
Abstract
To evaluate the efficacy of a type-III secreted proteins vaccine and a Lactobacillus-acidophilus–based direct-fed microbial (DFM) for controlling Escherichia coli O157:H7, cattle (n=864) were allocated to the following groups: DFM, finishing diets containing 109 colony-forming units (CFU)/animal/day L. acidophilus and Propionibacterium freudenreichii; VAC, finishing diets and 2 mL intramuscular injection of vaccine at allocation and 28 days later; or CON, finishing diets only. Cattle within replicates were stratified by initial levels of E. coli O157:H7 and randomized to experimental groups, with 30 pens allocated on June 15, 2011 (AS1), 18 pens allocated on June 28, 2011 (AS2), and 18 cattle per pen. Rectal fecal samples and perineal swabs were collected at 28-day intervals until shipment to slaughter (103–145 days on trial). Numbers of cattle with enumerable E. coli O157:H7 (≥1.6 CFU/g feces) were reduced in AS1 and AS2 by VAC (p=0.008), although interventions had no impact on numbers of E. coli O157:H7 shed. For AS1, VAC reduced prevalence of E. coli O157:H7 in feces (p=0.03) and perineal swabs (p=0.04) in the feeding period but not at shipment to slaughter. For AS2, prevalence of E. coli O157:H7 was not reduced in either feces or perineal swabs by VAC at any time. For AS1, DFM reduced prevalence of E. coli O157:H7 in perineal swabs (p=0.01) during the feeding period. For AS2, DFM increased E. coli O157:H7 detection in feces (p=0.03) and perineal swabs (p=0.01) at shipment to slaughter. Seventy-five percent of AS1 E. coli O157:H7 isolates had only stx1, while 87% of AS2 isolates had stx1 and stx2 genes. Of the two interventions, VAC shows the most potential for pre-harvest control of E. coli O157:H7, but due to variable efficacy of both DFM and VAC, additional product development is necessary to ensure more consistent pre-harvest control of E. coli O157:H7.
Introduction
C
Materials and Methods
Facilities and feed
The project was conducted in a small pen research facility (June–November 2011), using cattle housed in open air, dirt-floor pens with central feed alleys and porosity fencing. Feed and water were available ad libitum. Diets were standardized within replicate and included approximately 92% barley grain, 5% forage, 2% supplement, and 1% corn dried distiller grains (cDDG) on a dry-matter basis to meet beef cattle nutrient requirements (NRC, 2000). Diets were blended in truck-mounted mixer boxes equipped with electronic load cells, and contained monensin (Elanco Animal Health, Guelph, ON), tylosin (Elanco Animal Health), and melengestrol acetate (Pfizer Animal Health, heifer diets only).
The DFM was shipped on ice and frozen (-20°C) until use. The DFM was measured daily to target 1 g/animal/d and added to diets using cDDG carrier. Pens within a replicate were fed equal amounts of cDDG, and VAC and CON pens were fed prior to DFM pens to avoid cross-contamination. Residual DFM diet in feed trucks was removed by feeding nonstudy cattle.
Cattle, interventions
Cattle were from two feedlots; with most allocation set 1 (AS1) cattle from Feedlot A and most allocation set 2 (AS2) from Feedlot B. Using new gloves for each animal, feces (3–5 g) were rectally collected at feedlots A and B for detection and enumeration of E. coli O157:H7. Cattle were blocked by weight to facilitate marketing and stratified by E. coli O157:H7 level (Negative, no E. coli O157:H7 detected; Lower, insufficient for enumeration; Higher, ≥1.6 log colony-forming units (CFU)/g feces) and randomized to equate levels across pens within replicates (Table 1). Cattle were then transported 110 km to the research feedlot and penned as per previous randomization. Each pen contained at least 1 animal from each feedlot, with an average of 70% of AS1 cattle from Feedlot A and 80% of AS2 cattle from Feedlot B.
Number of animals per experimental group.
DFM, direct-fed antimicrobials; CFU, colony-forming units; LS, low-shedder detected by immunomagnetic separation.
Study experimental groups were as follows: DFM, finishing diets containing Bovamine® Culture Complex (Nutrition Physiology Company, LLC) with 109 CFU L. acidophilus and Propionibacterium freudenreichii fed/animal/d; VAC, standard finishing diets and 2 mL intramuscular injection of Econiche® (Bioniche Life Sciences Inc.) at allocation and 28 days later; or CON, standard finishing diets only. The vaccine is licensed in Canada for 3 doses (28 days apart), but the 2-dose regimen was approved by the manufacturer. Cattle were allocated on June 15 (AS1) and June 28 (AS2) 2011, respectively. Cattle (n=864) were housed by allocation set and experimental group in 48 pens with 18 cattle/pen and 1 pen from each experimental group constituting a replicate. Pens of AS1 cattle (n=30) were steers (initial weight 416.0±27.2 kg), while pens of AS2 cattle (n=18) were heifers (initial weight 402.1±28.3 kg). Experimental groups were in contiguous pens in separate feedlot alleys to avoid cross-contamination between interventions. Cattle were observed daily for animal health, managed using standard feedlot procedures, and followed for 103–145 days until shipment to slaughter.
