
Abstract
Pomegranate rind has been reported to inhibit several foodborne pathogens, and its antimicrobial activity has been attributed mainly to its tannin fraction. This study aimed to investigate the antimicrobial activity of the tannin-rich fraction from pomegranate rind (TFPR) against Listeria monocytogenes and its mechanism of action. The tannin-related components of TFPR were analyzed by high-performance liquid chromatography and liquid chromatography–mass spectrometry, and the minimum inhibitory concentration (MIC) of TFPR was determined using the agar dilution method. Extracellular potassium concentration, the release of cell constituents, intra- and extracellular ATP concentrations, membrane potential, and intracellular pH (pHin) were measured to elucidate a possible antibacterial mechanism. Punicalagin (64.2%, g/g) and ellagic acid (3.1%, g/g) were detected in TFPR, and the MICs of TFPR were determined to be 1.25–5.0 mg/mL for different L. monocytogenes strains. Treatment with TFPR induced a decrease of the intracellular ATP concentration, an increase of the extracellular concentrations of potassium and ATP, and the release of cell constituents. A reduction of pHin and cell membrane hyperpolarization were observed after treatment. Electron microscopic observations showed that the cell membrane structures of L. monocytogenes were apparently impaired by TFPR. It is concluded that TFPR could destroy the integrity of the cell membrane of L. monocytogenes, leading to a loss of cell homeostasis. These findings indicate that TFPR has the potential to be used as a food preservative in order to control L. monocytogenes contamination in food and reduce the risk of listeriosis.
Introduction
L
Currently, synthetic preservatives are commonly used to protect food from microbial spoilage or contamination. However, due to the increasing consumers' demand for natural foods, natural preservatives derived from plants, animals, or microflora have drawn extensive attention. In recent years, plant materials have been considered as important sources of natural preservatives and have been extensively tested for antimicrobial activity (Cowan, 1999; Aqil et al., 2005). It has been demonstrated that a variety of phytochemicals exhibited antimicrobial activity against pathogenic bacteria such as Escherichia coli, Staphylococcus aureus, and Pseudomonas aeruginosa (Ankri and Mirelman, 1999; Aqil et al., 2005; Qiu et al., 2010).
Pomegranate (Punica granatum) fruit has been extensively used in several countries for treatment of various diseases, including dysentery, helminthiasis, diarrhea, hemorrhage, and acidosis (Voravuthikunchai et al., 2005; Sánchez-Lamar et al., 2008; Al-Zoreky, 2009). Studies have shown that pomegranate peel is effective in inhibiting S. aureus (Braga et al., 2005), Salmonella (Al-Zoreky, 2009; Choi et al., 2011), L. monocytogenes (Hayrapetyan et al., 2012), E. coli (Al-Zoreky, 2009), and fungi (Glazer et al., 2012) in vitro. Its antimicrobial activity has been largely attributed to its hydrolyzable ellagitannins, among other polyphenols, which are the main phytochemicals of pomegranate rind (Voravuthikunchai et al., 2004, 2005; Glazer et al., 2012).
However, the majority of previous studies failed to report specific components included in the crude extracts obtained from pomegranate rind (Braga et al., 2005; Al-Zoreky, 2009; Choi et al., 2011; Hayrapetyan et al., 2012). Also, the mechanism of antimicrobial action has rarely been elucidated. The aims of this study therefore were to investigate the antilisterial effect of tannin-rich fraction from pomegranate rind (TFPR) and to explore its possible antimicrobial mechanism with a focus on the cell membrane.
Materials and Methods
Bacterial strains and culture conditions
L. monocytogenes CMCC54004 (serotype: 1/2a) was obtained from the National Center for Medical Culture Collections, Beijing, China. The strain is a L. monocytogenes–type strain and was commonly used as a reference strain in research related to L. monocytogenes in China. Six additional L. monocytogenes strains of different serotypes (Table 1) were taken from our laboratory collection. They were originally isolated from various food products in Shaanxi province, China. All isolates were used in minimum inhibitory concentration (MIC) assay, and only L. monocytogenes CMCC54004 was used in antimicrobial mechanism study. Before each experiment, bacterial cells were grown in LB broth (Beijing Land Bridge Technology Co., Ltd., Beijing, China) at 37°C for 18 h and harvested by centrifugation at 1900×g for 10 min (Model 5804R; Eppendorf, Hamburg, Germany). The cell pellets were resuspended in 0.9% (wt/vol) saline solution to obtain working cultures with an optical density at 600 nm of 0.5 (approximately 108 CFU/mL).
MICs of these two components were only tested on L. monocytogenes CMCC54004.
