
Abstract
Avian pathogenic Escherichia coli (APEC) strains harbor a number of virulence genes and cause extraintestinal diseases, such as septicemia, swollen-head syndrome, salpingitis, and omphalitis in poultry. APEC strains are not known to cause intestinal diseases. Herein, for the first time, it is reported that APEC strains were able to induce an enterotoxigenic-like effect in rabbit ligated ileal loops. Strain SEPT362 caused cell detachment of the intestinal villi, which also showed a flattened and wilted appearance, but the integrity of the tight junctions was maintained. Additionally, this strain did not adhere to enterocytes in vivo, although adhesin encoding genes ( fimH, csgA, lpfA2-3, and ECP) were present while other lpfA types, sfa, afa, papC, and ral genes were not. This enterotoxigenic-like activity was conserved after thermal treatment of the supernatant at 65°C but not at 100°C. Moreover, experiments based on filtering with different molecular weight cut-off (MWCO) pore sizes demonstrated that the component associated with the observed biological effect has a molecular weight >100 kDa. Blast search and polymerase chain reaction assays for known E. coli virulence factors showed that strain SEPT362 harbors the gene encoding for the toxin EAST-1 and the serine protease autotransporter (SPATE) Tsh, but is negative for genes encoding for the toxins LT-I, STh, STp, Stx1, Stx2, CNF-1, CNF-2, CDT and the SPATEs Sat, Pic, Vat, SigA, SepA, EatA, EspP, or EspC. A cloned copy of the tsh gene in E. coli K-12 was also tested and was shown to have an enterotoxic effect. These results suggest that APEC might induce fluid accumulation in the rabbit gut. The Tsh autotransporter seems to be one of the factors associated with this phenotype.
Introduction
T
In addition to causing disease in chickens, it has been hypothesized that APEC may also play a role in human disease and constitute a potential zoonotic risk, mainly associated with urinary tract infections and neonatal meningitis, due to the presence of similar virulence genes and phylogenetic backgrounds as well as sharing serotypes among some APEC and human ExPEC strains (Moulin-Schouleur et al., 2007; Bélanger et al., 2011; Mellata, 2013). Furthermore, there are experimental evidences that some human ExPEC strains are able to produce disease in chickens (Moulin-Schouleur et al., 2007; Bauchart et al., 2010).
The likelihood that E. coli from poultry may cause intestinal disease in other animals, and vice versa, is unclear. However, there is some evidence for this possibility. Recently, an E. coli strain isolated from a calf with diarrhea was found to be able to cause colibacillosis in poultry (Matsuda et al., 2010). Furthermore, a previous study found that the production of enterotoxins is unusual in APEC, but heat-labile toxin (LT) could be detected (Blanco et al., 1997). Also, enteroaggregative E. coli, a diarrheagenic pathotype, was associated with an outbreak of an extraintestinal disease (urinary tract infection) (Olesen et al., 2012). These findings led us to speculate that some APEC strains may possess virulence determinants that would allow them to potentially cause enteric disease in mammalians.
To determine whether APEC may cause enteric pathology in the mammalian gut and potentially be an enteric pathogen in certain animal species, APEC strain SEPT362 was selected for testing in an in vivo model (rabbit ligated ileal loop). This strain presents serotype OR:H10 and was isolated from the liver of a layer hen exhibiting clinical signs of septicemia due to colibacillosis and is resistant to streptomycin, tetracycline, and ampicillin. It is also able to adhere to and invade HeLa cells cultured in vitro but has no cytotoxic effects on Vero cells (de Pace et al., 2011).
