
Abstract
The aim of this study was to determine the molecular subtyping and antimicrobial susceptibility characteristics of Campylobacter coli isolates from different sources in China. One hundred thirteen C. coli isolates were subtyped by pulsed-field gel electrophoresis (PFGE), multilocus sequence typing (MLST), and porA and flaA short variable region (SVR) nucleotide sequences. Cluster analysis was performed based on the PFGE and sequence types (ST). Eighty-four PFGE patterns (SmaI) were observed in 113 isolates. Fifty-four STs (28 novel) and three clonal complexes (CC), 86% of which were clustered to CC828, were observed, as well as 52 porA and 37 flaA-SVR sequence alleles. MLST, porA, and flaA-SVR analysis demonstrated that many isolates from diarrheal patients shared identical genotypes with chicken isolates. Minimum inhibitory concentration values of 10 antibiotics were analyzed for 109 isolates isolated in 2011 using the E-test method. The most frequently observed resistance agents were nalidixic acid (100%) and ciprofloxacin (100%), followed by levofloxacin (99%), tetracycline (94%), metronidazole (93%), erythromycin (61%), streptomycin (72%), gentamicin (59%), ampicillin (50%), and chloramphenicol (29%). Multidrug resistance was detected in 108 of 109 C. coli isolates (99%).
Introduction
C
Bacterial subtyping schemes play an important role in epidemiological investigations to trace sources and routes of pathogen transmission by identifying the outbreak and differentiating the sporadic infection (Schweitzer et al., 2011; O'Mahony et al., 2011). Multilocus sequence typing (MLST) and pulsed-field gel electrophoresis (PFGE) have been employed successfully for the longitudinal epidemiology study of Campylobacter spp. from different sources and outbreak investigations (Dingle et al., 2001; Hume et al., 2002; Dingle et al., 2005; Michaud et al., 2005; Thakur et al., 2009).
In recent years, a number of studies have indicated an increase in emerging drug-resistance characteristics among Campylobacter spp. (Kos et al., 2006; Gibreel et al., 2006; Alfredson et al., 2007). Campylobacter can be transferred from food-producing animals to humans, and many reports have demonstrated that the drug-resistant strains involved in human disease are predominantly spread through the food chain (Smith et al., 1999; Piddock et al., 2000; Engberg et al., 2004; Smith et al., 2010; Schweitzer et al., 2011). Determination of the drug resistance status of Campylobacter strains from different sources is crucial for the control and prevention of human infection, particularly for the cases in which therapy is recommended.
In order to determine the genotype and drug resistance profiles of C. coli from China, antibiotic susceptibility tests and molecular subtyping analysis of isolates from different sources were carried out in this study.
Materials and Methods
Isolates collection
Molecular typing was performed on 113 isolates that were isolated from 2 regions (43 isolates from Beijing and 70 isolates from Shanghai) in China from 2010 to 2011. Among these, 48 were from the stool samples of diarrheal patients collected during April to October in each year, 32 were isolated from retail foods (27 from fresh chicken meat and 5 from raw pork), and 33 were from fecal samples obtained from food-producing animals during the same time and in the same region (18 from chickens and 15 from swine). Antimicrobial susceptibility tests were performed on 109 isolates that were isolated in 2011 in Shanghai. Among these, 61 isolates were from the stool samples of diarrheal patients collected from 11 hospitals and 48 isolates were obtained from the fecal samples of food-producing animals in the same region (27 from swine stools collected from 3 abattoirs and 21 from chicken stools collected from 5 poultry markets). In 51 of the 109 isolates, subtyping analysis was also performed. All of the tested isolates were randomly selected from the collection of a pilot study on Campylobacter infection and transmission supported by the Major State Basic Research Development Program in China.
Bacterial and species identification was performed by Gram staining, hippurate hydrolysis tests, and C. coli-specific polymerase chain reaction (PCR), according to previous reports (Klena et al., 2004). The bacteria were cultured on Skirrow's medium (Columbia Agar Base supplemented with 5% sheep blood) at 37°C for 48 h in a microaerobic atmosphere (5% O2, 10% CO2, and 85% N2). The genomic DNA of each isolate was extracted using a DNeasy tissue kit (Qiagen, Valencia, CA) according to the manufacturer's protocol.
