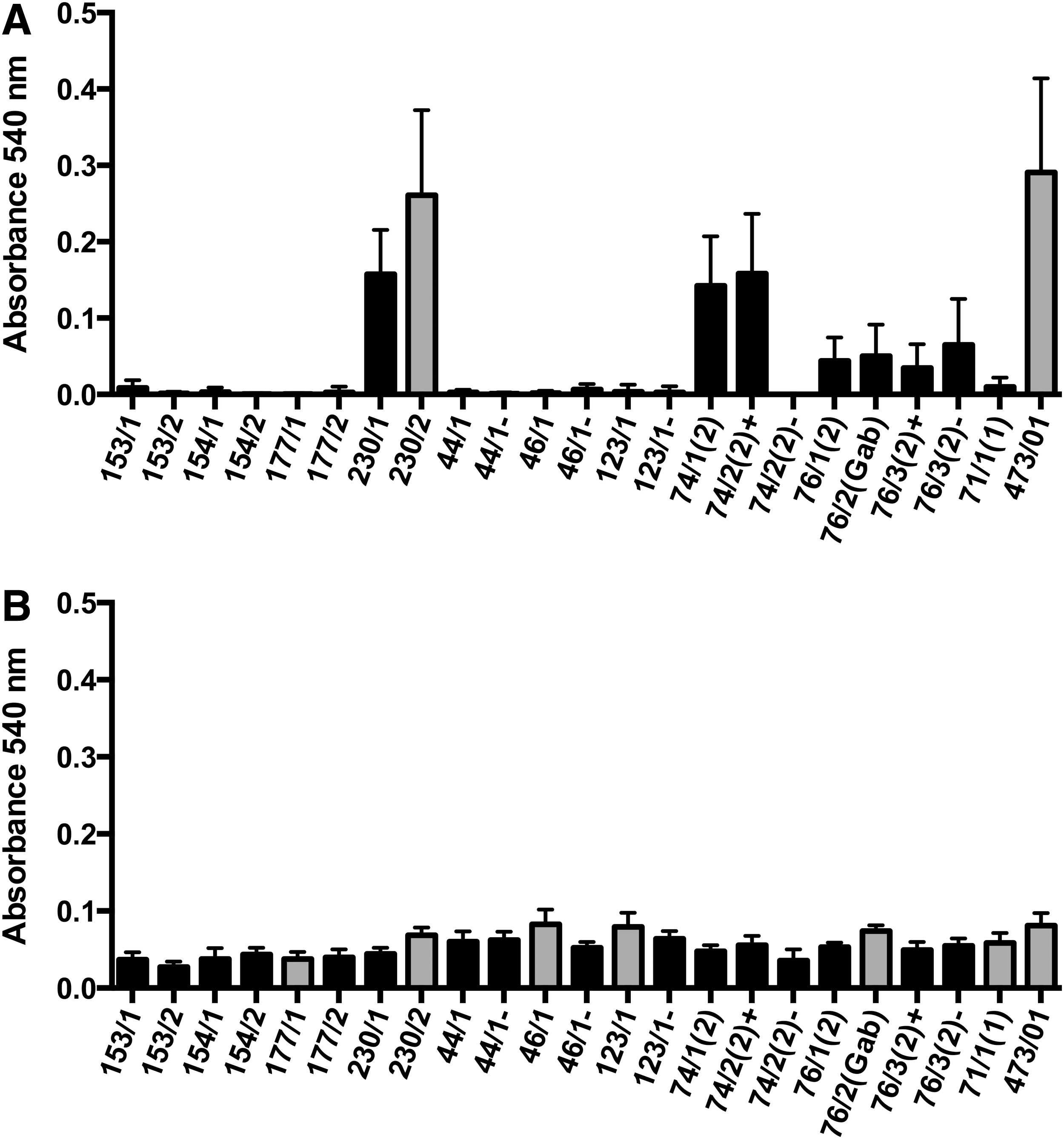
Quantitative comparison of biofilm formation on polystyrene
Expression of genes associated with biofilm formation
The only O157 STEC strain able to form biofilm on both surfaces exhibited a similar gene expression profile, except for ehaAα, which was not expressed in the glass assay (Table 2). All strains showed expression of csgA and ehaAβ genes under biofilm formation on glass. Expression of cah was detected in six strains, whereas expression of fimH, flu, and sab genes was only observed in non-O157 strains (Table 2).
P, polystyrene; G, glass; NT, not tested as these strains were negative when previously tested PCR.
Ability to adhere to epithelial cells
Most O157 strains displayed the localized-adherence-like (LAL) pattern (Table 1), and only one O157 strain showed a mixed pattern with LAL and aggregative adherence (AA) (Fig. 2). Different adherence behavior was observed in the non-O157 strains. Three strains demonstrated the diffuse adherence (DA) pattern, and mixed patterns (AA/DA) were observed for three other strains, and two isolates were nonadherent (Table 1).
