
Abstract
Staphylococcus aureus is one of the major bacterial species that may cause clinical infection and food-poisoning cases. Strains of this bacterial species may produce a series of superantigens (SAgs) (i.e., staphylococcal enterotoxins [SEs], staphylococcal enterotoxin-like toxins, and toxic shock syndrome toxin). In this study, S. aureus strains from clinical samples and food-poisoning cases in Taiwan were collected; their SAg profiles, and SmaI digestion patterns determined by pulsed-field gel electrophoresis (PFGE), were then analyzed. Results showed that their SAg gene profiles and SmaI digestion patterns of chromosomal DNA were highly diverse. Although PFGE has been used as a criterion standard for typing of S. aureus strains, and the SAg profiles have been used in combination with PFGE for typing of S. aureus strains, we found that strains grouped in these combined patterns could be further discriminated by the random amplified polymorphic DNA (RAPD) method. Thus, the combined use of SAg profiles, PFGE, and RAPD patterns permits high discrimination for typing of S. aureus strains from not only the clinical samples but also the food-poisoning cases. Such a combined method may be used as a highly accurate approach for epidemiological study and tracing of the contamination origin of staphylococcal infections either in hospitals or food-poisoning cases.
Introduction
S
Pulsed-field gel electrophoresis (PFGE) is regarded as an effective tool to distinguish S. aureus strains. SmaI restriction digestion of DNA has been reported to yield the best results in PFGE analysis and been routinely used for epidemiological study (Saulnier et al., 1993; Bens et al., 2006). Other methods for typing of S. aureus strains include random amplification of polymorphic DNA (RAPD) and toxin gene profiles (Pereira et al., 2002; Xie et al., 2011; Mehndiratta and Bhalla, 2012). RAPD involves random amplification of segments of target DNA using short primers, which is simple, fast, inexpensive, and could be used for epidemiological study of S. aureus (Morandi et al., 2010; Mehndiratta and Bhalla, 2012). Toxin gene profiles of S. aureus could be used for the elucidation of the genetic and pathogenic relatedness between strains. They are easy to interpret and have been used as epidemiological markers for typing of S. aureus (Xie et al., 2011; Mehndiratta and Bhalla, 2012).
Although horizontal transfer of mobile genetic elements containing virulence genes between S. aureus strains may lead to the emergence of many different toxin gene combinations in S. aureus cells during evolution (Diep et al., 2006; Xie et al., 2011), reports have shown that transfer of these virulence genes is infrequent and the virulence gene expression pattern in S. aureus is stable (Diep et al., 2006; Ziebandt et al., 2010). Several studies have characterized SAg genes in methicillin-resistant S. aureus (MRSA) for epidemiological investigation (Coia et al., 1992; Schmitz et al., 1997; Diep et al., 2006). Distribution of staphylococcal toxin genes in food-poisoning isolates collected in Taiwan has been reported also (Chiang et al., 2008). Xie et al. (2011) suggested that toxin gene typing, combined with PFGE or multilocus sequence typing (MLST), could increase the discriminatory power of genotyping S. aureus strains. Based on the above-described facts, in this study, S. aureus strains from clinical samples and food-poisoning cases were analyzed by PFGE, SAg genes (i.e., sea–see, seg–seu, and tst), and RAPD typing. These three typing methods were then combined for the discrimination of S. aureus isolates. Results showed that the combination of these three typing methods would improve the strain discrimination substantially.
Materials and Methods
Bacterial strains and cell cultivation
A total of 211 S. aureus isolates from food-poisoning cases and clinical samples were collected for this study. Of these isolates, 73 isolates (HCT 1–39, 41–47, 49–61, and 63–76) were from clinical blood samples of hospitalized patients with bacteremia. Another 138 isolates (HYT 1–50, 53–70, 72–86, 88–96, 98–112, 114–117, 119–126, 128–132, and 134–147) were from the vomit and fecal specimens of persons associated with food-poisoning cases. All these strains were identified according to the methods described in Bacteriological Analytical Manual (Food and Drug Administration, 1998). Reference enterotoxigenic strains from Bioresource Collection and Research Center, Hsin-Chu, Taiwan, and strains previously characterized as superantigenic S. aureus strains were used as positive controls (Chiang et al., 2006; Chiang et al., 2008). Bacterial cells were cultivated in tryptic soy broth (30 g in 1000 mL) or brain heart infusion (37 g in 1000 mL) followed by 8-h incubation at 37°C for further plating or DNA preparation. All media used in this study were purchased from Difco™ (Becton, Dickinson and Company, Sparks, MD).