Sample collection
At 28-day intervals, rectal fecal samples were collected from all cattle and perineal swabs were obtained from 4 random cattle/pen. All cattle were swabbed the day prior to slaughter (last handling event). To collect perineal swabs, a sterile SpongeSicle® (Med-Ox Diagnostics Inc., Ottawa, ON) scrubbed an area 100 cm2 below the anus and was transported in 45 mL modified E. coli broth (mEC). Samples were shipped chilled for processing within 24 h and stored at 5°C until completion of analyses.
E. coli O157:H7 enumeration, detection
Each fecal sample was mixed to promote uniformity, and two 1-g subsamples were enriched in 9 mL mEC (6 h at 37°C). Perineal swabs were incubated in mEC transport media (18 h at 37°C). Enriched samples were subjected to immunomagnetic separation (IMS) using anti-E. coli O157 Dynabeads® (Invitrogen, Carlsbad, CA). A 50-μL bead–bacteria mixture was plated on sorbitol MacConkey agar with 2.5 mg/L potassium tellurite and 0.05 mg/L cefixime (CT-SMAC) and incubated (18–24 h at 37°C). Three sorbitol-negative colonies/plate were evaluated using E. coli O157 latex tests (Oxoid, Nepean, Ontario, Canada), with positive colonies frozen in glycerol.
For enumeration, 1:10 dilutions from 1-g fecal subsamples positive by IMS were prepared in mEC with 20 mg/L novobiocin (EMD, Gibbstown, NJ), and 100-μL duplicates were plated on CT-SMAC. Plates contained 30–300 sorbitol-negative colonies, with 5 colonies latex tested for O157 antigen. Number of colonies/plate was adjusted as described by Stephens et al. (2009). From O157 colonies frozen in glycerol, polymerase chain reaction (PCR) confirmed E. coli O157:H7 (Gannon et al., 1997; Paton and Paton, 1998). Colonies were considered E. coli O157:H7 positive with eaeA, fliC, and at least one Shiga-toxin gene.
Pulsed-field gel electrophoresis (PFGE)
A random subset of isolates confirmed E. coli O157:H7 (n=42) balanced by allocation set; treatment and Shiga-toxin profile were subtyped by PFGE using XbaI restriction and standard 1-d protocol (Ribot et al., 2006). One E. coli O157:H7 isolate/positive sample was PFGE typed using a CHEF DR II electrophoresis unit (Bio-Rad Laboratories, Mississauga, ON). Banding patterns were viewed with UV illumination and photographed using Speedlight Platinum Gel Documentation System (Bio-Rad).
Statistical analyses
Analyses used SAS 9.2 (SAS Institute, Cary, NC) and were considered significant at p<0.05 with pen the experimental unit. Repeated-measure analyses for E. coli O157:H7 enumeration used mixed-models including experimental group, allocation set (AS), and days-on-trial as fixed effects, with three-way interaction of experimental group, AS and replicate as a random effect and days-on-trial as repeated variable. Prevalence of E. coli O157:H7 was analyzed using logistic methodology within GLIMMIX and as significant two- and three-way interactions with AS were present, analyses were performed by AS, with experimental group, sample type, and days-on-trial as fixed effects, and interaction of experimental group and replicate as random effects. Model-adjusted means (back-transformed to original scale) were reported, with efficacy estimated using the formula:
The PFGE patterns were classed as unique or grouped into restriction endonuclease digestion pattern clusters (REPC;≥90% similarity) using Dice similarity coefficients, unweighted-pair group methods arithmetic average algorithm, 1% position tolerance, and 0.5% optimization (BioNumerics 6.6, Applied Maths BVBA, Sin-Martens-Latem, Belgium). Within- and between-group PFGE profile similarities were tested using Dimensioning Techniques. For these analyses, band-matched binary character profiles were created after clustering using Dice coefficients. Bootstrap analysis (n=1000) then assessed the similarity of PFGE profiles.
