
Abstract
Salmonella is an important cause of foodborne illness; however, identifying the source of these infections can be difficult. This is especially true for Salmonella serotype Typhimurium, which is found in diverse agricultural niches. Extended-spectrum cephalosporins (ESC) are one of the primary treatment choices for complicated Salmonella infections. In Salmonella, ESC resistance in the United States is mainly mediated by bla CMY genes carried on various plasmids. In this study, we examined whether the characterization of bla CMY plasmids, along with additional information, can help us identify potential sources of infection by Salmonella, and used serotype Typhimurium as a model. In the United States, monitoring of retail meat, food animals, and ill persons for antimicrobial-resistant Salmonella is conducted by the National Antimicrobial Resistance Monitoring System. In 2008, 70 isolates (70/581; 12.0%) (34 isolates from retail meat, 23 food animal, and 13 human) were resistant to ceftriaxone and amoxicillin/clavulanic acid. All were polymerase chain reaction (PCR)–positive for bla CMY and 59/70 (84.3%) of these genes were plasmid encoded. PCR-based replicon typing identified 42/59 (71.2%) IncI1-bla CMY plasmids and 17/59 (28.8%) IncA/C-bla CMY plasmids. Isolates from chickens or chicken products with bla CMY plasmids primarily had IncI1-bla CMY plasmids (37/40; 92.5%), while all isolates from cattle had IncA/C-bla CMY plasmids. Isolates from humans had either IncA/C- bla CMY (n=8/12; [66.7%]) or IncI1- bla CMY (n=4/12 [33.3%]) plasmids. All of the IncI1-bla CMY plasmids were ST12 or were closely related to ST12. Antimicrobial susceptibility patterns (AST) and pulsed-field gel electrophoresis (PFGE) patterns of the isolates were also compared and differences were identified between isolate sources. When the source of a Typhimurium outbreak or sporadic illness is unknown, characterizing the outbreak isolate's bla CMY plasmids, AST, and PFGE patterns may help identify it.
Introduction
S
The National Antimicrobial Resistance Monitoring System (NARMS) determines antimicrobial susceptibility of Salmonella from humans, retail meats, and food animals. In 2008, Salmonella Typhimurium was the most common serotype isolated from retail chicken breasts, the fourth most common from chickens, the fifth most common from cattle, and the second most common serotype from humans (USDA, 2009; FDA, 2009a; CDC, 2010b). Antimicrobial resistance in serotype Typhimurium is associated with bloodstream infection, which is of concern because these patients are more likely to require antimicrobial treatment (Crump et al., 2011). Extended-spectrum cephalosporins (ESC), such as ceftriaxone, are one of the primary treatment choices for invasive salmonellosis (RedBook, 2012). However, ESC resistance among Salmonella is on the rise in the United States and threatens to complicate treatment options (FDA, 2009b). Among ceftriaxone-resistant Salmonella collected in the United States in 2008, Typhimurium was the second most common serotype found in humans, the first from chicken retail meat, the second from chickens, and the fourth from cattle.
Considerable research has been performed on identifying the mechanisms of cephalosporin resistance in Salmonella. In the United States, cephalosporin resistance is primarily mediated by AmpC β-lactamases, encoded by bla CMY genes (Philippon et al., 2002; Folster et al., 2010, 2011). These genes are commonly carried on various types of plasmids, which can be distinguished by their incompatibility/replicon type (Carattoli et al., 2005). Previous studies identified two major types of bla CMY plasmids among Salmonella in the United States: IncA/C and IncI1 (Folster et al., 2010, 2011). In this study, we examined bla CMY-positive Typhimurium isolates from retail meat, food animals, and humans to determine whether the phenotypic and genotypic characteristics of bla CMY plasmids can help us identify possible sources of infection by Typhimurium. We chose to focus on serotype Typhimurium isolates due to their commonality and widespread sources; however, it is our hope that this study will serve as a model of all bla CMY-positive Salmonella serotypes.
Methods
Isolate collection and testing
Salmonella isolates from ill persons were obtained from specimens submitted to clinical laboratories in the United States and forwarded to state public health laboratories. Participating state public health laboratories serotyped and submitted every 20th nontyphoidal Salmonella to the Centers for Disease Control and Prevention (CDC) National Antimicrobial Resistance Monitoring System (NARMS) laboratory for susceptibility testing. NARMS retail meat monitoring was conducted by the United States Food and Drug Administration (FDA)–Center for Veterinary Medicine as previously described (Zhao et al., 2008). NARMS monitoring of food animals at slaughter was conducted by the U.S. Department of Agriculture (USDA) Bacterial Epidemiology and Antimicrobial Resistance Research Unit of the Agricultural Research Service as previously described (Frye et al., 2008).