Detection of the genotypes of SAgs
Bacterial genomic DNA of S. aureus was prepared as previously described (Chiang et al., 2008). The SAg profiles of S. aureus isolates were then determined with polymerase chain reaction (PCR) primers specific for classical enterotoxins (SEA–E), new types of SAgs (SEG–SEI, SElJ–SElQ, SER, and SElU), and TSST-1 as previously described (Chiang et al., 2006, 2008).
PFGE
The isolation of chromosomal DNA of S. aureus and SmaI digestion were basically according to the methods described by Tsen et al. (1997). Electrophoresis was performed by using the CHEF-DR®III Variable Angle System (Bio-Rad, Hercules, CA). The conditions used were 6 V/cm for 10 h at 14°C (pulse time: 5–15 s) and an additional 15 h at 14°C (pulse time: 15–45 s). Bacteriophage λ DNA concatemers (Bio-Rad) were used as molecular weight markers. PFGE patterns were analyzed by the NTSYSpc software (numerical taxonomy and multivariate analysis system, version 2.10e, State University of New York, Stony Brook, NY). Strains were clustered by using the Dice coefficient of similarity, and cluster analysis by unweighted pair group method with arithmetic averages (UPGMA). The cutoff lines for clustering the PFGE types were set at 52% and 80% similarity, respectively.
Randomly amplified polymorphic DNA (RAPD)
RAPD was performed with primers OPL-05 (5′-ACGCAGGCAC-3′), AP4 (5′-GAGGGTGGCGGTTCT-3′), and M13 (5′-TCACGCTGCA-3′), respectively. The amplification conditions, the gel electrophoresis, and the analysis of amplified products were basically according to the methods of Sánchez et al. (2004) and Perillo et al. (2012). RAPD patterns were analyzed by the GelCompar software (version 2.5, Applied-Maths, Kortrijk, Belgium). Strains were clustered by using the Dice coefficient of similarity and cluster analysis by UPGMA. The cluster cutoff line for RAPD patterns was set at 90% similarity.
Results
PCR determination of the SAg profiles for S. aureus strains from clinical samples
Sixty-six of the 73 clinical isolates (90.4%) harbored at least one SE or TSST-1 gene; 7 isolates were nonenterotoxigenic or nonsuperantigenic. Most of the superantigenic strains of S. aureus harbored both classical and new SAg genes. Of these strains from clinical samples, there were a total of 36 different combinations of SAg genes; the main SAg profile was sea-seq (9 in 73 strains), followed by sep (8 in 73 strains). Details of the SAg profiles are listed in Table 1.
Total: 36 superantigen profiles.
Number of strains with the same superantigen gene profile.
ND, superantigen genes assayed were not detected.