Materials and Methods
Strains and inoculum preparation
Besides the strain SEPT362, further strains and plasmids used in this work are described in Supplementary Table S1 (Supplementary Data are available online at
In vivo rabbit ligated ileal loop assay
All experiments were approved by the Committee of Ethics on Animal Experimentation (Protocol number 2449-1) of the Institute of Biology, State University of Campinas (UNICAMP). Two New Zealand White male rabbits (2 months old) were used to perform the rabbit ligated ileal loop assay as previously described (Vieira et al., 2010) with slight modifications. A volume of 1.0 mL of each inoculum (bacterial suspension and differently treated supernatants) was inoculated onto each ligated loop using a 0.45×13-mm needle. As a negative control, either 1.0 mL of sterile PBS (pH 7.4) or LB medium was used. Eighteen hours after the surgery, the animals were sacrificed. The increase in the size of each ligated loop due to fluid secretion was assessed visually and scored: high (++++), medium (+++-), weak (++--), very weak (+---), and negative (----). The presence or absence of blood in the secreted fluid was also noted.
Light microscopy
The entire content of each ileal loop was fixed by the injection of a solution of 4% paraformaldehyde and 1% glutaraldehyde dissolved in 0.1 M sodium cacodylate buffer (pH 7.2). Subsequently, the ileal loop was opened, and fragments were excised and immediately immersed in the same fixative solution for 24 h. The tissues were dehydrated in graded concentrations of alcohol and embedded in historesin (Leica Microsystems) and paraffin. The historesin-embedded samples were serially sectioned (2 μm) and stained with hematoxylin phloxine.
Scanning electron microscopy (SEM)
For electron microscopy, small tissue fragments fixed in the 4% paraformaldehyde and 1% glutaraldehyde solution, as previously described, were postfixed in 1% osmium tetroxide for 1 h at 4°C. The samples were dehydrated with a graded series of ethanol concentrations and then dried to the critical point with CO2 in a Balzers CPD 030 critical point dryer. The specimens were coated with gold in a Balzers SCD 050 sputter coater and then examined with a JEOL 5800 LV SEM at 5–10 kV.
Search for virulence genes in silico
The SEPT362 genome sequence (Rojas et al., 2013) was downloaded from GenBank (accession no. AOGL00000000). The BLAST tool was used on a local platform against a local database. The software blast 2.2.26 with the application tblastn was used to search for known E. coli genes (lt, stp, sth, stx1, stx2, astA, tsh, sat, pic, vat, sigA, pet, sepA, eatA, espP, espC, ecp, fimH, lpfA1-1, lpfA1-2, lpfA1-3, lpfA1-4, lpfA1-5, lpfA2-1, lpfA2-2, lpfA2-3, csgA, sfaD/E, papC, afa, ral, cdt, cnf-1, and cnf-2). The software was executed with the default parameters except for the e-values that were adjusted to 1. The predicted protein was considered as present in SEPT362 genome if the identity percent was >90.
PCR for virulence-related genes
PCR was performed as described previously (Borges et al., 2012). Primers are shown in Supplementary Table S2. All control strains were obtained from the Bacterial Molecular Biology Laboratory except strains FVL16 and FVL35, which were kindly supplied by Dr. D.S. Leite, and strain PCAY, which was kindly provided by Dr. T. Yano from the State University of Campinas.
Results
APEC strain SEPT362 produces an enterotoxigenic-like activity in the rabbit ligated ileal loop assay
The inoculation of rabbit ligated ileal loops with 109 CFU/mL of strain SEPT362 resuspended in PBS resulted in abundant viscous fluid accumulation (++++) at 18 h postinoculation (Fig. 1A and B2). Additionally, the filtered (0.22 μm) supernatants of bacteria grown overnight also caused the same remarkable fluid accumulation in ileal loops (++++) (Fig. 1C). The loops inoculated with filtered supernatants treated with heat at 65°C still accumulated fluid (++++), although this effect was significantly compromised (+---) following inactivation with heat at 100°C (Fig. 1C). Moreover, the filtered supernatant, which was further filtered with 3000, 10,000, 30,000, and 100,000 MWCO membranes, induced no fluid accumulation in the loops (----) (Fig. 1C). These results indicate that (1) the strain SEPT362 causes liquid accumulation in a mammalian ileal loop; (2) the observed liquid accumulation could be due to a substance or substances that are either produced and secreted by the bacterial cell or released after bacterial lysis and are thus present in the bacterial supernatant; (3) the liquid has a very viscous aspect and could be released by the intestinal cells; (4) the secondary filtration of any of the supernatants through different MWCO membranes eliminated liquid accumulation in the ileal loop assay; and (5) heat treatment (100°C, 15 min) remarkably decreased the level of fluid production. These results suggest that the substance(s) that produce(s) this effect is (are) partially heat-resistant at 65°C and heat-sensitive at 100°C with a theoretical size >100 kDa.