Results and Discussion
Study timing and location
Based on previous Alberta studies (Stanford et al., 2005a; Stephens et al., 2009), peak E. coli O157:H7 prevalence usually occurs during August and September. Consequently, initial screening began in May, with cattle allocation by June. At allocation, E. coli O157 shedding was balanced across replicates and experimental groups (Table 1), with an average of 10% fecal positives in AS1 and AS2 (Fig. 1). Monitoring occurred during E. coli O157:H7 season, with cattle slaughtered September through early November. Although prevalence was highest in July (p<0.001; Table 2), E. coli O157:H7 was sufficiently prevalent for intervention comparison throughout the feeding cycle (Fig. 1).
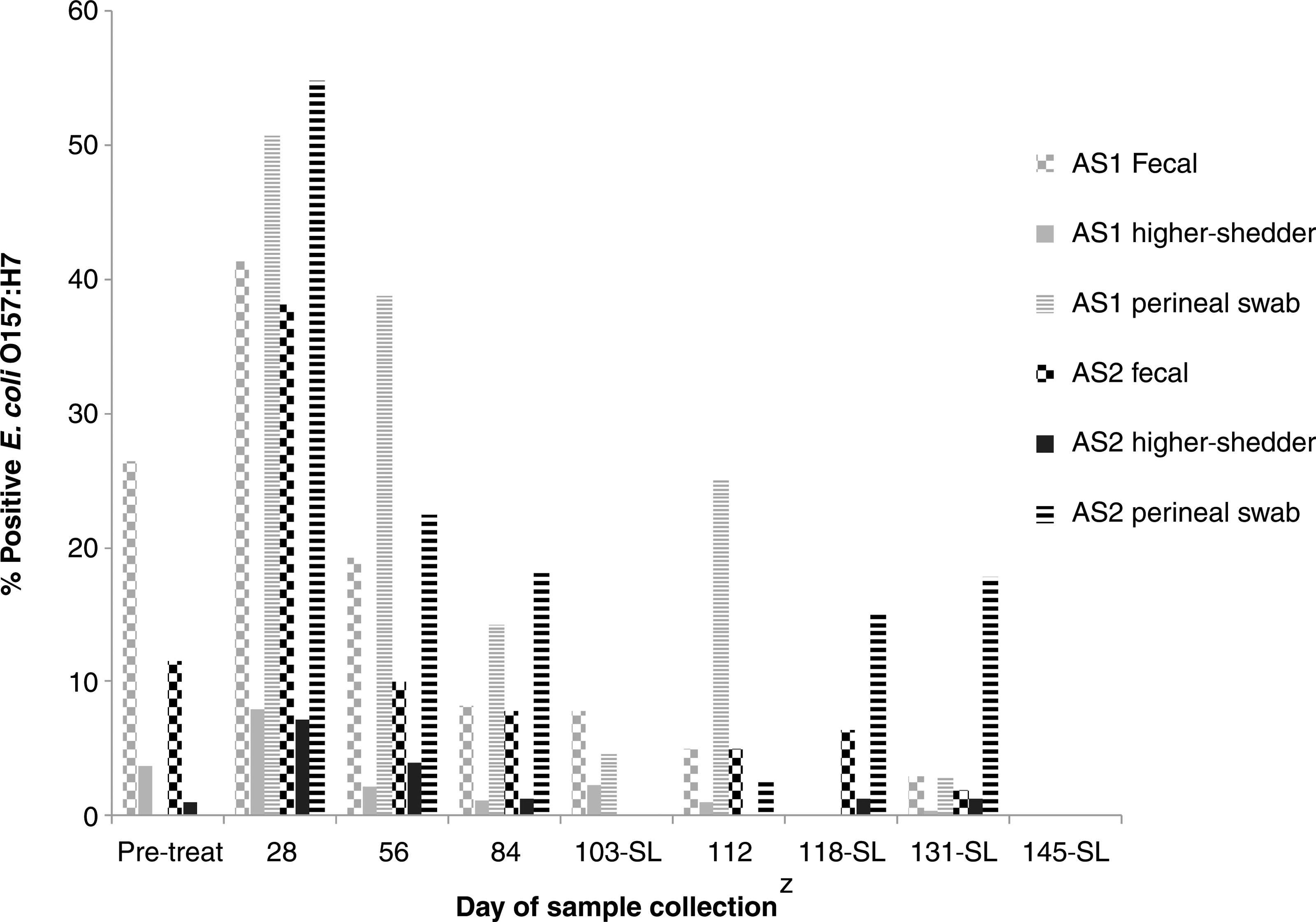
Fecal samples and perineal swabs positive for Escherichia coli O157:H7 and fecal samples with sufficient E. coli O157:H7 for enumeration (higher-shedder, ≥1.6 log colony-forming units/g feces) from steers (n=540) allocated on June 15, 2011 (AS1) and heifers (n=324), allocated on June 28, 2011 (AS2).
Allocation set 1, steers (n=540) allocated on June 15, 2011 into 30 pens.