PFGE patterns and SAg profiles
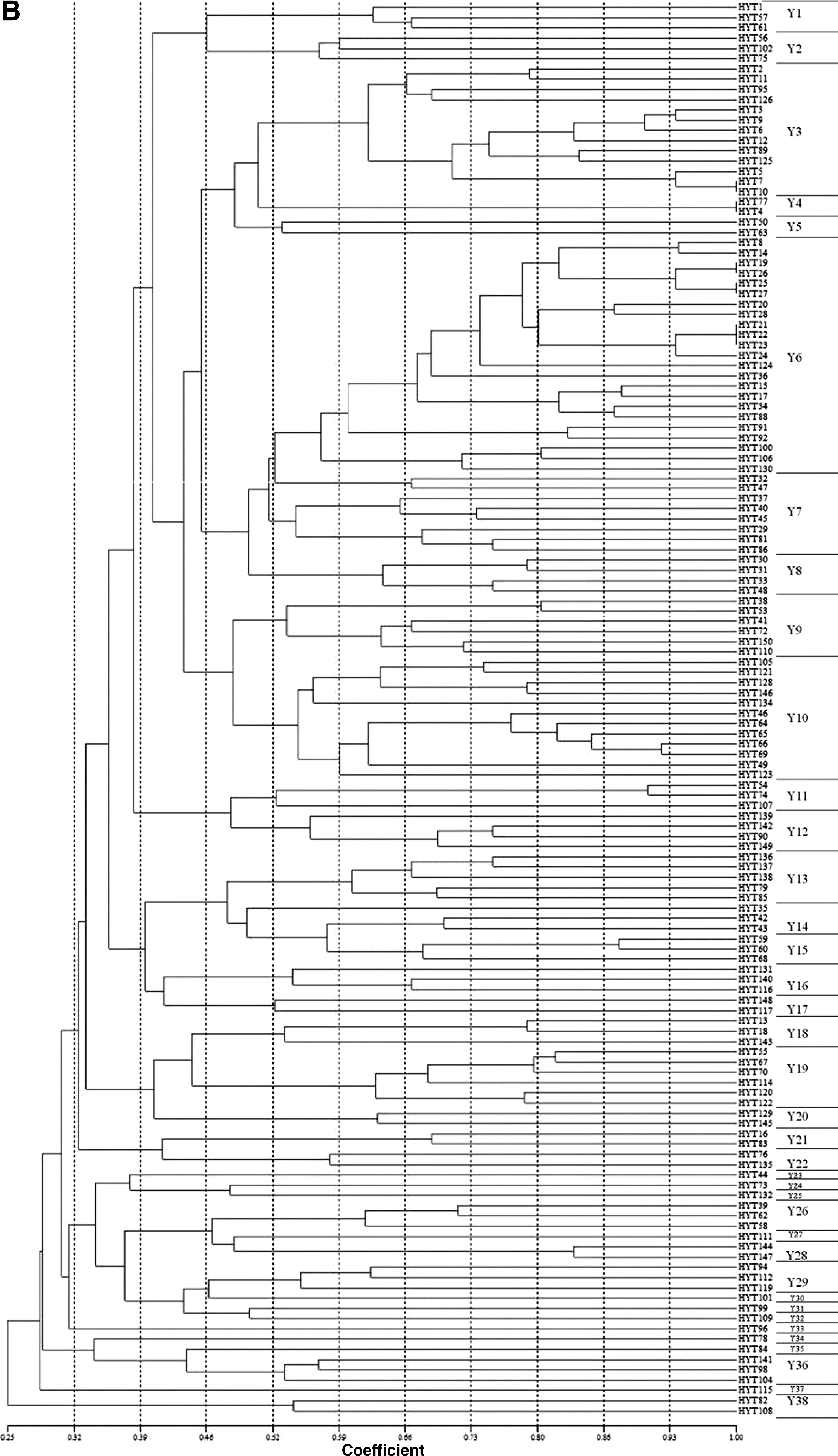
The SAg profiles and PFGE patterns were used for the discrimination of S. aureus strains from clinical samples and food-poisoning cases. When chromosomal DNA of the 73 clinical and 138 food-poisoning isolates were digested with SmaI, highly genetic diverse patterns for strains from both origins were found. Thus, the cutoff was set at 52% similarity for clustering. With the 52% similarity, 20 main clusters (i.e., C1–C20) were found for clinical isolates (Fig. 1A, Table 2). On the other hand, for food-poisoning isolates, 38 PFGE clusters (i.e., Y1–Y38) were observed (Fig. 1B, Table 3). Most of these individual clusters consisted of more than one isolate. When the cutoff was set at 80% similarity, the number of clusters obtained was greatly increased.