Rabbit ligated ileal loops inoculated with APEC SEPT362 or phosphate-buffered saline (PBS) after 18 h and lacking bacterial colonization.
APEC SEPT362 harbors the genes encoding for the enterotoxin astA and the autotransporter tsh
The in silico blast analysis was used to identify several genes in strain SEPT362. To confirm the in silico result, the gene was further tested by PCR. Then, the in silico and PCR analysis showed that SEPT362 did not harbor the genes encoding known E. coli toxins LT-I (heat labile), STIa (STh) and STIb (STp) (heat stable), or Stx1 and Stx2 (Shiga toxins 1 and 2). However, SEPT362 possessed DNA sequences encoding for the EAST-1 toxin (enteroaggregative heat-stable toxin, encoded by astA). APEC SEPT362 also harbored the serine protease autotransporter (SPATE) gene tsh but not sat, pic, vat, sigA, pet, sepA, eatA, espP, or espC. This strain was also negative for cnf-1, cnf-2, and cdt-I genes.
Regarding the presence of possible adhesins, SEPT362 contained sequences corresponding to type 1 fimbriae ( fimH), curli (csgA), long polar fimbria O44 (lpfA2-3) genes and the ECP (E. coli common pilus) operon but did not contain lpfA1, and the remaining lpfA2 types, S fimbria adhesin (sfa), afimbrial adhesin (afa), or P fimbrial (papC) genes, or the REPEC (rabbit-specific enteropathogenic E. coli) adherence locus (ral).
APEC SEPT362 induced effacement of the enterocyte brush border
The light microscopy results showed that there were no bacterial cells adhering to the intestinal villi, and the primary histopathological changes were found mainly at the villus tip (Fig. 2C and D). It was possible to detect a discontinuous disruption at the apical domain of the enterocytes that resulted in effacement of the eosinophilic brush border. The lateral and basal cellular domains showed no apparent rupture of the boundaries, which appeared clearly delimited by tightly bound junctional complexes. In addition, the villi cores exhibited well-preserved lamina propria. In contrast, microscopic examination of histological sections of the intestine loops inoculated with PBS and LB (controls) revealed no effacement of the eosinophilic brush border (Fig. 2A and B).
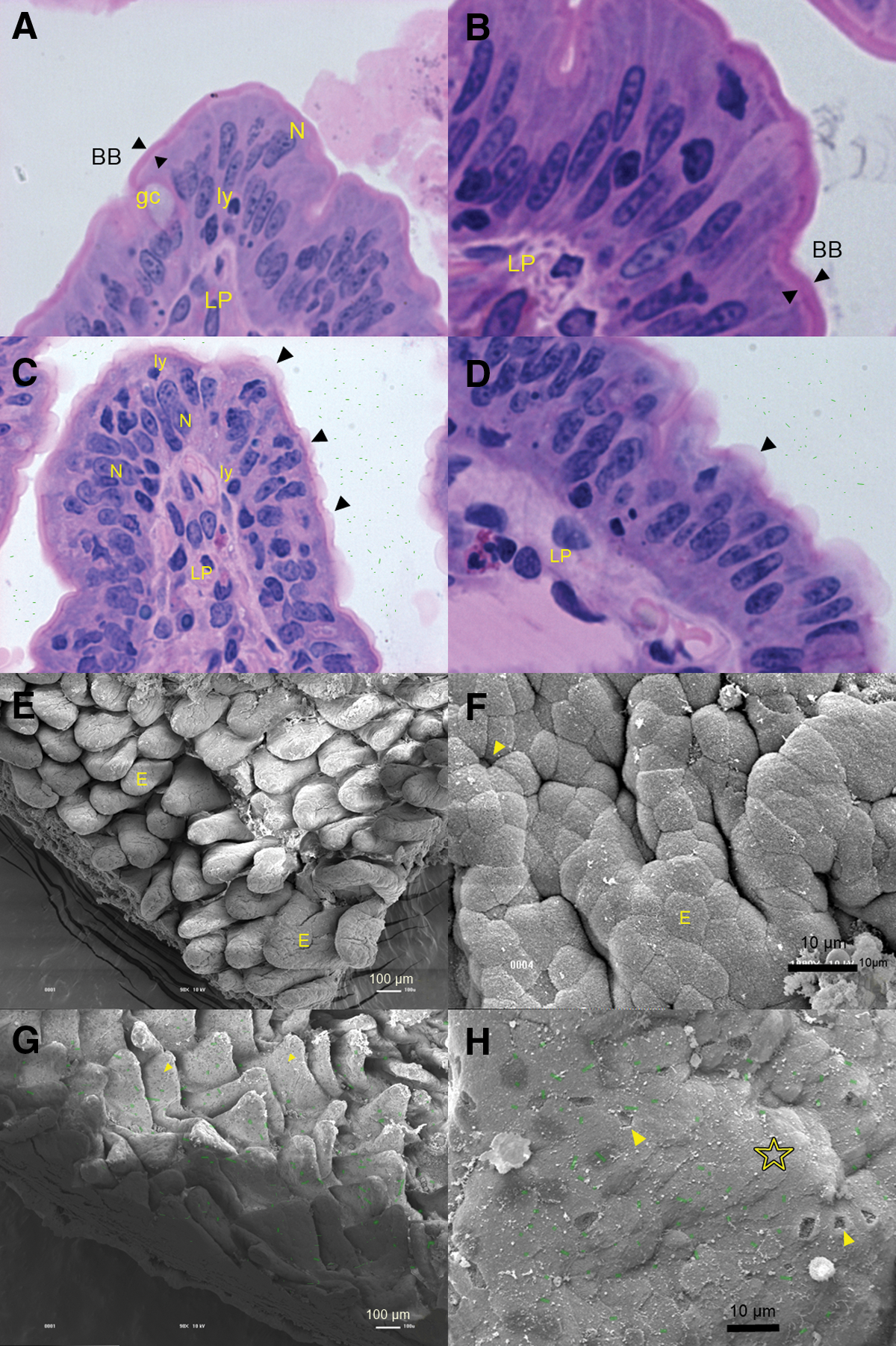