Allocation set 2, heifers (n=324) allocated June 28, 2011 into 18 pens.
Includes both perineal swab and fecal grab positives.
CI, confidence interval.
Cattle were housed in small rather than in commercial pens (200–400 animals/pen). Animal behavior and disease transmission are known to differ in large- and small-pen comparisons (Jim et al., 1993; Perrett et al., 2008); however, identifying sufficient E. coli O157:H7-positive cattle is a challenge in large-pen studies. The small-pen design used ensured adequate and initially balanced prevalence of E. coli O157:H7 for experimental group comparisons and prevented cross-contamination of interventions. The DFM dosage was chosen to control E. coli O157:H7 as per past studies (Younts-Dahl et al., 2004; Peterson et al., 2007a; Vasconcelos et al., 2008) and has not previously improved feedlot performance (Elam et al., 2003; Peterson et al., 2007a; Vasconcelos et al., 2008).
Previous work evaluating pre-harvest interventions against E. coli O157:H7 ensured sufficient prevalence by inoculating cattle with E. coli O157:H7 (Allen et al., 2011) or using naturally colonized cattle sampled only during peak season (Cull et al., 2012). In the present study, a single fecal sample was used to balance E. coli O157:H7 levels across experimental groups and ensure sufficient prevalence as each pen had at least one animal shedding E. coli O157:H7 prior to initiation of interventions. Use of perineal swabs also enhanced detection as E. coli O157:H7 was 1.9 times more likely (p<0.001) to be detected from perineal swabs than from feces (Table 2), even though swabs were collected from 4 of 18 cattle during the feeding period and feces were collected from all cattle per pen.
E. coli O157:H7 enumeration
“Super-shedders” (104 or greater CFU E. coli O157:H7/g feces; Chase-Topping et al., 2008) have increased contamination of hides of pen mates (Stephens et al., 2009) and beef carcasses (Jacob et al., 2010). In contrast to super-shedders, most cattle positive for E. coli O157:H7 shed at relatively low levels (<100 CFU/g of feces) (Ferens and Hovde, 2011). As less than 10% of cattle may be super-shedders (Chase-Topping et al., 2008), cattle in this study were divided between “lower-shedders” (detectable by IMS) and “higher-shedders” (sufficient E. coli O157:H7 to enumerate; ≥1.6 log CFU/g feces).