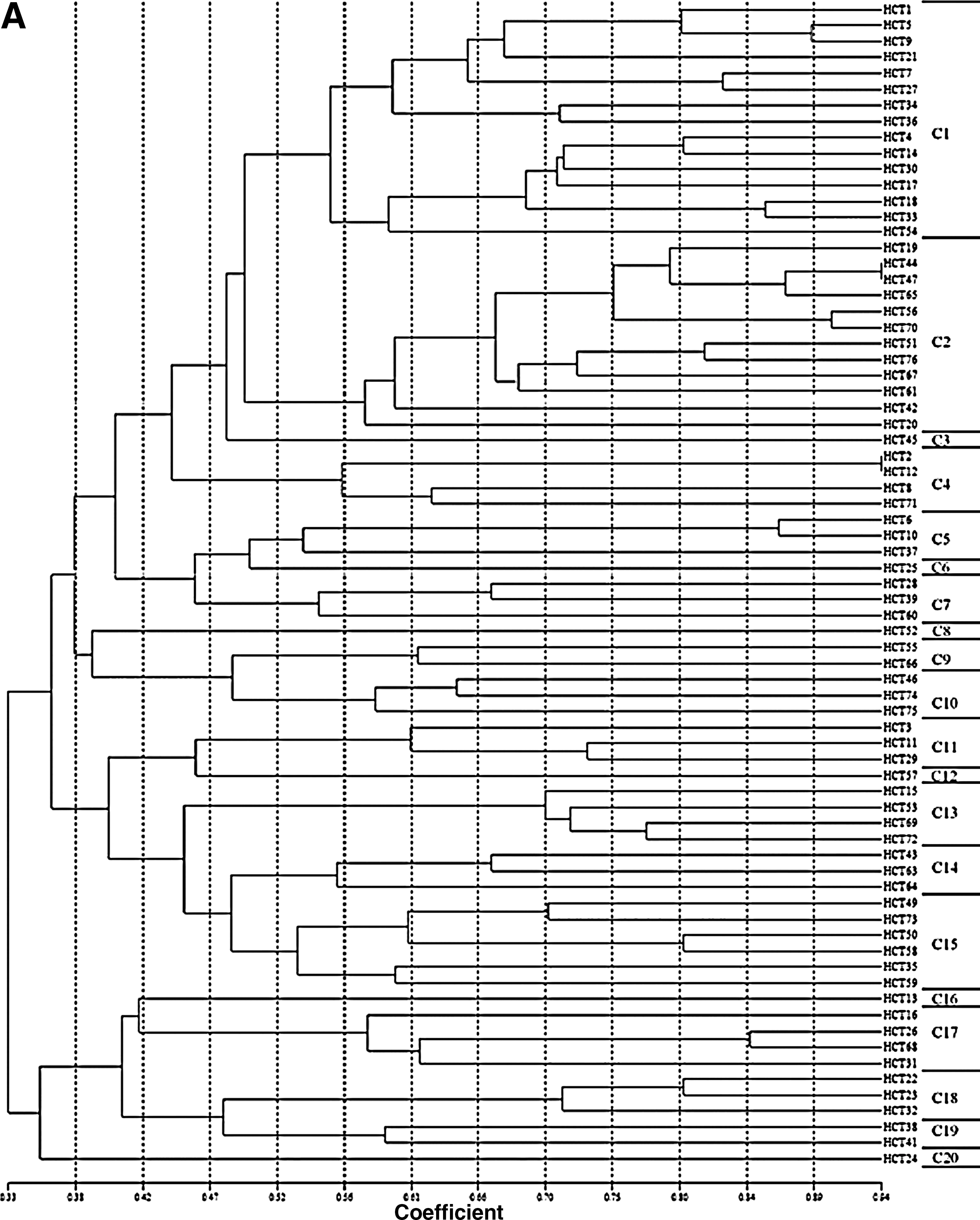
Dendrogram showing the similarities of pulsed-field gel electrophoresis patterns for Staphylococcus aureus strains isolated from
Total: 20 PFGE clusters (with cutoff at 52% similarity), 59 combined patterns.
Number of strains with the same superantigen gene profile.
ND, superantigen genes assayed were not detected.
Total: 38 PFGE clusters (with cut-off at 52% similarity), 114 combined patterns.
Number of strains with the same superantigen gene profile.
ND, superantigen genes assayed were not detected.