Microscopy images of the intestinal villi of loops inoculated either with phosphate-buffered saline (PBS) (control) or SEPT362 showing the lack of adhesion (bacterial cells were artificially depicted in green).
SEM demonstrated that the bacteria caused tissue damage
The SEM results confirmed that strain APEC SEPT362 was unable to adhere to and colonize the intestinal villus (Fig. 2G and 2H) and was washed off during the preparation of the samples for microscopy. Although there was no bacterial colonization of the intestinal cells, there were detached epithelial cells all over the villi (Fig. 2G and H). The villi also had a flattened and wilted appearance (Fig. 2G), in contrast to the loops inoculated with PBS (Fig. 2E and F).
The presence of tsh gene conferred the enterotoxin effect in the rabbit ileal loop
Based on the data obtained from screening for the presence of potential virulence genes in strain SEPT362 and size exclusion, which indicated a product in the supernatant with a molecular mass exceeding 100 kDa, the possibility that the Tsh autotransporter could confer the enterotoxic phenotype was investigated. Supernatants and bacterial cultures of the APEC reference strain χ7122, an isogenic mutant of χ7122 (χ7273), as well as a cloned copy of tsh in an E. coli K12 strain were tested in the ileal loop model. Results demonstrated that APEC χ7122 also caused the enterotoxic effect (Fig. 3A), although loss of Tsh in this strain did not completely abrogate fluid accumulation (Fig. 3B). This suggests that Tsh may be the predominant protein conferring enterotoxic activity to strain χ7122, but that other components may also contribute to enterotoxicity. In strain χ7122, it is likely that in addition to Tsh, other factors mediating enterotoxicity are encoded on plasmid pAPEC-1, since χ7273 cured of pAPEC-1 showed a decreased fluid accumulation (Fig. 3C). To more directly determine whether Tsh can confer an enterotoxic effect, an E. coli K-12 strain containing a cloned copy of tsh was tested (Fig. 3D). The E. coli K-12 strain containing tsh was also able to induce fluid accumulation, whereas the K-12 control strain was negative for enterotoxicity in the rabbit ileal loop (Fig. 3E). Taken together, these results support a potential and novel feature of the Tsh autotransporter: the potential to induce fluid accumulation and features of enterotoxicity.