The number of higher-shedder cattle was reduced 71.4%, by VAC (p=0.008; Fig. 2), which likely reduced the load of E. coli O157:H7 in VAC pens, although VAC impact on shedding duration was not assessed. While overall incidence of higher-shedders was reduced, VAC did not prevent cattle from becoming higher-shedders; VAC cattle shed up to 107 CFU E. coli O157:H7/g feces 1 month after booster and before final sampling (data not shown). Similar reductions in higher-shedder cattle have been reported for siderophore receptor-based vaccines (Fox et al., 2009; Cull et al., 2012).
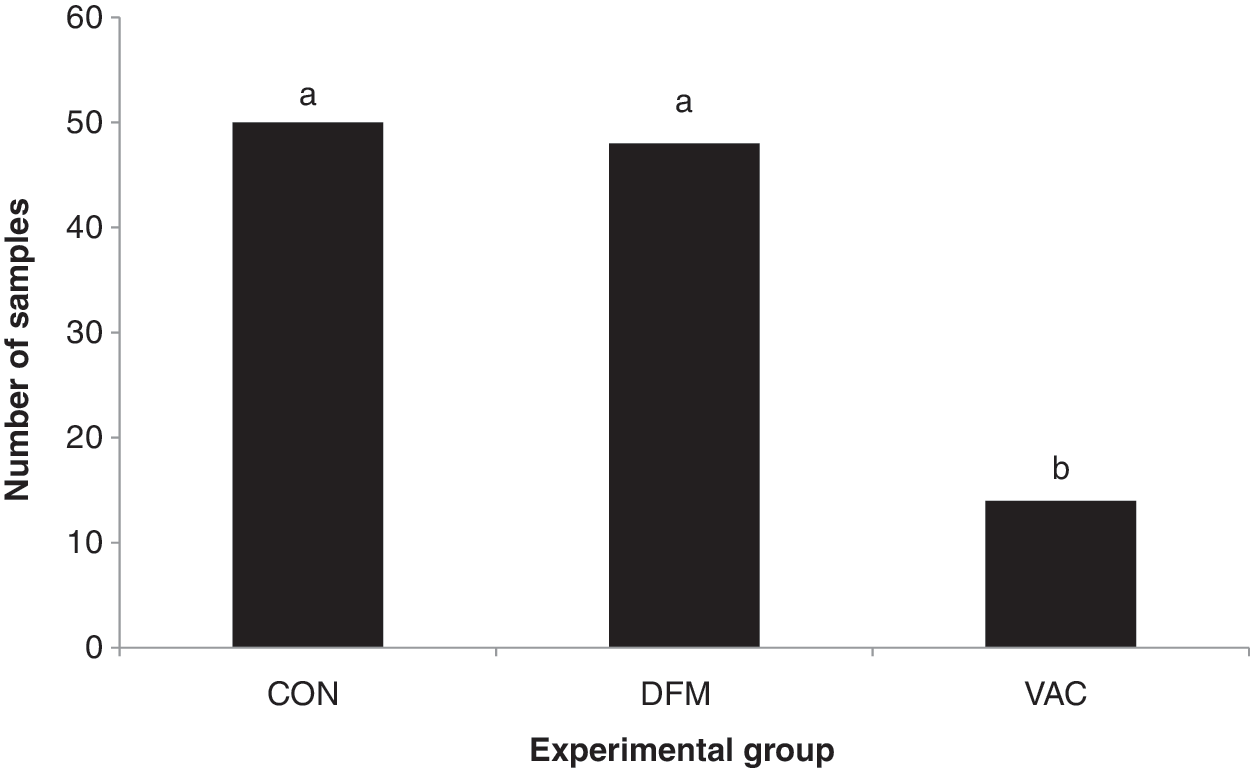
Incidence of fecal samples with sufficient Escherichia coli O157:H7 for enumeration (≥1.6 log colony-forming units (CFU)/g) from day 28 until shipment to slaughter for cattlez fed the direct-fed microbial Bovamine® (Nutrition Physiology Company, LLC) at a dosage of 109 CFU/animal/d Lactobacillus acidophilus and Propionibacterium freudenreichii starting on day 0 (DFM), given a 2-mL intramuscular injection of Econiche® (Bioniche Life Sciences Inc., Belleville, Ontario, Canada) on day 0, and a booster on day 28 (VAC), or given standard feedlot diets without DFM and not vaccinated (CON).
For cattle shedding E. coli O157:H7, average number of E. coli O157:H7 in feces did not differ by intervention, although AS2 feces were approximately 1 log higher (p=0.004) than AS1 (Fig. 3). Previous response to VAC for numbers of E. coli O157:H7 shed has varied. Potter et al. (2004) reported significant reductions in cattle inoculated with the organism, while Moxley et al. (2009) suggested that naturally colonized cattle shed relatively similar numbers regardless of VAC. For DFM, previous responses have also been mixed, with fecal E. coli O157:H7 numbers reduced at dosages of 109 CFU/d (Stephens et al., 2007a), while a companion study found no impact on numbers in cattle feces with DFM doses from 107 to 109 CFU/d (Stephens et al., 2007b).