Preparation and analysis of TFPR
Pomegranate (a local variety named Yicheng ruanzi) was purchased from Lintong District, Xi'an, Shaanxi province in China in October 2011. The fruits were washed with tap water and gently brushed in order to remove any dirt, and the rinds were then separated from the seeds and juice. The pomegranate rind was dried at 50°C for 48 h and then ground into powder using a mechanical grinder (Model FW135, Taisite, Tianjin, China). Approximately 200 g of the powder were extracted ultrasonically (40°C, 90 W) in 1000 mL of 60% (vol/vol) acetone for 15 min. The acetone was removed under reduced pressure in a rotary evaporator at 40°C and then the concentrate was stored at 4°C prior to use. Purification of the concentrate was achieved by following the method described previously by Seeram et al. (2005), resulting in the freeze-dried form of tannins to yield TFPR. The dried TFPR was stored at 4°C prior to use.
The high-performance liquid chromatography (HPLC) analysis was performed on a Waters 600E
Determination of antibacterial activity parameters
MIC
The agar dilution method of the European Committee for Antimicrobial Susceptibility Testing (EUCAST, 2000) was adopted to determine MIC of TFPR. Streptomycin was used as control. The final concentrations of TFPR utilized in the solidified LB agar poured in Petri dishes were 0, 1.25, 2.5, 5, and 10 mg/mL, and that of streptomycin was 10 μg/mL. After the media were solidified, 100 μL (approximately 106 CFU/mL) of the bacteria culture were spread on the agar plate. After drying for 5 min, the plates were inverted and incubated at 37°C for 24 h. The MIC was defined as the lowest concentration of TFPR resulting in no growth of L. monocytogenes on spread agar plates, compared to the control.
Extracellular amounts of potassium
L. monocytogenes cell suspensions were prepared as previously described. Then different amounts of TFPR were added to the suspension so that the final TFPR concentrations were 0, MIC, and 2MIC, respectively. Every 30 min, 5 mL of bacterial suspension were sampled and centrifuged. The extracellular potassium concentrations were measured in the supernatant by using a SmartSpec™ Plus Spectrophotometer (Bio-Rad, Hercules, CA) and a potassium assay kit (Nanjing Jiancheng Bioengineering Institute, Nanjing, China) following the manufacturer's protocols.
Cell constituents' leakage
The leakage of cell constituents was investigated according to a modified method by Rhayour et al. (2003). Twenty milliliters of cell suspension were incubated at 37°C with agitation (130 rpm) in the absence or presence (at MIC) of TFPR. Then, 1 mL of sample was taken every 15 min and centrifuged for 5 min at 1900×g. The supernatant was diluted and the concentration of the released cell constituents was determined at 260 nm using a spectrophotometer (Nano-200; Hangzhou Allsheng Instruments Co., Ltd., Hangzhou, China).
Intra- and extracellular ATP concentrations
TFPR was added into the bacterial cell suspensions until the final concentrations of TFPR were 0 (control), MIC, and 2MIC. The suspensions for each treatment were incubated at 37°C for 30 min, then centrifuged (10 min at 1900×g), and maintained on ice to prevent ATP loss until measurement. The extra- and intracellular ATP concentrations were measured using an ATP assay kit (Nanjing Jiancheng Bioengineering Institute). The ATP concentration of the supernatant, which represents the extracellular concentration, was determined spectrometrically following the instructions contained in the kit. In order to determine the intracellular ATP concentrations, the cell pellets were washed with 0.9% saline solution. Following centrifugation, the supernatant was removed and the cell pellets were resuspended in 100 μL of 0.9% saline solution. After boiling for 10 min to disrupt the microorganisms' cell wall, the intracellular ATP concentrations were measured as described for extracellular ATP. For comparison, the intracellular and extracellular ATPs of cells with no TFPR treatment were used as controls.
Membrane potential
A slight modification of the method described by Sanchez et al. (2010) was followed to determine changes of membrane potential caused by TFPR. In 25 mL of bacterial suspension, 2.5 μL of membrane potential-sensitive fluorescent probe bis-(1,3-dibutylbarbituric acid) trimethine oxonol (DiBAC4[3]; Molecular Probes, Eugene, OR) (1 mM) were added and mixed for 5 min at 25°C, followed by the addition of TFPR at four different concentrations (0, 1/2MIC, MIC, and 2MIC). At 3 h, fluorescence was measured at the excitation and emission wavelengths of 492 and 515 nm, respectively, by a fluorescence spectrophotometer (RF-5301; Shimadzu, Kyoto, Japan). The excitation and emission slit widths were 3 and 5 nm, respectively. Background fluorescence resulting from the DiBAC4[3]”added to 0.9% saline solution was determined and the results were normalized.
Intracellular pH (pHin)
The pHin of L. monocytogenes was analyzed according to the spectrofluorometric method described by Breeuwer et al. (1996). The fluorescent intensities of three fluorescently labeled cell suspensions with TFPR at 0, MIC, and 2MIC, respectively, were measured every 30 min using an excitation wavelength of 490 nm and an emission wavelength of 520 nm by a fluorescence spectrophotometer (RF-5301, Shimadzu). The excitation and emission slit widths were 3 and 5 nm, respectively. The pHin of bacteria was determined from the fluorescence intensities at the pH-sensitive wavelength (490 nm).