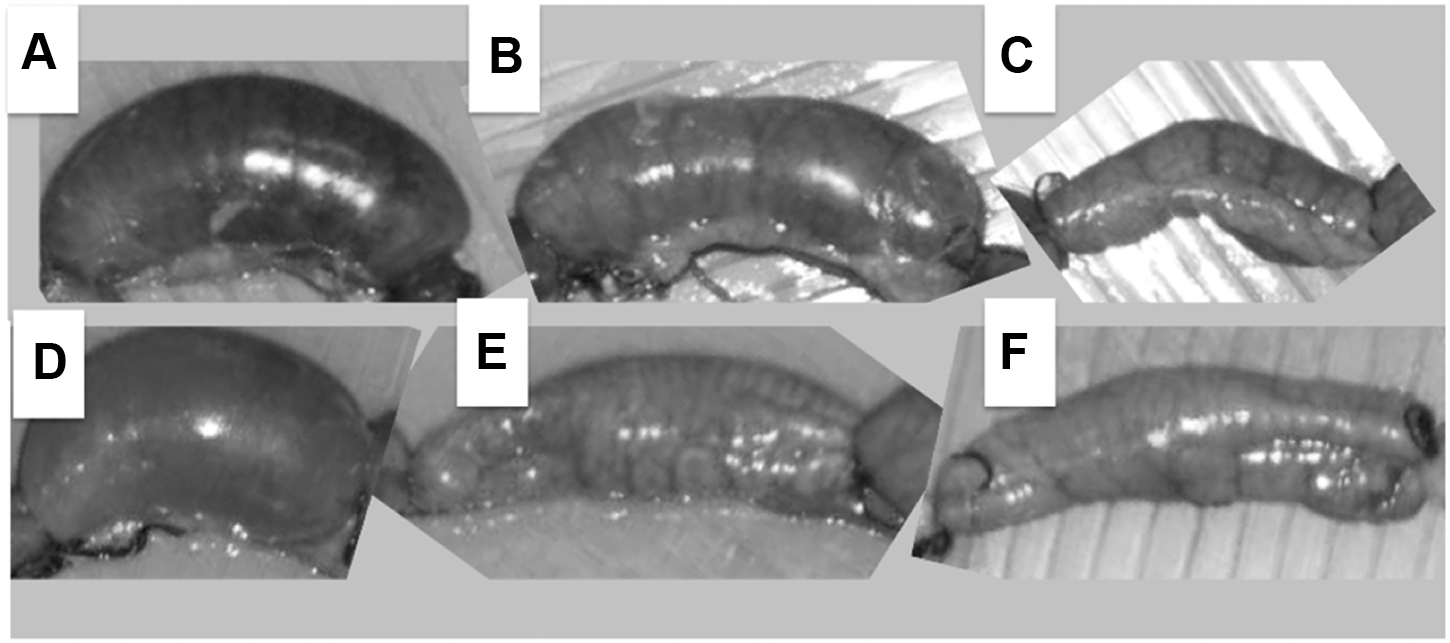
Rabbit ileal loop assay using the following samples:
Discussion
In this study, we investigated the in vivo intestinal activity of an APEC strain in a mammalian host (rabbit) and demonstrated that this strain exhibits an enterotoxigenic-like activity, with the production of viscous liquid and histopathological alterations in the ileal loops. Furthermore, the Tsh autotransporter was shown to be associated to this enterotoxin-like phenotype, as a clone presenting tsh in E. coli K-12 produced identical effects compared to wild-type APEC strains. However, inactivation of the tsh gene in APEC reference strain χ7122 did not completely abrogate enterotoxinogenic properties, suggesting that other components may also contribute to fluid accumulation and enteric pathologies, at least with this strain. To the best of our knowledge, this is the first report demonstrating enterotoxic properties in an APEC strain.