PFGE
PFGE was performed according to the standard PulseNet PFGE protocol for C. coli (
MLST
MLST was performed by sequencing seven housekeeping gene loci (aspA, glnA, gltA, glyA, pgm, tkt, and uncA) according to previously described methods (Dingle et al., 2005). The allele of each gene was obtained from the comparison of the seven alleles in the MLST database (
FlaA–short variable region (SVR) and PorA sequencing
PCR amplification of flaA-SVR was performed using modified forward and reverse primers flaA-SVR-F (5′-TCCGCAGCAGATGATGCTTCA-3′) and flaA-SVR-R (5′-GCTCTAAC[A,T]CCTGTTTTATC-3′). Nucleotide sequence extension reactions were carried out on both DNA strands using the flaA-SVR primer pairs. The flaA-SVR allele numbers were assigned by comparing the newly generated sequences against existing ones (
Phylogenetic analysis based on MLST
The most likely ST founder of the 113 isolates was determined using eBURSTv3 software (Feil et al., 2004). The stringent group definition was based on isolates with six matches to the seven housekeeping gene alleles placed in the same complex and a bootstrapping method (resampling 1000 times) was conducted to provide statistical confidence in the assignment of the ST as the putative founder of the complex. Minimum spanning tree analysis was used to infer the relationships of the genetic population among the isolates from different sources and was constructed using BioNumerics based on the STs and the sequences of porA and flaA-SVR of the isolates.
Antimicrobial susceptibility testing
The minimum inhibitory concentrations (MICs) of 10 antibiotics (erythromycin, streptomycin, gentamicin, ampicillin, chloramphenicol, tetracycline, metronidazole, nalidixic acid, ciprofloxacin, and levofloxacin) against 109 C. coli isolates were determined using an E-test (AB Biodisk) method, as previously described (Zhang et al., 2010). The breakpoint values for erythromycin, ciprofloxacin, and tetracycline used in this study were set as ≥32 μg/mL, 4 μg/mL and 16 μg/mL, respectively, based on Clinical and Laboratory Standards Institute (CLSI, 2006). The breakpoint value for ampicillin, chloramphenicol, gentamicin, and streptomycin used in this study was that the MIC ≥16 μg/mL and the resistance breakpoints for nalidixic acid, levofloxacin, and metronidazole were set as MICs ≥32 μg/mL, 4 μg/mL, and 1 μg/mL, respectively, according to previous publications (Luangtongkum et al., 2006; Miflin et al., 2007; Marinou et al., 2012). C. jejuni ATCC 33560, Staphylococcus aureus ATCC 29213, Escherichia coli ATCC 25922 and the erythromycin-resistant C. coli strain WHO C-9.1 (MIC 32 μg/mL) from the External Quality Assurance System of the WHO Global Foodborne Infections Network were established as erythromycin-resistant controls.
Statistical analysis for antimicrobial resistance rates
The frequencies of antimicrobial resistance patterns from different sources were compared using a χ2-test and Fisher's exact two-tailed test with SPSS19.0 software, where applicable. Values of p<0.05 were considered statistically significant.
Results
PFGE typing
In total, 84 PFGE patterns were detected among the 113 examined isolates. Five closely related groups, sharing >75% similarity in banding patterns, were found for 80 of the tested isolates (Fig. 1). Isolates from different sources were distributed across various clusters. The multidrug-resistant isolates, which are marked with black circles, presented in different clusters (Fig. 1).

Dendrogram of pulsed-field gel electrophoresis (PFGE) cluster analysis of 113 Campylobacter coli isolates from different sources. PFGE was performed according to the standard PulseNet PFGE protocol for C. coli with the restriction enzyme SmaI. Cluster analysis was performed using BioNumerics software (Version 4.1, Applied Maths, and Austin, TX). Five clusters were found with ≥75% similarity in banding patterns. Multidrug-resistant isolates (marked with a black circle on the left side of the isolate ID) were distributed in different clusters.