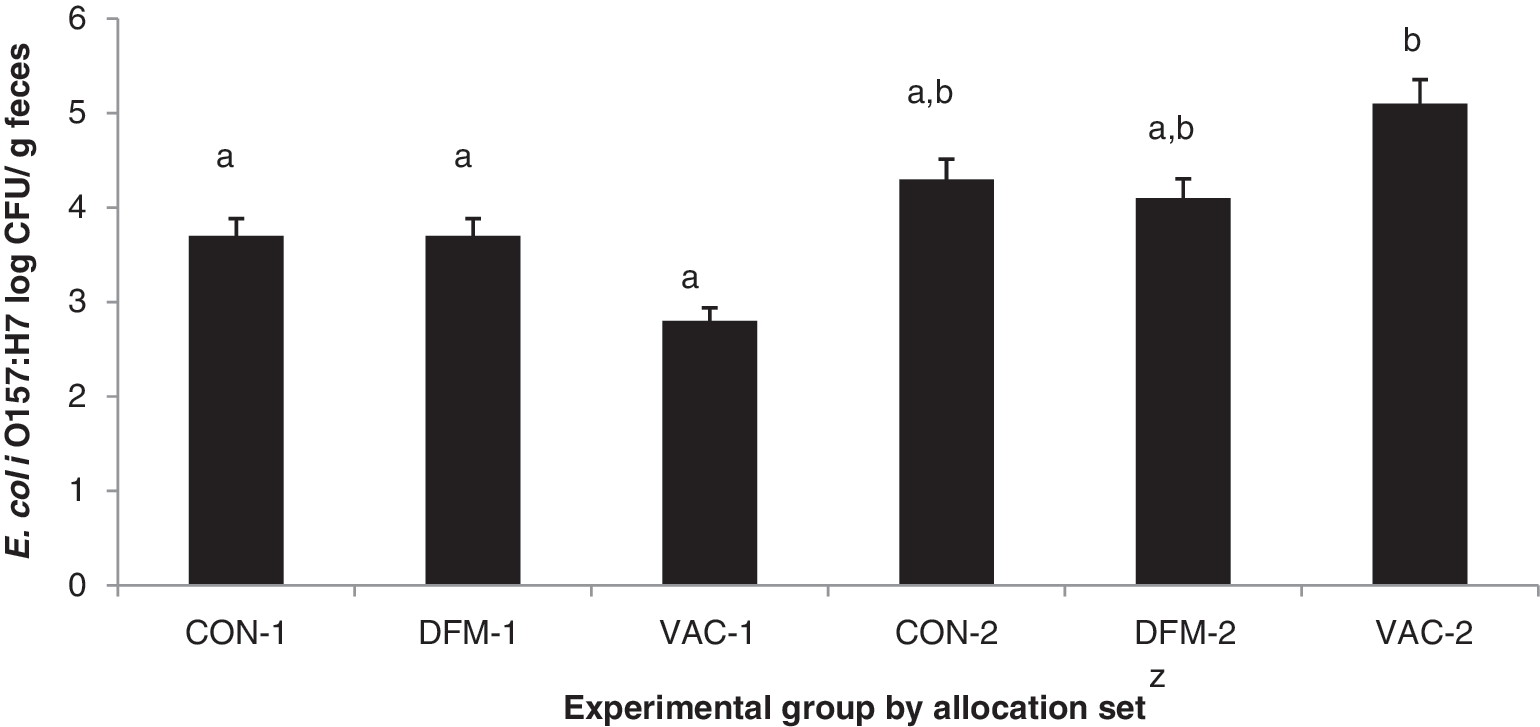
Average number of Escherichia coli O157:H7 per gram of feces by experimental groupy and allocation set (1 or 2)z for complete study period.
E. coli O157:H7 detection
E. coli O157:H7 prevalence in samples and intervention responses differed (p=0.02) by AS (Figs. 4 and 5). For AS2, neither intervention reduced E. coli O157:H7 prevalence. Compared to CON, AS2 VAC and DFM cattle had increased prevalence of E. coli O157:H7 in perineal swabs within the feeding period and at exit sampling (p<0.05). It is unlikely that interventions promoted E. coli O157:H7 shedding in AS2 cattle and would instead reflect high variability of shedding among individual AS2 cattle due to negligible impact of interventions. In contrast, VAC reduced E. coli O157:H7 prevalence in feces (p=0.03) and in perineal swabs (p=0.04) in AS1 cattle during the feeding period, although neither reduction was significant at exit sampling. Efficacy of VAC in AS1 during the feeding period for reducing E. coli O157:H7 prevalence was 60% for feces and 56% for perineal swabs (Fig. 4). For DFM, E. coli O157:H7 prevalence was reduced (p=0.01) in AS1 perineal swabs during the feeding period, although not at exit sampling (Fig. 5). However, fecal E. coli O157:H7 prevalence was never reduced in AS1 DFM-fed cattle. Having >10% of cattle positive for E. coli O157:H7 prior to intervention initiation (Fig. 1) likely negatively influenced DFM efficacy as proposed DFM mechanisms may be most effective prior to gastrointestinal tract colonization with E. coli O157:H7 (Callaway et al., 2004).
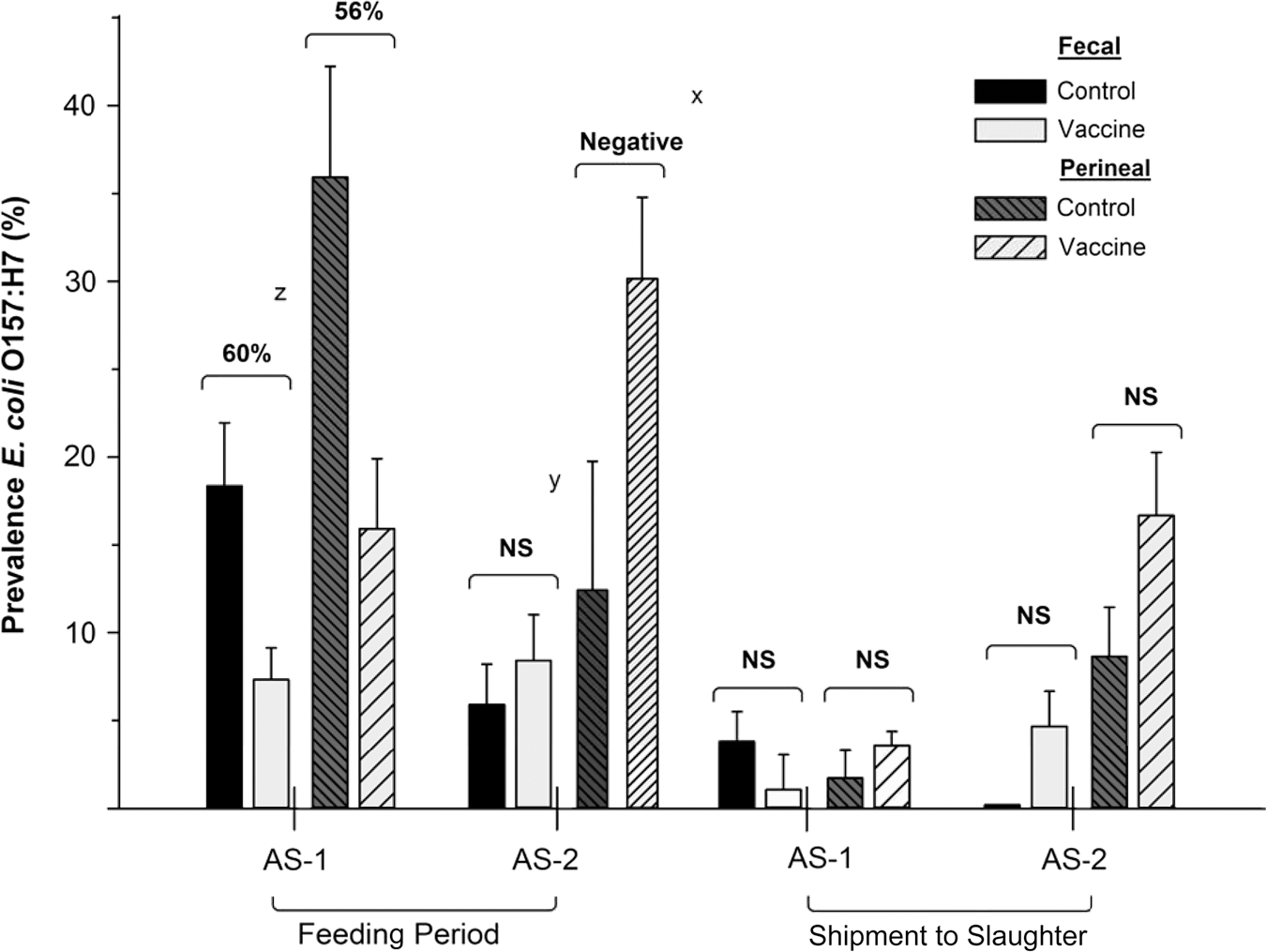
Overall impact of two doses of Econiche vaccine® (Bioniche Life Sciences Inc., Belleville, Ontario, Canada) on day 0 and a booster on day 28 on detection of Escherichia coli O157:H7 in feces and hide swabs from allocation set 1 (AS1; steers allocated June 15, 2011) and allocation set 2 (AS2; heifers allocated June 28, 2011) during the feeding period (cumulative incidence from day 56 to day 112) until shipment to slaughter (single-day incidence occurring day 103 to day 145).