Broth microdilution (Sensititre®, Trek Diagnostics Systems, Thermo Fisher Scientific Inc., Cleveland, OH) was used to determine the minimum inhibitory concentrations for 15 antimicrobial agents. Resistance was defined by the Clinical and Laboratory Standards Institute (CLSI) interpretive standards, when available (CLSI, 2013). For streptomycin, where no CLSI interpretive criteria for human isolates exist, the resistance breakpoint is 64 μg/mL (FDA, 2009b). Testing was performed according to the manufacturer's instructions; the following quality-control strains were used: Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 29213, E. coli ATCC 35218, and Pseudomonas aeruginosa ATCC 27853.
Polymerase chain reaction (PCR) amplification of blaCMY genes
DNA templates for PCR were prepared by lysing the bacteria at 95°C and collecting the supernatant following centrifugation for 10 min at 20,000×g (Sorvall RC5B Plus, SS-34 rotor, Thermo Fischer Scientific Inc., Waltham, MA). PCR reactions contained 2x HotStar PCR Master Mix (Qiagen Inc., Valencia, CA), 0.4 μM of each primer, 5 μL template DNA, and sterile PCR water to a final volume of 50 μL. Thermal cycling was performed using the following conditions: 15 min at 95°C, followed by 30 cycles of 95°C for 30 s, 56°C for 30 s, and 72°C for 90 s. To determine the presence of bla CMY genes, primers ampC1 (5′-ATGATGAAAAAATCGTTATGC-3′) and ampC2 (5′-TTGCAGCTTTTCAAGAATGCGC-3′) were used (Winokur et al., 2001).
Plasmid purification and characterization
Plasmids were purified using the QiaFilter Midi kit (Qiagen Inc.), following a modified manufacturer's protocol (Folster et al., 2010). Electroporation of each plasmid into E. coli DH10B Electromax competent cells (Invitrogen, Carlsbad, CA) was performed as previously described (Folster et al., 2010). Cells were plated on LB agar plates containing 100 mg/L of ampicillin or 4 mg/L ceftriaxone (Sigma-Aldrich, St. Louis, MO). All transformants were confirmed as bla
CMY positive by PCR analysis using primers ampC1 and ampC2. DNA templates for PCR from transformants were prepared as described above. Plasmid PCR-based replicon typing (PBRT) was performed as previously described (Carattoli et al., 2005) on the transformants. Plasmid multilocus sequence typing (pMLST) was performed on IncI1 plasmids as previously described (Garcia-Fernandez et al., 2008). Sequencing was performed using Big Dye version 3.1 (Applied Biosystems, Foster City, CA) and sequence reactions were cleaned with Centri-sep plates (Princeton Separations, Adelphia, NJ). The reactions were electrophoresed through POP-7 polymer (Applied Biosystems) on a 3730 DNA Analyzer (Applied Biosystems) equipped with a 48-capillary, 50-cm array. Sequence analysis was performed using Lasergene 8 software (DNASTAR Inc., Madison, WI). Sequences were submitted to the pMLST web page (
Pulsed-field gel electrophoresis (PFGE)
Two-enzyme (XbaI and BlnI) PFGE was performed according to the CDC PulseNet protocol (Ribot et al., 2006; Jackson et al., 2007). Isolates were grown overnight on trypticase soy agar with 5% defibrinated sheep blood (Becton Dickinson Biosciences). Bacterial cell concentration was adjusted by diluting with sterile cell suspension buffer (100 mM Tris, 100 mM EDTA, pH 8.0) to a turbidity measurement of 0.48–0.52 (Dade Microscan Turbidity Meter). Agarose-embedded cells were lysed by proteinase K treatment and extensively washed. Agarose plugs containing genomic DNA were digested with 50 U of XbaI and BlnI restriction enzymes (New England Biolabs, Ipswich, MA) and incubated at 37°C for 2 h. The fragments were then separated by PFGE using a CHEF Mapper (Bio-Rad Laboratories) with the following conditions and reagents: 1% SeaKem Gold agarose in 0.5% Tris-borate-EDTA buffer, voltage at 6 V/cm, run time at 18 h with switch times ranging from 2.16 to 63.8 s, temperature at 14°C. Salmonella enterica serovar Braenderup H9812 was used as a molecular reference marker. PFGE profiles generated were submitted to the PulseNet national database administered by CDC (NARMS-FDA and NARMS-CDC) or USDA VetNet (NARMS-USDA). Gel images were captured using the GelDoc XR system (Bio-Rad Laboratories) and Quantity One 1-D analysis software (Bio-Rad Laboratories). Pattern analysis and unweighted-pair group method with arithmetic mean dendrogram generation were performed using BioNumerics software (Applied Maths, Saint-Martens-Latem, Belgium) with the Dice coefficient and tolerance of 1.5%.