When SAg profiles and SmaI digested patterns with 52% similarity were combined to discriminate these strains, 59 and 114 combined patterns were obtained for the clinical and food-poisoning isolates, respectively (Tables 2 and 3). The SAg genes were highly diverse in strains of each of the PFGE clusters. For most of the individual PFGE clusters, more than one of the SAg profiles were found except for cluster Y28, in which no SAg genes were detected, and those with only one strain. Nevertheless, there were 62 strains that remained indistinguishable, for example, strains in clusters of combined patterns of C1-sea-seq and Y3-sea. However, if the cutoff was set at 80% similarity, only 15 strains remained indistinguishable (data not shown).
RAPD for further discrimination of S. aureus strains
Strains remaining indistinguishable by the combined use of SAg profiles and PFGE were further analyzed by RAPD (cutoff with 90% similarity); 24 clinical and 38 food-poisoning isolates were divided into 11 and 16 RAPD clusters, respectively (Fig. 2). When all the three methods (i.e., PFGE, SAg profiles, and RAPD) were combined for the discrimination of these 24 clinical and 38 food-poisoning isolates, 21 and 25 combined patterns were obtained, respectively (Table 4). Some strains within the same combined patterns of PFGE and SAg profiles could be further subdivided by the RAPD method. Taking account of RAPD, the discriminatory power could be further improved for typing of S. aureus strains from both origins.
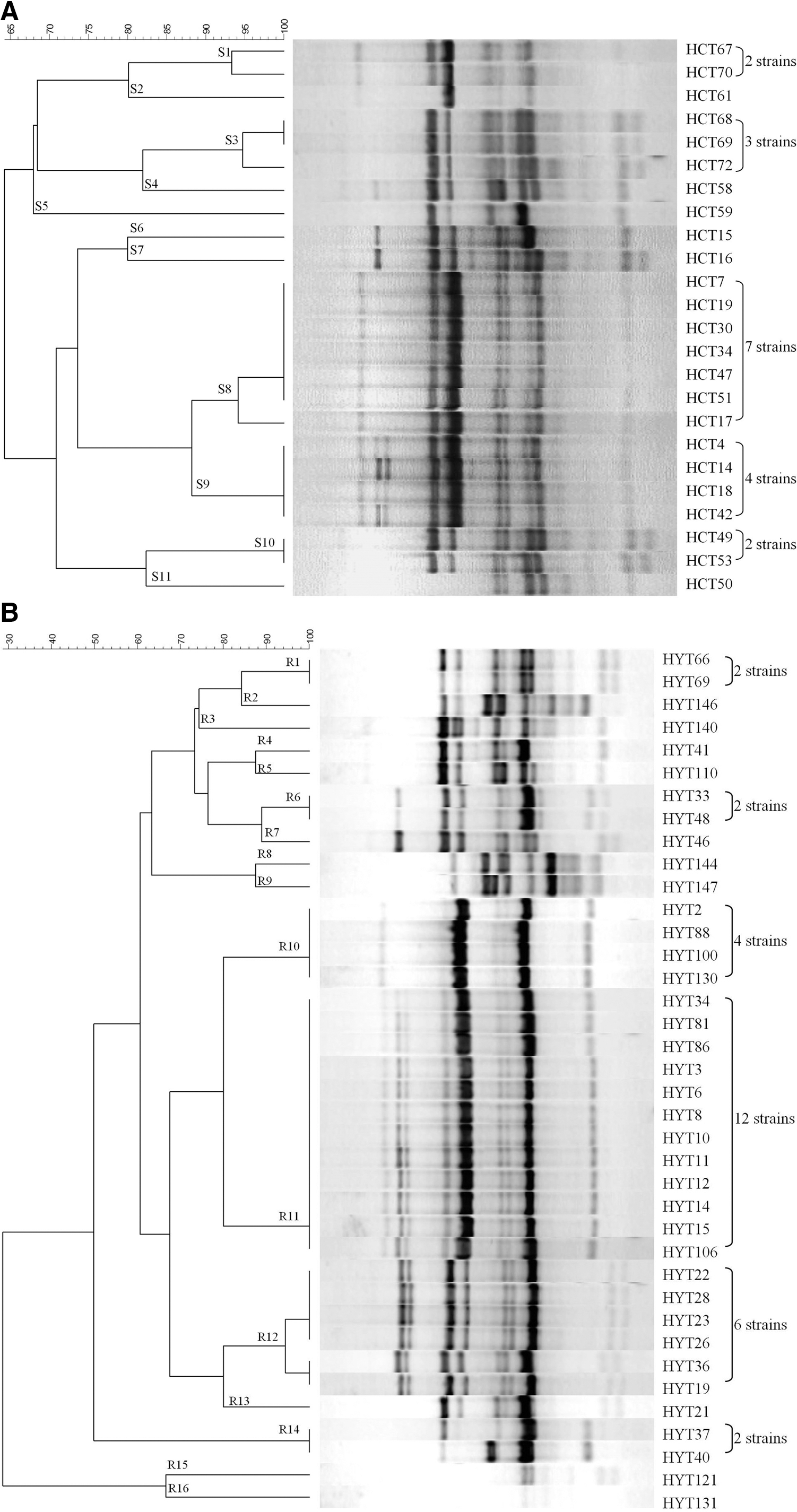
Dendrogram showing the similarities of random amplified polymorphic DNA patterns using primer OPL-05 for Staphylococcus aureus strains isolated from
Number of strains within the same superantigen gene profile.