Overall impact of the direct-fed microbial (DFM) Bovamine® (Nutrition Physiology Company, LLC) fed to allocation set 1 (AS1; steers allocated June 15, 2011) and allocation set 2 (AS2; heifers allocated June 28, 2011) at a dosage of 109 colony-forming units (CFU) Lactobacillus acidophilus and Propionibacterium freudenreichii per animal per day during the feeding period from day 0 until shipment to slaughter on detection of Escherichia coli O157:H7 in feces and hide swabs during the feeding period (cumulative incidence from day 28 to day 112) until shipment to slaughter (single-day incidence occurring day 103 to day 145).
Although AS1 and AS2 were steers and heifers, respectively, gender would not likely impact intervention efficacy. A 2-week sampling schedule difference may have contributed to differing efficacies, but if minor changes in sample collection timing were responsible for these differences, DFM and VAC would be of limited utility for controlling E. coli O157:H7 in commercial livestock. Possibly, differences in E. coli O157:H7 populations for AS1 and AS2 influenced intervention impacts. From PCR analyses, toxin gene frequencies differed between AS (p=0.004; data not shown), with 87% of isolates from AS2 having stx1 and stx2, while 75% of isolates from AS1 cattle had only stx1. Although stx2 has suppressed development of cellular immune responses to E. coli O157:H7 in cattle (Hoffman et al., 2006), evaluation of a subset of AS2 data excluding isolates with stx2 did not improve efficacy (data not shown). Matthews et al. (2013) proposed that cattle shedding E. coli O157:H7 with stx2 were more likely to be super-shedders and that presence of stx2 was a primary risk factor for human disease. Consequently, reduced efficacy of VAC in AS2 as compared to AS1 cattle may limit impacts of VAC on human health. Additional study is required regarding estimated impacts of interventions on human health and the role of stx2 on intervention efficacy.