Sequence typing and genetic relation
The background, STs, porA, and flaA-SVR are summarized in Table 1. A total of 54 STs, including 28 novel types, were observed from the 113 isolates. Three clonal complexes (CC)—CC21 (1 isolate), CC1150 (2 isolate), and CC828 (97 isolate)—were generated from 100 isolates; 12 STs from 13 isolates could not be assigned to any of the defined CCs. Twenty-six STs were observed from 48 human isolates. The most frequently observed ST among the human isolates was 1625 (19%, 9/48), followed by 1145 (13%, 6/48). The most frequently observed STs among the chicken fecal or chicken meat isolates were 829 (11%, 5/45) and 860 (11%, 5/45), followed by 1625 (9%, 4/45) and 855 (9%, 4/45). The most frequently observed ST among the swine or pork isolates was 1464 (20%, 4/20). Thirty-four STs were observed in 51 multidrug-resistant isolates. Two CCs—CC1150 (1 isolate) and CC828 (43 isolate) —were generated from 44 isolates; 7 STs from 7 isolates could not be assigned to any of the defined CCs. No specific STs or CCs were related to the drug-resistant profile.
Fifty-two porA alleles were detected from 113 isolates. The four most frequently presented alleles were 1427 and 687, followed by 1428 and 915, which are mostly from chicken or human isolates. Allele 1716 was generated from five swine isolates. Thirty-seven flaA-SVR sequence variants were found and the most common allele, 16, was generated from 23 isolates, 16 of which were from diarrheal patients. The next most frequently observed allele was 17, which is predominantly from diarrheal patients or chicken isolates.
eBURST analysis indicated that the most likely founder ST of the 113 isolates in this study was ST860. ST825, ST829, ST1055, ST1625, and ST828 were the subfounders (Fig. 2). The genetic relatedness based on the STs and the alleles of porA and flaA-SVR from the 113 isolates is presented in Figure 3. The isolates from diarrhea patients showed identical or similar genotypes (with more than seven genes identical) with the isolates from chicken.
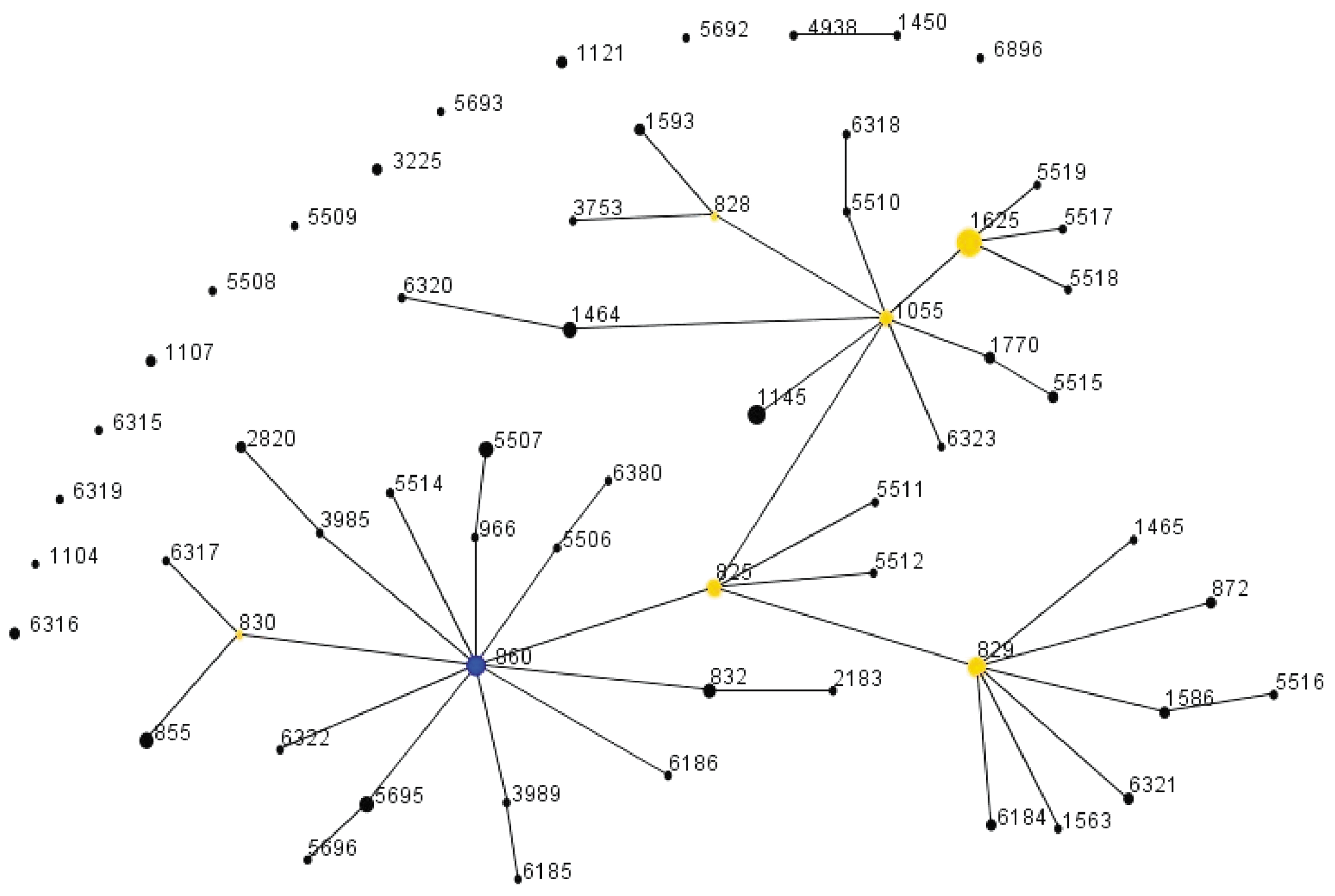