Results
Identification of bla CMY-positive Salmonella Typhimurium isolates
NARMS received and performed antimicrobial susceptibility testing on 581 isolates of Salmonella Typhimurium from food animals, retail meat, and humans in 2008. Of these, 70 (12.0%) displayed resistance to ceftriaxone and amoxicillin–clavulanic acid, suggesting the presence of a bla CMY gene. Of the 70 isolates, 34 (48.6%) were from retail meat—specifically, chicken breasts. Twenty-three isolates (23/70 [32.9%]) were from food animal samples; 17/23 (73.9%) were from chickens and 6/23 (26.1%) were from cattle. Thirteen isolates were recovered from clinically ill humans: 2/13 (15.4%) were from male patients and 11/13 (84.6%) were from females. The median age was 29.9 years, with a range of <1 year to 77 years. PCR analysis confirmed that all 70 resistant isolates carried a bla CMY gene. Besides resistance to ceftriaxone and amoxicillin/clavulanic acid, all 70 isolates were resistant to the additional β-lactams tested (ampicillin, cefoxitin, and ceftiofur) (Fig. 1). The most common additional resistance observed among the isolates was to sulfisoxazole (n=68), tetracycline (n=64), streptomycin (n=24), and chloramphenicol (n=14). Resistance to kanamycin (n=9) and gentamicin (n=3) was less common and resistance to amikacin, ciprofloxacin, nalidixic acid, and sulfamethoxazole/trimethoprim was not observed.
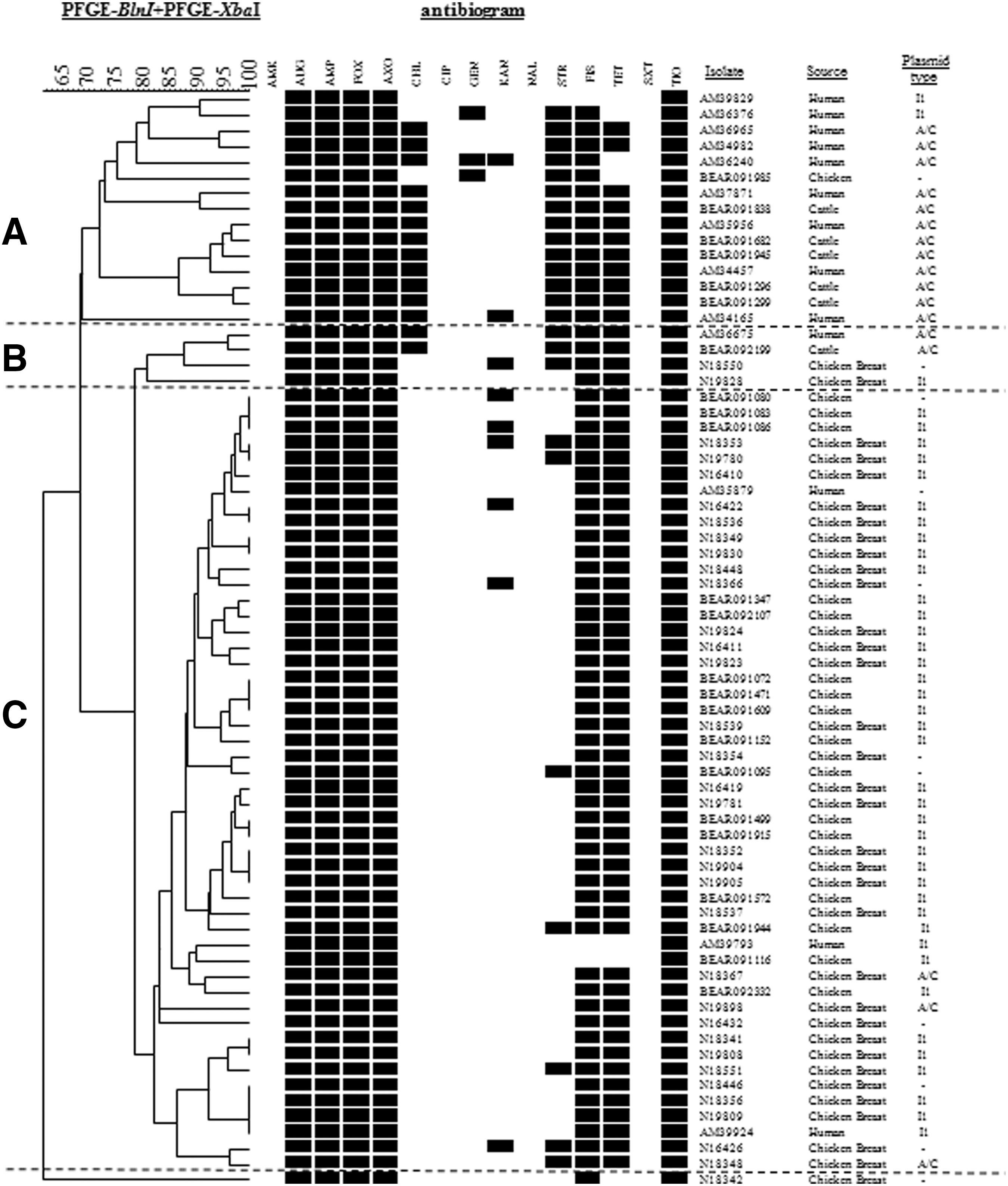
Pulsed-field gel electrophoresis (PFGE) patterns of bla CMY-positive Salmonella enterica Typhimurium isolated from food animals, retail meat, and humans in the United States in 2008. Dendrogram of percent genetic similarity by PFGE was generated using BioNumerics software (Applied Maths, Saint-Martens-Latem, Belgium) based on XbaI and BlnI restriction digestion. Pattern analysis and unweighted-pair group method with arithmetic mean dendrogram generation were performed using BioNumerics with the Dice coefficient and tolerance of 1.5%. Percent similarity is located above the dendrogram. The antibiogram displays the