Variable efficacy of VAC and DFM in different populations of cattle would be consistent with other studies. A review of vaccines to reduce ruminant fecal shedding E. coli O157:H7 (Snedeker et al., 2012) concluded that care should be taken interpreting findings due to variability among studies. When pre-harvest and at-harvest outcomes were considered for either type-III secreted proteins or siderophore receptor and porin protein vaccines, meta-analysis also showed heterogeneous results (Varela et al., 2012), although these authors found homogeneity in pre-harvest outcomes in two-dose studies of VAC. As E. coli O157:H7 is a commensal organism not causing disease in cattle, variable immune responses by animals may contribute to heterogeneous vaccine efficacy (Snedeker et al., 2012). In the present study, VAC response varied by AS for pre-harvest measures and did not impact at-harvest measures of E. coli O157:H7 contamination.
A large-scale feedlot evaluation of DFM efficacy (Cull et al., 2012) also reported DFM ineffective for reducing fecal-shedding of E. coli O157:H7, although these authors did not measure hide contamination and used 106 CFU L. acidophilus/head/d. Feeding the 109 CFU dosage, DFM previously reduced E. coli O157:H7 detection in feces and hides (Brashears et al., 2003) or in feces but not hides (Younts-Dahl et al., 2005; Stephens et al., 2007b). In this study, it is intriguing that both DFM and VAC interventions were most effective in AS1 and did not control E. coli O157:H7 prevalence in AS2. More universally effective pre-harvest interventions are required, and the vaccine manufacturer in this study has recently announced initiation of research for a second-generation E. coli O157 vaccine (Anonymous, 2013).
PFGE genetic variability
From PFGE analyses, seven REPC and nine unique isolates were detected, with most isolates from AS1 and AS2 in separate REPC (data not shown). In REPC with co-mingling of AS1 and AS2, only atypical AS2 isolates lacking stx2 were present. Generally, isolates in REPC were from a single AS and had similar stx genes; the only exception was REPC 6, where AS1 cattle with stx1 or both stx1 and stx2 were present. As AS1 and AS2 cohorts were mostly from different feedlots and would have been penned separately at those feedlots as well as in the present study, PFGE subtype bifurcation by AS is not surprising. E. coli O157:H7 subtypes can vary by farm (Rice et al., 1999; Stanford et al., 2005b) and by pen within commercial feedlots (Sargeant et al., 2006; Stanford et al., 2012), possibly related to the farm of origin of the cattle.
From bootstrapping analyses, AS1 isolates had more similar PFGE profiles compared to all isolates (p<0.001), while AS2 isolates showed less uniformity (p=0.08; Table 3). The PFGE profiling revealed that within-group similarity of isolates with stx1 alone was higher than when compared to all isolates (p<0.001), while within-group similarity of isolates with stx1 and stx2 genes was lower when compared to all isolates (p=0.11). Subdividing isolates with stx1 and stx2 genes by AS, both sets of PFGE profiles were diverse. Overall, PFGE analyses demonstrated that AS1 E. coli O157:H7 subtypes were relatively homogeneous compared to increased diversity of AS2 isolates. This may at least partially explain the limited efficacy of interventions for AS2. Subtypes of E. coli O157:H7 from AS1 and AS2 may differently express genes in the locus of enterocyte effacement (LEE) encoding the type III proteins secretion system (Roe et al., 2004). Alternatively, proteins other than those encoded by LEE also contribute to E. coli O157:H7 adherence in the bovine gastrointestinal tract (Kudva et al., 2012) and may be more common in AS2. Studies to further characterize E. coli O157:H7 isolates from AS1 and AS2 are in progress.
NA, not applicable as minimum of two needed for comparisons.
Conclusions
Although VAC reduced the numbers of cattle shedding enumerable E. coli O157:H7, it did not prevent individuals shedding up to 107 CFU/g feces. Neither VAC nor DFM consistently reduced prevalence of E. coli O157:H7 in feces or perineal swabs from either AS of cattle, and neither intervention lowered numbers of E. coli O157:H7 shed in feces. Intervention efficacy may have been negatively influenced by an initial shedding rate (>10% of cattle) and presence of different E. coli O157:H7 subtypes. As VAC and DFM had variable efficacy, additional development is required prior to recommending either pre-harvest E. coli O157:H7 intervention to the commercial beef industry.
Footnotes
Acknowledgments
This work was funded by the Alberta Livestock and Meat Agency.
Disclosure Statement
Bioniche Life Sciences Inc. and Nutrition Physiology Company LLC. contributed product and funding to Feedlot Health Management Services Ltd. to collectively offset 9% of project costs.