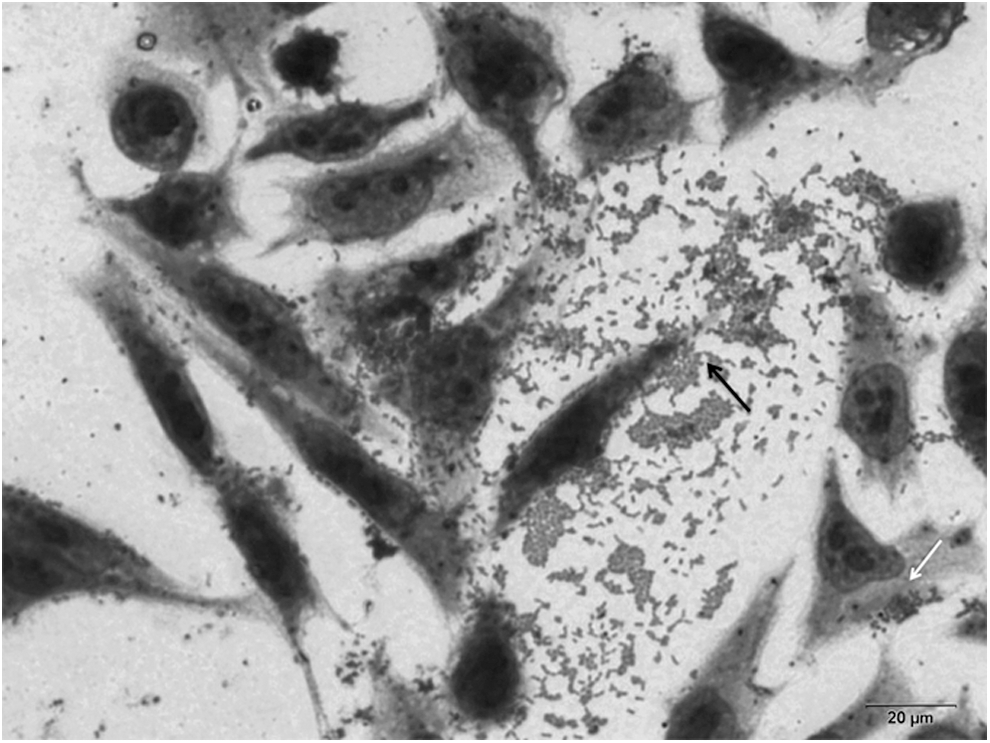
Mixed pattern of adherence of O157:H7 Shiga toxin–producing Escherichia coli 153/2 isolate on HeLa cells after 6 h of incubation. The white arrow indicates the localized adherence-like pattern, characterized by loose and compact clusters of bacteria, and the black arrow indicates the aggregative pattern, characterized by a stacked-brick-like arrangement on the cell surface as well as in the coverslips.
Invasion of Caco-2 and T84 cells
In the screening assays using undifferentiated Caco-2 cells, only five (22.7%) strains, all belonging to the O157 serogroup, showed invasive properties. When these five strains were subjected to assays using differentiated cells, a decrease in invasion rates was observed for all strains in both cell lines (Fig. 3). Invasion indices were significantly higher in T84 cells except for one strain (154/2). Only one strain did not have the ability to invade this cell line when compared to the positive control. In Caco-2 cells, two strains did not exhibit invasive properties (Fig. 2).