Electron microscopy
Scanning electron microscopy and transmission electron microscopy (TEM) were employed to examine the cell damage caused by TFPR. Bacterial cell suspensions containing TFPR at three concentrations (0, MIC, and 2MIC) were incubated for 2 h at 37°C, and cells were collected and immobilized with 2.5% glutaraldehyde at 4°C for 10 h. The cells were then centrifuged and washed with phosphate-buffered saline (PBS), followed by dehydration in ethanol solution. After freeze drying, cells were fixed on scanning with Au–Pd and images were obtained using a Hitachi model S-4800 field emission scanning electron microscope (Hitachi, Tokyo, Japan).
L. monocytogenes cells for TEM observation were immobilized with 2.5% glutaraldehyde applied for 10 h followed by 1% osmic acid for 2 h. The cells were dehydrated in acetone solution after washing with PBS, and they were infiltrated and embedded with resin. Ultrathin sections were double stained with saturated lead citrate–uranyl acetate. The slices were visualized with a Hitachi model HT7700 TEM (Hitachi).
Statistical analyses
All the experiments were performed in triplicate, and two samples in each replicate were tested for each measurement. Duncan's multiple-range tests were used for statistical analysis by SPSS version 19.0 (IBM, New York, NY). Differences between means were considered significant when p≤0.05.
Results
Analysis of TFPR
Purification and subsequent analysis of TFPR revealed the presence of a large amount of punicalagin (64.2%, isomeric forms of α- and β-punicalagin together) and a small amount of ellagic acid (3.1%) as determined by the standard curves with commercial standards (Fig. 1) and confirmed also by LC-MS/MS (data not shown). Other peaks could not be identified due to the lack of corresponding standards.
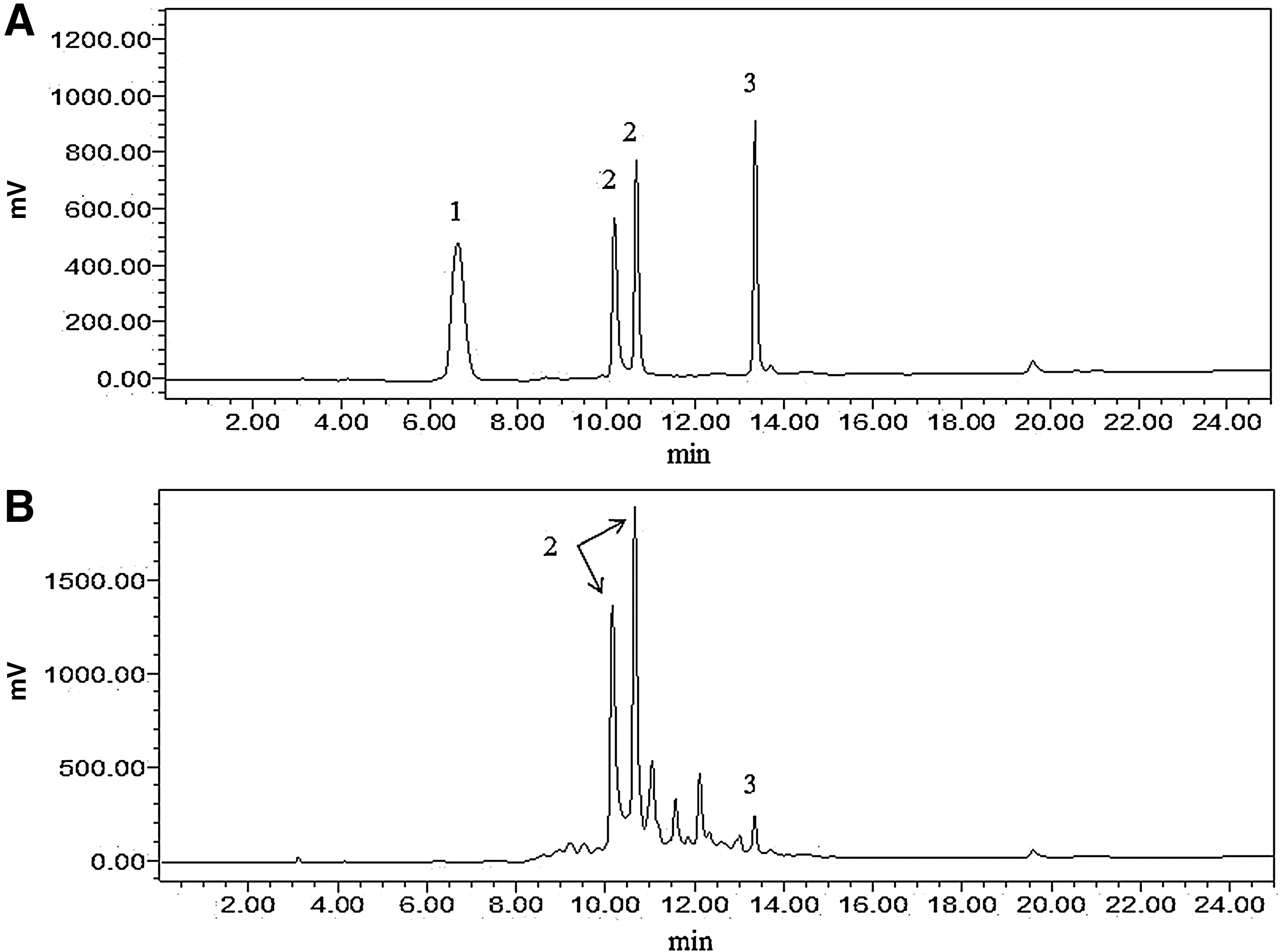
High-performance liquid chromatography analysis of tannin-rich fraction from pomegranate rind (TFPR). The chromatograms of
Antibacterial activity parameters
The MICs of TFPR against seven L. monocytogenes strains were determined and ranged from 1.25 to 5 mg/mL (Table 1). The MICs of TFPR, punicalagin, and ellagic acid against the L. monocytogenes CMCC54004 strain were found to be 5, 2.5, and >10 mg/mL, respectively.