The microscopy data presented herein showed that the extraintestinal strain SEPT362 was not able to colonize the mammalian intestine. However, E. coli strains harboring ExPEC-related genes (Dr hemagglutinin and P-, type 1-, and S-fimbrial adhesins) have been shown to colonize colonic and ileal human enterocytes in cell cultures (Adlerberth et al., 1995). Furthermore, a single E. coli clone (containing a typical ExPEC gene repertoire) dominated the coliform gut microbiota of healthy pigs (Schierack et al., 2008), possibly due to in vivo colonization of the intestine. These facts indicate that some ExPEC-associated virulence genes could support intestinal colonization. Apparently, if expressed, the adhesion factors encoded by fimH, lpfA2-3, cgsA, and ECP detected in this study do not contribute to gut colonization by an extraintestinal strain in this in vivo model. This strain has been shown to adhere to and invade HeLa cell monolayers in vitro, a characteristic that the cited factors could be associated with de Pace et al. (2011). However, the factors do not seem to be associated with effects on Vero cells in vitro, once the strain did not adhere or invade these cells. This contrast could be due to the presence of specific receptors in HeLa cells that might be absent in Vero cells.
Although the strain SEPT362 was not found to colonize the small intestine, its potential to cause enteric disease in mammalians cannot be discounted. This strain or indeed other APEC strains that may encode enterotoxin-like activity due to Tsh or other factors may successfully colonize the intestine. Also, as tsh is typically located on conjugative ColV or similar plasmids, transfer of the plasmid encoding the putative factor(s) associated with the liquid accumulation in the intestine to bacterial strains that readily colonize the gut could take place.
The microscopy results, showing the irregular surface of the intestinal villi with an increased number of detached cells, goblet secretion, and the brush border effacement produced by SEPT362 in the rabbit ileal loops, also favor this gut pathogen potential hypothesis. Because the rabbit ligated ileal loop assay is a closed system, we speculate that the damages might have appeared due to the accumulation of a bacterial product, either secreted or released after bacterial lysis, even without bacterial adhesion to the rabbit enterocytes.
The biological effect demonstrated here does not appear to be caused by EAST-1 toxin, which is encoded by astA and is able to lead to an enterotoxigenic effect in vitro (Savarino et al., 1991). This conclusion is due to the low molecular weight (MW) (4104.0 Da) of EAST-1 (Ménard et al., 2004), which contrasts with the high MW (>100 kDa) of the component associated with the biological effect produced by this strain.
The temperature-sensitive hemagglutinin (Tsh) was the only SPATE detected in this work. This SPATE has MW >100 kDa (Provence and Curtiss, 1994) and SPATEs have been associated with increased secretion by intestinal cells (Navarro-Garcia et al., 2010). Furthermore, in Vibrio cholerae, a serine protease plays a role in an hemorrhagic effect in the rabbit ileal loop model (Syngkon et al., 2010). Indeed, the results demonstrated herein represent the first indication of a possible role for Tsh as a potential enterotoxin in the mammalian gut.
Conclusions
The results of this work suggest that avian extraintestinal E. coli might be able to cause fluid accumulation in the rabbit gut. The Tsh autotransporter seems to be one of the factors associated with this phenotype. Further studies are necessary to confirm this new role of Tsh and the hypothetical role of avian extraintestinal E. coli in mammalian enteric diseases.
Footnotes
Acknowledgments
In Brazil, this work was funded by FAPESP (2012/05073-3, 2010/50596-9, 2010/51421-8 and 2012/04931-6), CNPq (142718/2010-6), and CAPES. In Canada, it was funded by a Discovery grant from NSERC (RGPIN 250129-07) and a Canada Research Chair.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.