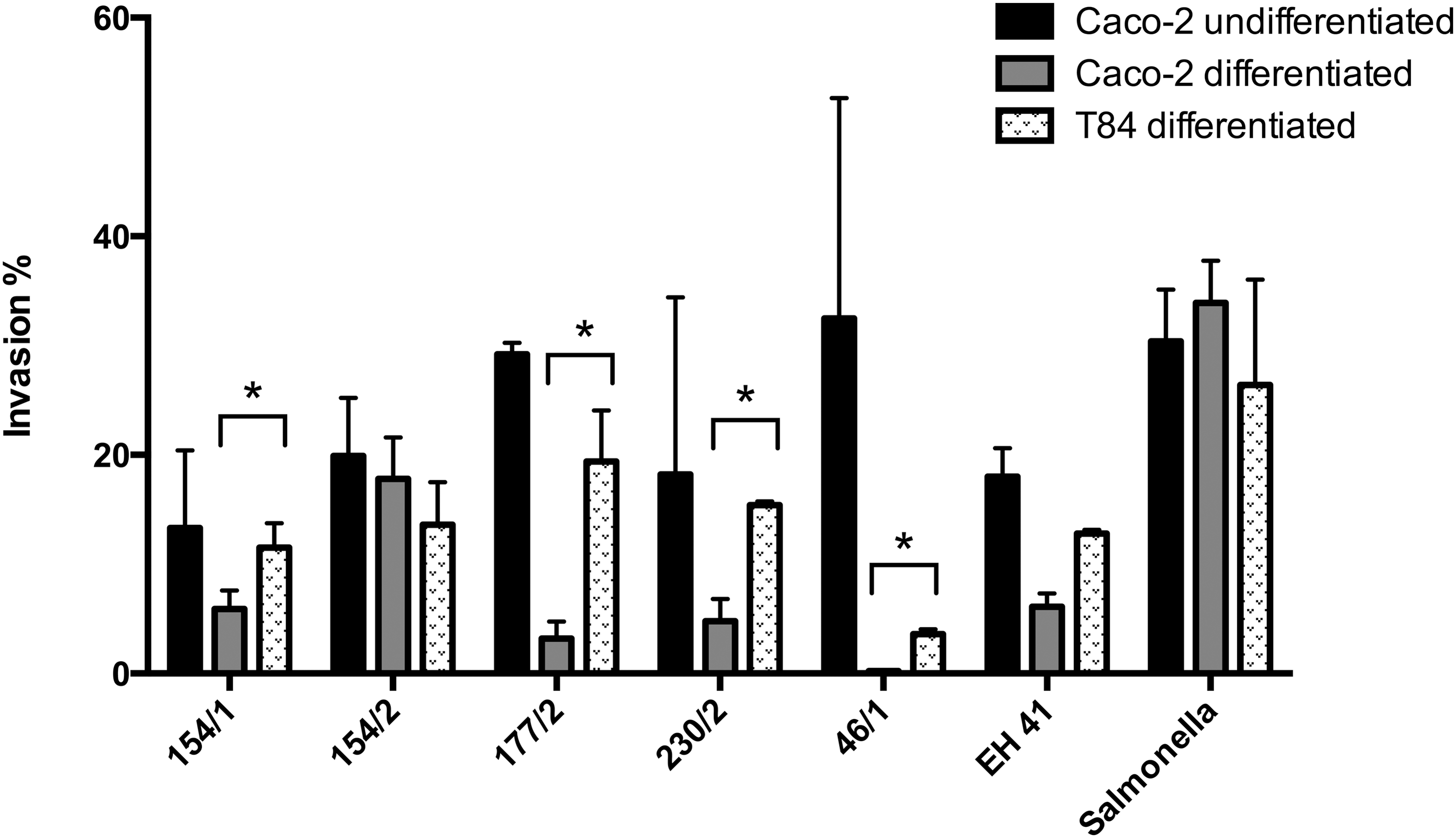
Comparative invasion percentage in Caco-2 and T84 cells by O157:H7 Shiga toxin–producing Escherichia coli strains after 4 h of infection and gentamicin protection assay. Each bar represents the mean of three independent assays performed in duplicate. Asterisk indicates statistically significant difference when the two cell lines were compared.
Discussion
Biofilm formation is an important way that some microorganisms persist in the environment. Several studies have demonstrated the ability of STEC strains to form biofilms, but they usually involved O157:H7 strains (Torres et al., 2005; Uhlich et al., 2006; Saldaña et al., 2009; Simpson Beauchamp et al., 2012; Carter et al., 2012) or the LEE-positive O26, O111, O145, O103, and O91 serogroups (Wang et al., 2012; Fouladkhah et al., 2013). Few studies have described the ability of non-O157 STEC strains isolated from different sources, including animal samples, to form biofilms (Cookson et al., 2002; Biscola et al., 2011). To our knowledge, this is the first study to evaluate the ability of several O157 and non-O157 STEC strains isolated from bovine hide and carcass to form biofilm. This property was identified in 6 of the 22 (27%) strains studied, and it was mainly observed in isolates recovered from animal hide. It is not clear why animal hide strains formed increased levels of biofilms and is not explored within this study. Nevertheless, the fact that STEC strains are able to form biofilms on an animal carcass is an important piece of information related to potential downstream contamination. We observed that four O157 and two non-O157 isolates were able to form biofilm at 28°C on glass surface, and only one O157 STEC strain was able to form biofilm on polystyrene surface. Wang et al. (2012) also demonstrated a reduced capacity of O157 strains to form biofilm on polystyrene surfaces, but they found that these strains had low or no ability to adhere to glass. Low levels of adherence of O157 strains to polystyrene were also detected by Rivas et al. (2007), while Biscola et al. (2011) described higher frequencies of biofilm-forming strains. Bacterial surface hydrophobicity is an important factor related to adherence and can be influenced by surface proteins, including fimbriae and flagella (Gross and Logan, 1995). Glass as well as some metal surfaces have a hydrophilic surface (Rose et al., 2003), and the ability of the STEC strains isolated from bovine hide and carcass described herein to form biofilm on glass surface indicates a possible and important way that these strains persist in a meat production plant, which may lead to cross contaminations. The only one O157 STEC strain (230/2) that produced biofilm on polystyrene was also able to adhere on glass, indicating the important ability of this particular strain to interact with different abiotic surfaces. This certainly suggests that different sets of adhesive structures in STEC isolates may be related to biofilm formation on different surfaces, and further studies will be necessary to identify what these structures might be. The relation between biofilm formation on polystyrene and curli expression detected by CFA-CR has been previously described in O157 STEC strains (Biscola et al., 2011). Although this association was confirmed in the O157 as well as the non-O157 strains presently studied, it was interesting to observe that curli expression was only detected by RT-PCR under biofilm formation on glass or polystyrene conditions. This finding suggests that CFA-CR assays may not be adequate to analyze the role of curli fimbriae in the mechanism of biofilm formation.