The extracellular potassium concentrations increased after treatment with different concentrations of TFPR (Fig. 2). L. monocytogenes treated with TFPR at MIC (5 mg/mL) and 2MIC (10 mg/mL) showed a significant increase (p≤0.05) in the secreted potassium concentration compared to the control at any given time (30, 60, and 90 min). Higher concentrations of extracellular potassium were observed after treatment with TFPR at 2MIC than at MIC.
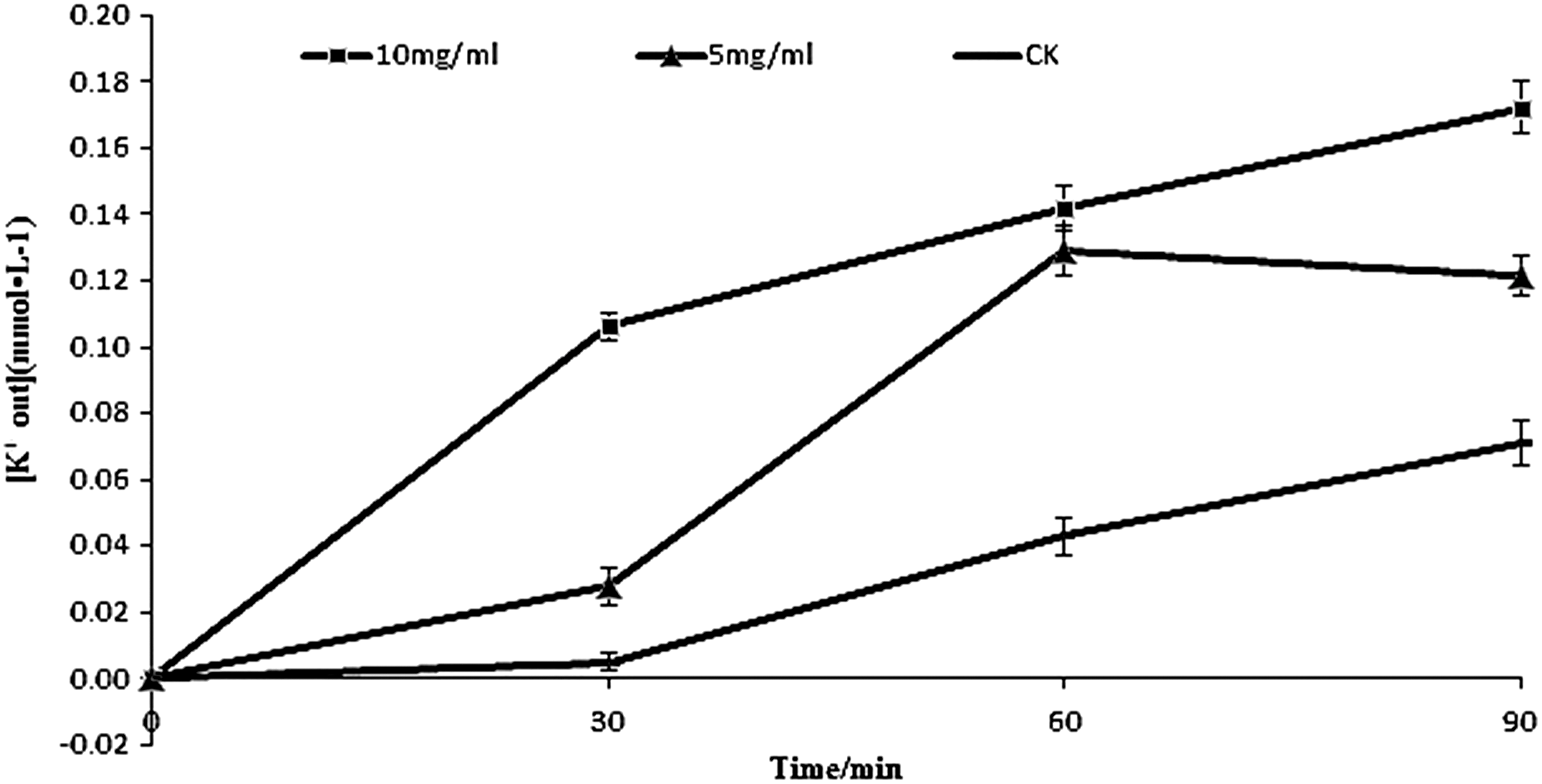
Effect of treatment with different concentrations of tannin-rich fraction from pomegranate rind on the extracellular amount of potassium for Listeria monocytogenes. Bars represent the standard deviation (n=3).