antimicrobial resistance profile of the isolates; a black box indicates resistance to that antimicrobial. AMK, amikacin; AUG, amoxicillin/clavulanic acid; AMP, ampicillin; FOX, cefoxitin; AXO, ceftriaxone; CHL, chloramphenicol; CIP, ciprofloxacin; GEN, gentamicin; KAN, kanamycin; NAL, nalidixic acid; STR, streptomycin; FIS, sulfisoxazole; TET, tetracycline; SXT, trimethoprim-sulfamethoxazole; TIO, ceftiofur. Isolate number, source, and plasmid incompatibility type are listed to the right of the antibiogram. The “-” symbol indicates that no bla CMY plasmid was found. The isolates are groups into three clusters, labeled A, B, and C.
Characterization of the bla CMY plasmids
To determine whether the bla CMY genes were located on a plasmid or the chromosome, plasmid DNA preparations were used to transform competent E. coli by electroporation. PCR analysis showed successful transfer of bla CMY genes to E. coli for 59/70 (84.3%) from the ESC-resistant Typhimurium isolates, suggesting that 11 isolates likely carried bla CMY chromosomally. PBRT performed on the transformants revealed that all 59 bla CMY-plasmid positive Typhimurium isolates carried the bla CMY gene on one of two plasmid types; 42/59 (71.2%) had IncI1- and 17/59 (28.8%) had IncA/C-bla CMY plasmids (Fig. 1). Among the 51 chicken/chicken breast isolates, 38 had IncI1-bla CMY plasmids, 3 had IncA/C-bla CMY plasmids, and 10 isolates had no bla CMY plasmids. Among the six cattle isolates, all had IncA/C-bla CMY plasmids. Among the 13 isolates from humans, 4 isolates had IncI1-bla CMY plasmids, 8 isolates had IncA/C-bla CMY plasmids, and 1 had no bla CMY plasmid. IncI1 plasmids were compared using pMLST (Garcia-Fernandez et al., 2008). All 42 IncI1-bla CMY plasmids were sequence type 12 or very closely related. Three isolates had plasmids with an identical point mutation in the pilL allele (G56–A56) while two other isolates had plasmids with two identical point mutations in the sogS allele (A175–G175 and T177–A177). All five of these isolates were isolated from chicken breasts.