ND, superantigen genes assayed were not detected.
Different cutoff values have been used for PFGE typing of S. aureus strains; however, the cutoff value with 80% pattern similarity was the one often used (Donate-Correa et al., 2011; Xie et al., 2011; Aklilu et al., 2012; Camoez et al., 2013). Thus, in addition to the 52% cutoff value, which generated the main PFGE clusters shown in Tables 2 and 3, the PFGE patterns we obtained for all strains were also analyzed with 80% similarity. Results showed that the discrimination power for S. aureus strains could be greatly improved. However, some strains still needed the RAPD methods for further discrimination (Table 5). As for the strains not discriminated by the combined typing methods, including PFGE, SAg profiles, and RAPD, especially when the cutoff line of the PFGE pattern was set at 80% similarity, these strains may be regarded as the same strain. These strains, when analyzed with RAPD methods using different primers (i.e., AP4, and M13) were still not discriminated (Table 5).
Number of strains within the same superantigen gene profile or RAPD clusters generated by primers OPL05, AP4, or M13.
Not determined.
ND, superantigen genes assayed were not detected.
Discussion
Distribution of staphylococcal toxin genes in S. aureus isolates from different origins has been reported. For clinical S. aureus isolates in China, 68.1% were positive for at least one virulence gene (Wang et al., 2010). For foodborne S. aureus isolates in Turkey; 62.6% were enterotoxigenic (Aydin et al., 2011). Previously, we assayed the distribution of SEA to SEE, SEG to SER, SEU, and TSST-1 genes in S. aureus isolates from food-poisoning cases in Taiwan; 91.8% were enterotoxigenic or superantigenic strains (Chiang et al., 2008). In this study, for all the 73 clinical isolates from hospitals, 67 (90.4%) are positive for at least one SE (SAg) or TSST-1 gene. Such a ratio indicates that the percentage of enterotoxigenic or superantigenic strains in either the clinical or the food-poisoning isolates collected in Taiwan was considerably high.
PFGE has been considered a criterion standard for typing of MRSA strains due to its highly discriminatory power and reproducibility (Mehndiratta and Bhalla, 2012). In this study, all the clinical and food-poisoning isolates of S. aureus were analyzed by the PFGE method, and diverse patterns were found for these strains. However, a number of strains remained indistinguishable by using PFGE alone, even when the cutoff line for PFGE clustering was adjusted from 52% to 80%. Thus, PFGE alone may not be sufficient for the discrimination of S. aureus strains. Since Xie et al. (2011) has reported that for genotyping S. aureus, PFGE appears to be more discriminatory than MLST, and toxin profiles combined with PFGE or MLST are able increase the discriminatory power of genotyping S. aureus strains, we thus combined the SAg profiles and PFGE for typing of S. aureus strains. In this regard, the stability of SE or SAg genes in S. aureus strains should be considered. In addition to the earlier-described fact that there is horizontal transfer of the SE or SAg genes between MRSA strains (Diep et al., 2006), Ziebandt et al. (2010) surveyed the composition and variability of the exoproteome of 25 clinical isolates of S. aureus from Netherlands and found that the virulence gene expression pattern may be very stable over extended periods of time. In addition, characteristic consensus virulence-gene profiles were observed; only relatively few variants deviate from the consensus. Furthermore, it is reported that combinations of SE genes were highly conserved (Varshney et al., 2009). Thus, the SAg profiles of S. aureus strains may be used to improve the discrimination of S. aureus strains.