Starting from 15 min, a significant (p≤0.05) increase of the cell constituents' leakage was observed when L. monocytogenes cells were treated with TFPR at a concentration of 5 mg/mL compared to the control (data not shown).
The effect of TFPR on the intra- and extracellular ATP concentrations for L. monocytogenes cells is shown in Figure 3. After L. monocytogenes cells were treated with TFPR, the intracellular ATP concentrations decreased significantly along with an increase in the extracellular ATP concentration (p≤0.05). A significant difference (p≤0.05) of intracellular ATP concentration between treatments using two different concentrations of TFPR (MIC or 2MIC) was observed. However, no significant difference (p>0.05) of the extracellular ATP concentration was found between treatments using TFPR at the two concentrations.
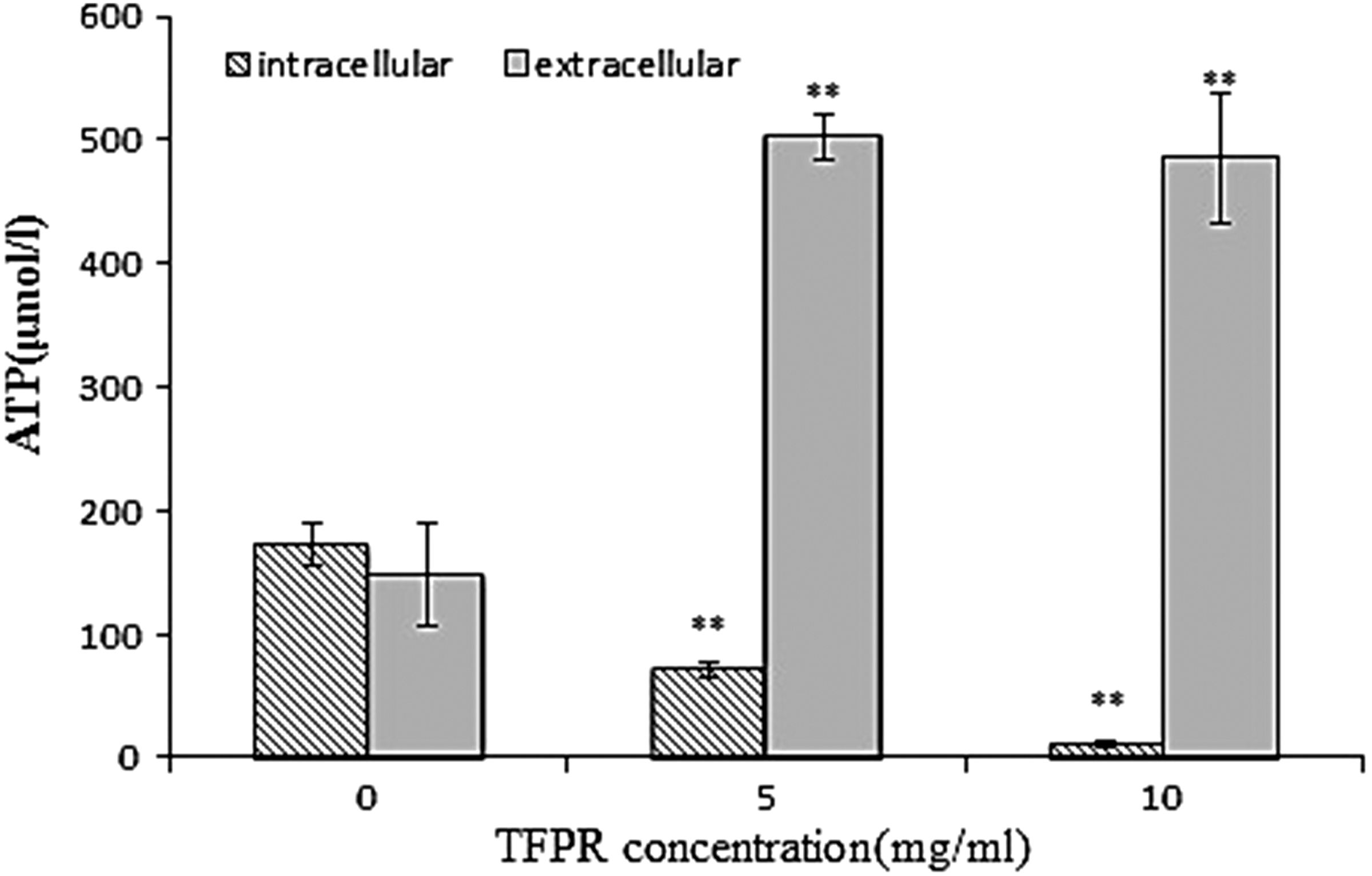
Effect of treatment with different concentrations of tannin-rich fraction from pomegranate rind (TFPR) on the intra- and extracellular ATP concentrations for Listeria monocytogenes. Bars represent the standard deviation (n=3). *p<0.05, **p<0.01.