eBURST analysis for 113 Campylobacter coli isolates. The sequence types (ST) founders of 113 isolates were determined using eBURSTv3 software. The stringent group definition was based on isolates with six matches to the seven housekeeping gene alleles placed in the same complex, and the bootstrapping method (resampling 1000 times) was conducted to provide statistical confidence in the assignment of the ST as the putative founder of the complex. The size of the circle is proportional to the number of isolates with the indicated ST. The most likely founder ST of the 113 isolates in this study is ST860 (blue). ST825 (yellow), ST829 (yellow), ST1055 (yellow), ST1625 (yellow) and ST828 (yellow) are the subfounders.
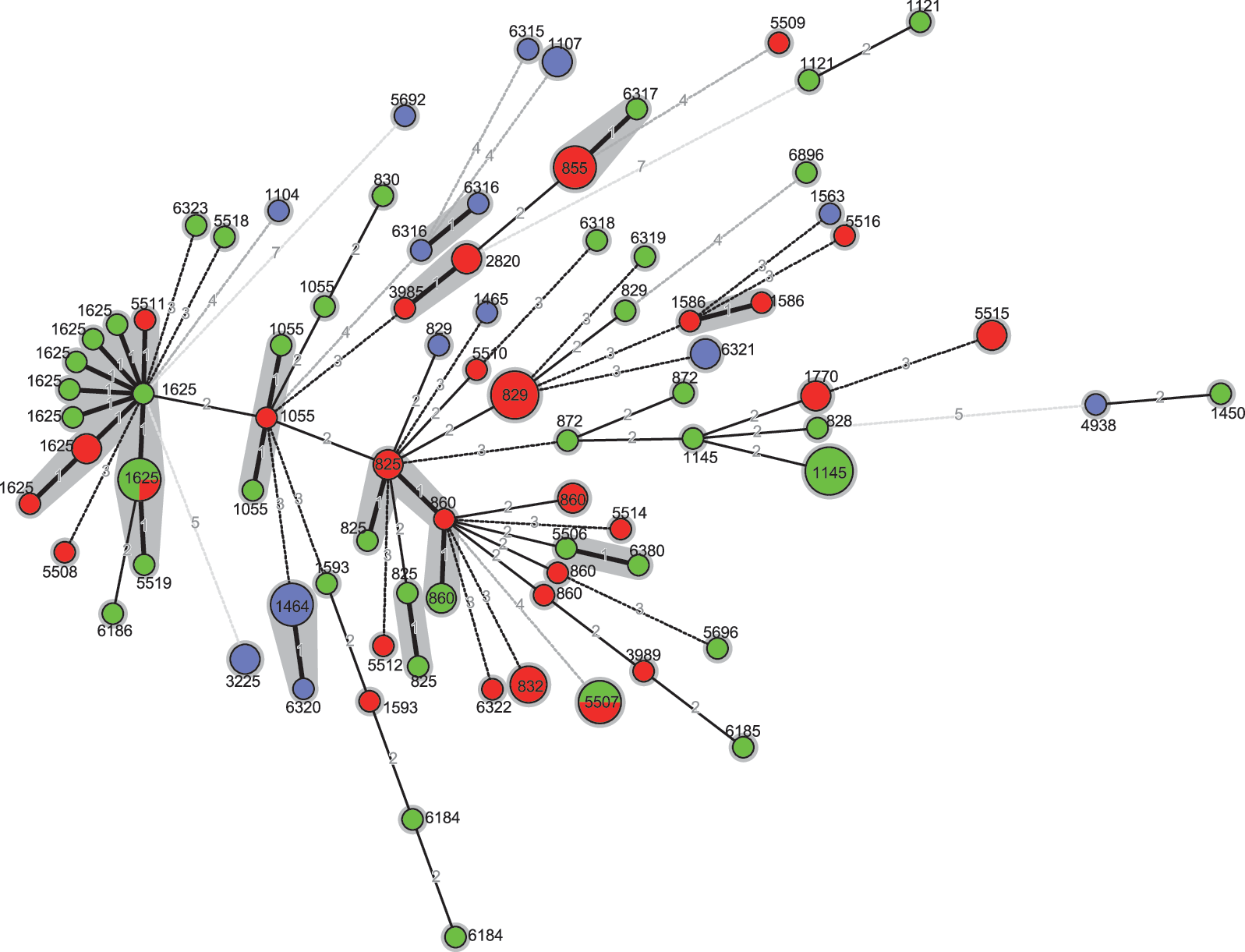
The genetic relatedness of 113 Campylobacter coli isolates from different sources. The minimum spanning tree was constructed using the alleles of porA, flaA-SVR, and sequence types (ST) from 113 Campylobacter coli isolates from different sources (chicken, red; diarrheal patient, green; swine, purple). The size of the circle is proportional to the number of isolates with the indicated ST. The numbers along the lines indicate the difference of the alleles between different isolates (active line: one or two alleles different; dotted line: more than two alleles different). The isolates from diarrheal patients show a more similar genotype to the isolates from chicken.
Results of antimicrobial susceptibility tests
The MIC50 and MIC90 values for each antibiotic are shown in Table 2. The antibiotic-resistance ratio and the statistical analysis for each drug on isolates from different sources are shown in Table 3. Nalidixic acid and ciprofloxacin resistance were found in 100% of C. coli isolates recovered from all sources. Otherwise, the most frequently observed resistance was to levofloxacin (99%, 108/109), tetracycline (94%, 103/109), metronidazole (93%, 101/109), erythromycin (61%, 67/109), streptomycin (72%, 78/109), gentamicin (59%, 64/109), ampicillin (50%, 54/109), and chloramphenicol (29%, 32/109). Multidrug resistance (resistance to 4 or more categories of antibiotics) was observed in 99% (108/109) of isolates.