In this study, for all the S. aureus strains, SAg profiles of these strains were highly diverse. Varshney et al. (2009) investigated the prevalence of 19 toxin genes in 207 S. aureus strains obtained from hospitals in New York State and found that S. aureus strains exhibited distinct combinations of SE genes, even if their MLST backgrounds or PFGE patterns demonstrated clonality. Our findings agreed with the results described by Varshney et al. (2009). Xie et al. (2011) found that diversified toxin profiles were present in the clinical isolates of S. aureus collected in China. The genomic diversity of these clinical isolates was also evaluated. The authors concluded that for PFGE, even with the cutoff line set at 80% similarity, the toxin profile could further increase the discriminatory power for typing of S. aureus strains. When we discriminated our S. aureus strains by PFGE, even with a cutoff line set at 80% similarity, the SAg profiles were needed to increase the strains discrimination.
Grundmann et al. (2002) reported that DNA-based typing methods come to very similar conclusions. There was a high degree of agreement between PFGE and MLST, and a fair degree of agreement between MLST and RAPD for typing S. aureus strains. It is also reported that sequence-based typing methods (for example, MLST and spa typing) showed good correlation with PFGE (Frickmann et al., 2012). The combining of different techniques for typing of S. aureus may not increase the discriminatory power to a large extent (Grundmann et al., 2002). However, different conclusions were made by Morandi et al. (2010). In their study, the distribution of toxin genes (sea, sec, sed, seg, seh, sei, sej, and sel), biochemical profiles, and other molecular techniques including RAPD patterns of S. aureus strains were investigated. They found that an effective strain differentiation can be achieved by the combination of different typing techniques and toxin gene patterns.
In this study, we found that although the use of PFGE in combination with SAg profiles would improve the strain discrimination for S. aureus, there were 24 clinical and 38 food-poisoning isolates remaining indistinguishable. As the cutoff line was adjusted from 52% to 80% similarity, the total number of the indistinguishable strains could be reduced from 62 strains to 15 strains. Since it is reported that the overall correlation between PFGE and RAPD is lower than that between PFGE and MLST (Grundmann et al., 2002), thus, although RAPD is less discriminatory than PFGE for typing of S. aureus (Saulnier et al., 1993), the low correlation between PFGE and RAPD may allow us to combine RAPD to improve the discriminatory ability.
In addition, the presence of toxin genes is not associated with particular RAPD patterns (Morandi et al., 2009, 2010), and the correlation of groupings obtained between the classical enterotoxin typing and the RAPD techniques is low (Pinto et al., 2005). Martin et al. (2004) suggested that RAPD in combination with toxin profiles could be used as an alternative typing method to PFGE. Thus, in our study, RAPD was applied to those strains remaining indistinguishable by PFGE and SAg profiles. When all three methods (i.e., PFGE, SAg profiles, and RAPD) in combination were used, some strains from not only the clinical samples but also the food-poisoning cases could be further differentiated into distinct subtypes. Such methods may allow us to understand the persistence or transmission of S. aureus strains in a precise way for epidemiological study.
In conclusion, we demonstrated that the SAg profiles are highly diverse for S. aureus strains. The sole use of PFGE, even with a pattern similarity at 80%, may not be sufficient for the discrimination of S. aureus strains. The combination of different typing methods could increase the discriminatory ability for the differentiation of S. aureus strains, and the combined use of SAg profiles, PFGE, and RAPD is highly discriminatory for typing of S. aureus strains. Such a typing approach may also allow us to differentiate S. aureus strains from other origins.
Footnotes
Acknowledgments
We would like to thank the National Science Council, Taipei, Taiwan for supporting this project (NSC 94-2313-B-241-001 and NSC101-2313-B-241-004-MY2).
Disclosure Statement
No competing financial interests exist.