As evidenced by a reduction in fluorescence, during monitoring of membrane potential, cells treated with different concentrations of TFPR displayed cell membrane hyperpolarization (data not shown). Increased concentration of TFPR caused a higher degree of hyperpolarization of the cells.
A clear change in pHin was observed after treatment with TFPR (Fig. 4). The pHin of L. monocytogenes exposed to the TFPR at the concentration of 2MIC (10 mg/mL) decreased more (p≤0.05) than that at MIC (5 mg/mL), although no significant reduction (p≤0.05) of pHin with time was observed for each treatment.
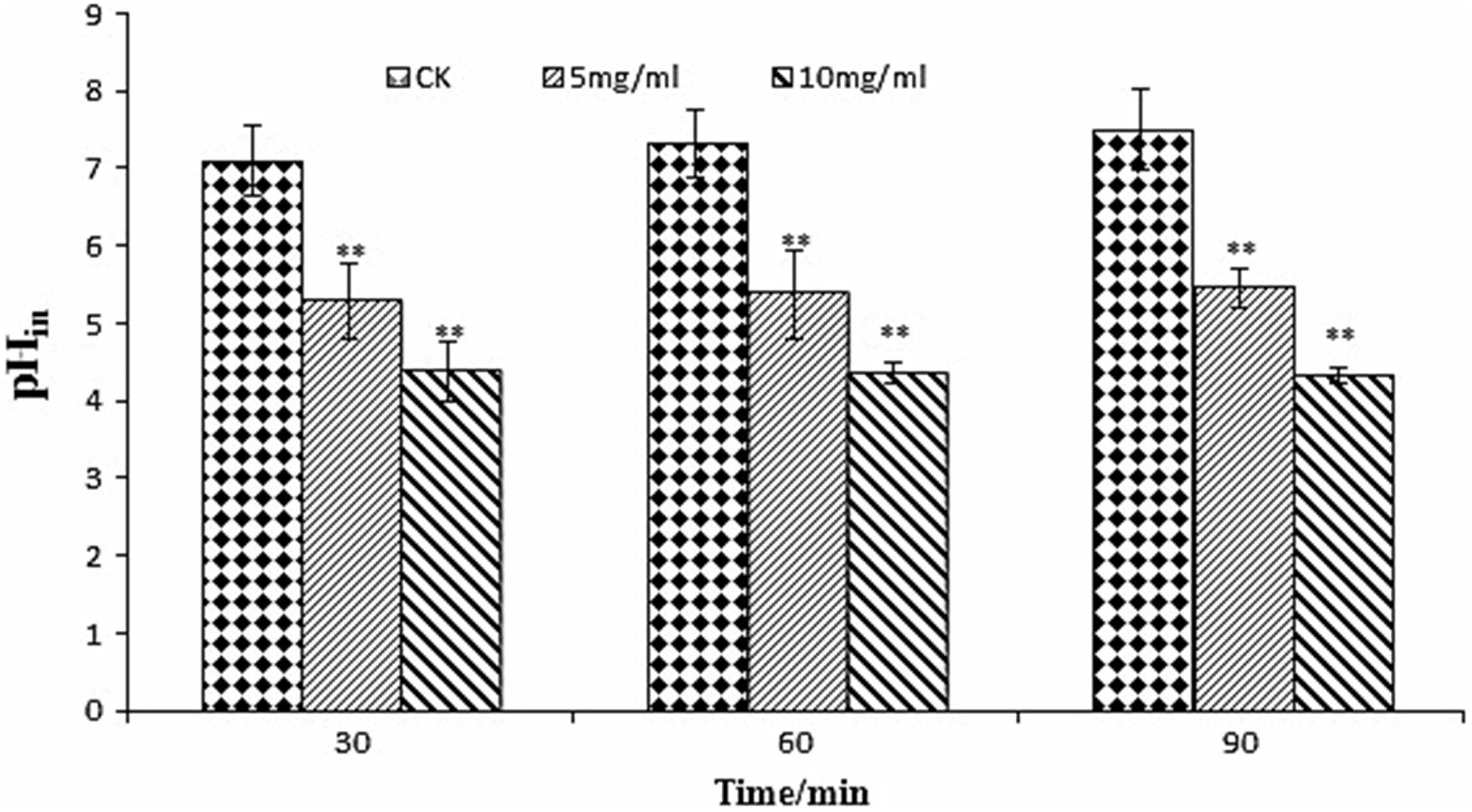
Effect of treatment with different concentrations of tannin-rich fraction from pomegranate rind on the intracellular pH (pHin) of Listeria monocytogenes. Bars represent the standard deviation (n=3). *p<0.05, **p<0.01. CK, control.