Determining similarity by PFGE of the bla CMY-positive isolates
Two-enzyme PFGE was used to evaluate the genetic relatedness of the bla CMY-positive Typhimurium isolates from different sources (Fig. 1). Of the 70 isolates, 56 (80%) had unique two-enzyme patterns, suggesting very little clonal spread. There were nine groups of isolates with indistinguishable patterns, and the largest group contained four isolates. The isolates grouped into three main clusters (labeled A, B, and C) and one outlier (bottom of dendrogram). The largest group, cluster C, contained closely related (>75% with two enzymes) isolates from poultry sources (chicken breast [n=31] and chickens [n=16]) and humans (n=3). None of these isolates displayed resistance to chloramphenicol. Isolates in cluster C primarily contained IncI1-bla CMY plasmids (n=39/50; 78%) or no bla CMY plasmids (8/50; 16%). Only three isolates contained IncA/C-bla CMY plasmids. Cluster A was the next-largest cluster with 15 isolates. Although not as related (<75%) as cluster C, cluster A contained almost all of the isolates with chloramphenicol resistance (12/14; 85.7%) and most of the isolates with streptomycin resistance (14/24; 58.3%), meeting the definition of multi-drug resistant (MDR)-AmpC (Harbottle et al., 2006). Isolates in cluster A primarily contained isolates of either human (n=9) or cattle (n=5) sources and only a single poultry isolate. Cluster A also consisted primarily of isolates with IncA/C-bla CMY plasmids (12/15; 80%), with only two IncI1-bla CMY containing isolates and a single isolate with no bla CMY plasmid. Cluster B contains only four isolates and is mixed with respect to antimicrobial resistance patterns (two isolates displayed chloramphenicol resistance), isolate source (two isolates from chicken breast, one cattle, and one human source) and bla CMY plasmid type (two IncA/C-bla CMY, one IncI1-bla CMY, and one without a bla CMY plasmid).
Discussion
Laboratory-based disease surveillance and foodborne outbreak detection and investigation require high-quality epidemiological data and detailed agent information. With the evolution of new strain typing techniques, investigators have sought to combine various tools to provide more specific agent information in an effort to improve the surveillance and investigative processes. We examined the value of combining phenotypic data on antimicrobial resistance and serotype with genetic data represented by PFGE patterns and differences in plasmid content.
IncI1 plasmids are a narrow-host-range plasmid type and are limited to enteric bacteria (Johnson et al., 2011). They are commonly associated with Salmonella and E. coli from avian and porcine sources, and are more frequent among pathogenic than commensal E. coli strains found among avian and human sources in the United States (Johnson et al., 2007). IncI1 plasmids commonly carry genes conferring ESC resistance, including bla CMY and bla CTX-M. Previous studies have identified IncI1 plasmids as the primary plasmid type carrying bla CMY among humans, with Salmonella serotypes commonly associated with poultry, and IncI1 is the most prominent bla CMY plasmid type among serotype Heidelberg isolates from humans and poultry sources (Folster et al., 2010, 2011).
IncA/C plasmids have a broad host range and have been isolated from diverse groups of Proteobacteria found in the environment, animals, and humans (Lang et al., 2012). IncA/C plasmids are commonly large, multidrug resistant, and have been identified among isolates from food animals including beef, chicken, turkey, and pork, suggesting that they may be responsible for the spread of MDR from food animals to humans (Mulvey et al., 2009). IncA/C plasmids are one of the most frequent plasmid types carrying bla CMY in the United States (Giles et al., 2004). IncA/C plasmids are the most common plasmid type carrying bla CMY among humans with Salmonella serotypes usually associated with cattle and beef sources, including Newport and Dublin (Folster et al., 2010). A study of bla CMY plasmids from E. coli and Salmonella in Canada found that bacteria from cattle and beef all had IncA/C plasmids (Martin et al., 2012).