The expression of genes detected in this study by RT-PCR suggests that most of them may be involved in the biofilm formation process. However, it was observed that some strains did not express all genes previously detected by PCR. Despite the fact that flu has been associated with bacterial autoaggregation and biofilm formation (Danese et al., 2000; Kjaergaard et al., 2000) we observed that only one non-O157 biofilm-forming strain expressed the Ag 43. Interestingly, the strain that lacked Ag 43 was able to express cah and sab genes, which may be a compensatory mechanism in the biofilm process, since Sab has been described as promoting biofilm formation (Herold et al., 2009) and Cah shows homology with Ag 43 (Torres et al., 2002). Although Biscola et al. (2011) reported that the expression of Cah was absent or low in non-O157 and O157 strains, respectively, the expression of Cah was observed in both non-O157 strains and all O157 biofilm-forming strains studied herein, except for the 177/1 isolate. Indeed, the differences observed in the expression of Cah could be related to the method used, since Biscola et al. (2011) did not study Cah expression under biofilm conditions. Apart from the 177/1 isolate, it was also observed that another O157 strain (230/2) did not express the ehaAα gene when subjected to biofilm formation on glass. The 230/2 strain also produced biofilm on a polystyrene surface, but ehaAα was expressed under this condition. The particular behavior displayed by 230/2 isolate indicates that the expression of genes associated with biofilm formation may depend on the material surface.
Mixed patterns of adherence to HeLa cells such as LAL/AA and DA/AA were detected in O157 and non-O157 strains, respectively. Although uncommon, the aggregative pattern has been described in an O157 (Kim and Kim, 2004) and in some non-O157 strains (Aslani and Bouzari, 2009; Mora et al., 2009). The presence of AA suggests an ability of a bacterial strain to aggregate and form biofilm. However, in the present study no correlation was found between the AA pattern, observed in some STEC strains, and the ability to form biofilm. In fact, although most of the strains studied were able to adhere to human cells, and few had the ability to interact with abiotic surfaces, suggesting that the apparatus conferring adherence to human cells differs from that promoting adherence to abiotic surfaces.
There are few studies describing the ability of a group of STEC strains isolated from different sources to invade human cells. In a study by Luck et al. (2005), the invasiveness of STEC strains was usually associated with non-O157 serotypes, while O157 strains were unable to invade human cells. Since Gabastou et al. (1995) reported that the invasion rates were not decreased by the use of undifferentiated Caco-2 cells, with efficient cell-entry of bacteria being decreased only in fully differentiated cells, we initially performed invasion assays using undifferentiated Caco-2 cells. Using this approach, it was observed that only five strains, all belonging to the O157:H7 serotype, displayed invasive ability, in contrast to the observations by Luck et al. (2005). Further assays carried out with two differentiated cell lines showed that the O157 strains had a higher capacity to invade T84 compared to Caco-2 cells, with invasion rates ranging from 3.6% to 20.2% and 0.22% to 17.8%, respectively. Considering that T84 cells are similar to colonic crypt cells and that Caco-2 cells represent small-intestinal epithelial cells (Nataro et al., 1996), the results obtained here are in agreement with previous observations showing that STEC strains exhibit tropism toward colonic tissues (Stevens et al., 2002; Naylor et al., 2003; van Diemen et al., 2005).
Conclusions
The data presented in this study suggest that STEC strains can have different sets of structures involved in the mechanism of biofilm production on different surfaces. Additionally, the ability of non-O157 strains that do not carry the LEE pathogenicity island to form biofilm highlights an industrial and health problem that cannot be neglected. Moreover, the detection of an O157 STEC strain that is able to form biofilm on different surfaces and adhere to and invade human cells indicates an important ability to persist in the environment and to interact with the host. Further studies will be conducted to better understand the interactive behavior of this O157 isolate.
Footnotes
Acknowledgments
This work was supported by grants from Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Disclosure Statement
No competing financial interests exist.