Electron microscopy observation
L. monocytogenes had a typical Gram-positive structure with striated membranes and clear cell–cell boundaries (Fig. 5A). The intracellular protoplasm had uniform distribution and high density (Fig. 5D). Cells treated with TFPR had damaged morphology with membranes disrupted (Fig. 5B–F). Serious plasmolysis was evident with depletion of cell content, confirming that the cell membrane structures of bacteria had been severely damaged by the TFPR.
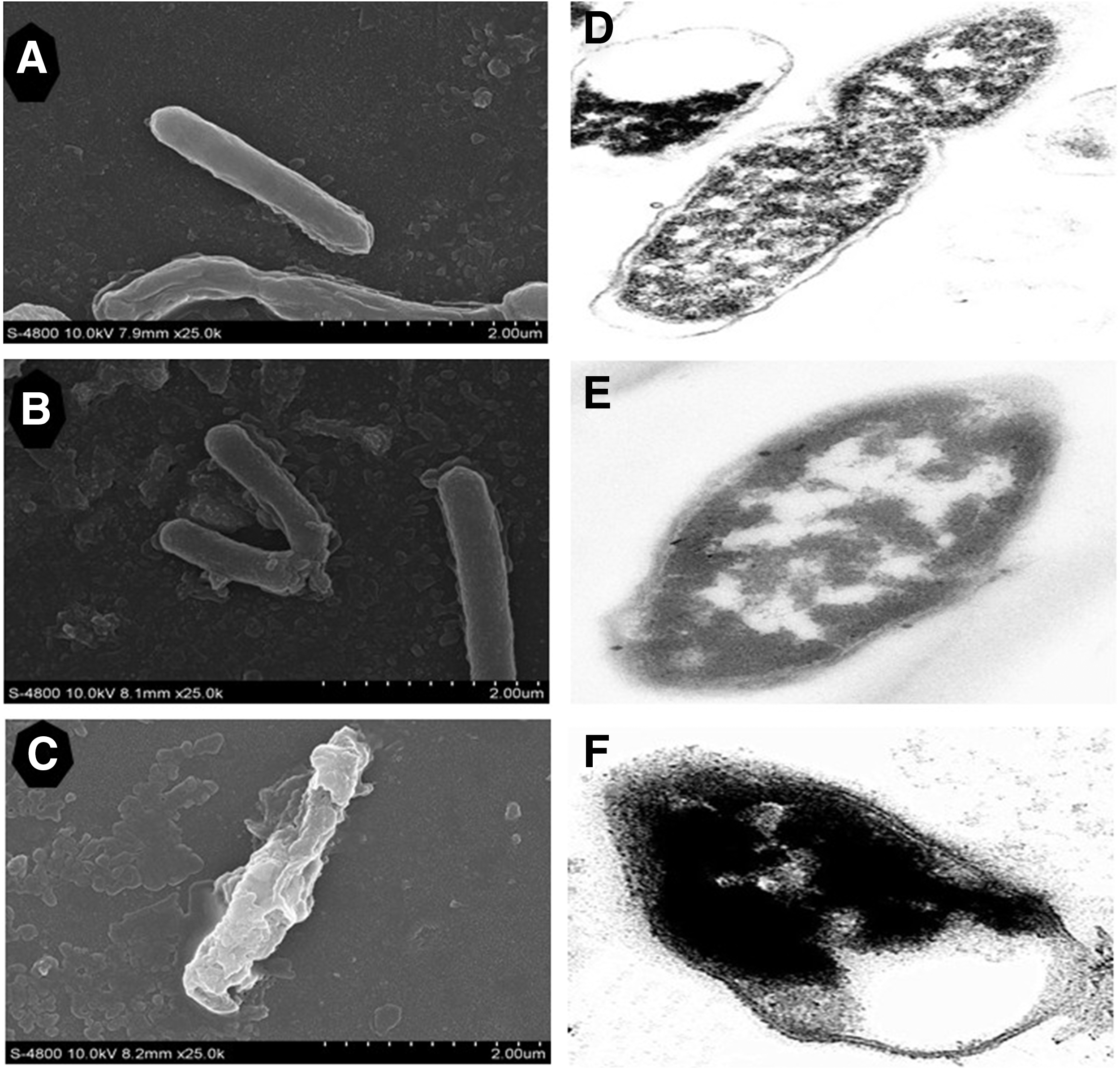
Effect of treatment with different concentrations of tannin-rich fraction from pomegranate rind (TFPR) on the cell morphology of Listeria monocytogenes.
Discussion
Pomegranate rind extracts have been reported to inhibit several foodborne pathogens, but most studies used crude extracts with unknown constituents and so the mechanism of antimicrobial action remained obscure. In this work, pomegranate rind was subjected to tannin-directed extraction and purification, and its antimicrobial activity against L. monocytogenes was investigated. In addition, there has been an effort to explore the antimicrobial mechanism of action by focusing on the cell membrane. It was demonstrated that TFPR could destroy the permeability of the cell membrane, leading to the leakage of potassium and cell constituents, along with a decrease of the intracellular ATP concentration and an increase of extracellular ATP concentration. Moreover, cell membrane hyperpolarization and a reduction of the pHin were found, and the damaged membranes of L. monocytogenes were also observed.