In this study, we identified and characterized 70 ceftriaxone and amoxicillin/clavulanic acid–resistant Salmonella Typhimurium isolates. All of the isolates contained a bla CMY gene, and over 80% were plasmid encoded. We identified two bla CMY plasmid types, IncI1 and IncA/C, with IncI1 comprising >70% of bla CMY plasmids identified. When we compared plasmid type to the source of each nonclinical isolate, we found that nearly all of the isolates with bla CMY plasmids from chicken or chicken products had IncI1-bla CMY plasmids (92.5%), while all of the cattle isolates had IncA/C-bla CMY plasmids. This shows a correlation between the animal source of Typhimurium isolates and the replicon type of bla CMY plasmid they carry. However, a larger study set of isolates over several years is needed to confirm this observation. Clinical isolates from humans with bla CMY plasmids had both IncA/C-bla CMY (n=8) and IncI1-bla CMY (n=4) plasmids; however, no information was available regarding the source of infection of these routine surveillance isolates.
When we compared plasmid type, source, and antimicrobial resistance patterns, we found that all of the IncA/C-bla CMY isolates from cattle and humans were resistant to chloramphenicol. Chloramphenicol resistance is commonly conferred by IncA/C plasmids (Lindsey et al., 2011). However, the three IncA/C-bla CMY isolates from chicken breasts were all chloramphenicol susceptible, and none of the IncI1-bla CMY–positive isolates from chicken or chicken breasts were chloramphenicol resistant. This suggests that chloramphenicol resistance, when observed along with the bla CMY-mediated resistance phenotype, may point to a possible cattle/beef source.
PFGE analysis showed a significant amount of genetic variation among all 70 isolates, but the isolates grouped into 3 main clusters (A, B, and C). When we compared this to isolate source, bla CMY plasmid type, and chloramphenicol resistance, we again found a strong correlation. Cluster C, the largest cluster with 50 isolates, contained 92.2% of the poultry isolates and no cattle isolates, 91.6% of the IncI1-bla CMY plasmids, and no chloramphenicol resistant isolates. In contrast, cluster A, containing 15 isolates, had 5 of 6 isolates from cattle, 70.6% (12/17) of the IncA/C-bla CMY plasmids, and 85.7% (12/14) of the chloramphenicol-resistant isolates. If we further divide cluster A, the bottom group (containing seven isolates) contains isolates with an identical resistance profile and plasmid type (IncA/C) but isolates from both cattle (n=4) and humans (n=3), suggesting that the human isolates were likely acquired from a beef source.
Interestingly, even though Salmonella is rarely isolated from ground beef and none of the isolates in this study were isolated from ground beef, the majority of isolates from humans had IncA/C-bla CMY plasmids, which we interpret to indicate that humans were more likely to acquire bla CMY isolates with plasmids that resemble those from cattle sources than poultry sources. This suggests that human infections with bla CMY-positive Typhimurium isolates may be from another beef source that is not being sampled or that current sampling and/or isolation methods are not detecting Salmonella in ground beef. The latter may be due to processing events specific for ground beef that could result in a temporary decrease in Salmonella numbers at the time of sampling (Harris et al., 2012). It is also possible that humans are acquiring bla CMY-positive Salmonella Typhimurium from unsampled nonmeat sources, including vegetables.
Conclusion
Although the numbers of isolates in the study were small, especially from cattle, we did find associations between bla CMY plasmid type and food animal source (IncA/C and cattle, IncI1 and poultry). This work may be useful in generating hypotheses about the sources for foodborne outbreaks and sporadic illness caused by Salmonella. However, the resistance plasmid type needs to be examined along with careful consideration of all of the available information. This is particularly important for some serotypes such as Typhimurium, which has great genetic diversity and a wide range of hosts. Further work is necessary to determine whether these correlations occur with other Salmonella serotypes, other enteric bacteria, and additional antimicrobial resistance plasmids. Analysis of a larger set of isolates over a longer period of time is needed to determine how best to combine plasmid, PFGE, serotype, and susceptibility data to effectively guide investigations of foodborne disease.
Footnotes
Acknowledgments
We thank the NARMS participating public health laboratories for submitting the isolates, Anne Whitney for DNA sequencing, and Alessandra Carattoli for the plasmid incompatibility typing control strains. This work was partially supported by an interagency agreement between CDC, USDA, and the FDA Center for Veterinary Medicine.
Disclosure Statement
No competing financial interests exist.