The loss of cytoplasmic constituents after TFPR treatment described herein was consistent with several studies, which reported that natural components can damage the membrane structure of microorganisms and promote the loss of cellular components, such as ions, proteins, and nucleic acids (Ultee et al., 1999; Rhayour et al., 2003; Turgis et al., 2009; Sanchez et al., 2010; Lv et al., 2011; Stojanović-Radić et al., 2012). Ultee et al. (1999) reported an increase of the permeability of the cytoplasmic membrane for potassium ions due to carvacrol. Lv et al. (2011) found that the release of cell constituents increased when E. coli, Saccharomyces cerevisiae, S. aureus, and Bacillus subtilis were treated with essential oils. All authors indicated that natural components might compromise the cell membrane integrity of microorganisms and cause leakage of cytoplasmic constituents.
TFPR caused a decrease of intracellular ATP concentration, while inducing an increase of extracellular ATP concentration. This was in accordance with Sanchez et al. (2010), who showed that extracts from four types of plants caused a decrease in the cytoplasmic ATP concentration of Vibrio cholerae. Helander et al. (1998) found that carvacrol and thymol decreased the intracellular ATP pool of E. coli and at the same time increased extracellular ATP. Lee et al. (2002) observed that linolenic acid with or without monoglyceride changed the intra- and extracellular ATP concentrations of B. cereus and S. aureus. It is hypothesized that the membrane of the L. monocytogenes was impaired by TFPR, and the intracellular ATP leaked through the damaged membrane. Furthermore, the diminished ATP concentration inside the cells can be correlated with possible hydrolysis by the proton-pumping ATPase (Ultee et al., 1999).
Hyperpolarization (more negative charge inside the cell) of L. monocytogenes membranes was observed after treatment with TFPR. Sanchez et al. (2010) also reported that extracts from three edible and medicinal plants were able to disrupt the cell membranes of V. cholerae cells, causing cell membrane hyperpolarization. In addition to depolarization, hyperpolarization is also an important type of membrane damage (Yuroff et al., 2003), which is now considered to happen due to a pH change or by increasing movement of ions, specifically K+, which diffuses out of the cell membrane through K+ channels and affects cellular homeostasis (Bot and Prodan, 2009).
Different bacteria can exhibit a wide range of pHin values, from 5.6 to 9 (Breeuwer et al., 1996), and even a slight variation of this parameter can be indicative of changes taking place in the bacterial cell (Turgis et al., 2009). The pHin is vitally important for the control of a great many cellular processes, such as DNA transcription, enzyme activities, and protein synthesis. Many studies have proved that plant materials can lower pHin. Lambert et al. (2001) reported that the pHin of P. aeruginosa and S. aureus was dissipated by oregano essential oil. Turgis et al. (2009) demonstrated that mustard essential oil at the concentration of MIC lowered the pHin in E. coli O157:H7, L. monocytogenes, and Salmonella typhi. Consequently, it is proposed that TFPR interacts with the bacterial membrane and changes the permeability for ions like H+, leading to a disrupted pH homeostasis.
The membrane structure of L. monocytogenes had been severely impaired by the addition of TFPR, which is consistent with the leakage of K+ and cell constituents, as well as with the increase of extracellular ATP concentration. Our findings are in good agreement with the behavior of other phenolic compounds used against microbial growth (Lv et al., 2011; Yao et al., 2012). An incomplete and deformed shape was observed when E. coli, B. subtilis, S. aureus, and S. cerevisiae were treated with essential oil combinations, accompanied by the lack of their cell walls (Lv et al., 2011). P. fluorescens and P. aeruginosa treated with polymethoxylated flavone presented damaged morphology with disrupted membranes and depleted cell contents (Yao et al., 2012).
In summary, TFPR exhibited antimicrobial activity against L. monocytogenes, and it may exert its antilisterial effect by damaging the cell membrane, causing an increase in membrane permeability, changes in extra- and intracellular ATP concentrations and cell morphology, cell membrane hyperpolarization, and a decrease in cytoplasmic pH. Other possible mechanisms of actions such as inhibition of several enzymes or protein synthesis need further research in the future. TFPR could be considered as a promising alternative food preservative for the food industry. Nonetheless, before its application in food, problems such as large-scale extraction and purification need to be resolved, and its toxicity in humans should be assessed.
Footnotes
Acknowledgments
This work was supported in part by the Twelve-Five Science and Technology Support Porgram (No. 2012BAK17B06, 2012BAH30F03), National Natural Science Foundation of China (No. 31101347), Science and Technology Development Plan Program of Shaanxi Province (No. 2013KJXX-16), and Special Funds for Talents from Northwest A&F University (No. Z111021002).
Disclosure Statement
No competing financial interests exist.