Examining the isolates from diarrheal patients and the isolates from chickens, the resistance ratios for nine of the antibiotics were found to be similar (p>0.05). Only the resistance ratio of chloramphenicol was noted to be significantly higher in chicken isolates compared to the isolates from diarrheal patients (p=0.0019). However, there are significant differences between the resistance ratios of gentamicin, streptomycin, ampicillin, and metronidazole between the isolates from diarrheal patients and isolates from swine (Table 3). The resistance frequency of swine isolates to streptomycin was higher than that of human isolates (p=0.0081). In contrast, the resistance frequencies of swine isolates to gentamicin, ampicillin, and metronidazole were less than those of the isolates from diarrheal patients. The isolates from diarrheal patients shared similar resistance profiles to the isolates from chickens rather than to those from swine. There was no significant difference between the frequencies of resistance to nalidixic acid, ciprofloxacin, levofloxacin, and tetracycline among the isolates from different sources (p>0.05).
Discussion
C. coli bacteria colonize in the intestines of food-producing animals and can contaminate the associated foods during processing. Consumption of contaminated food has been implicated as the major source of human campylobacteriosis (Mylius et al., 2007; Hermans et al., 2012). Studies from China and other countries have demonstrated that C. coli–associated human campylobacteriosis accounts for approximately 11%–40% of cases (Popovic-Uroic, 1989; Uzunovic-Kamberovic, 2001; Uzunovic, 2003). Molecular subtyping of isolates from the clinical cases and other sources is a potential way to provide information about the source attributes, and source attribution studies can subsequently provide valuable information regarding infection control and disease prevention. The present study found that the majority of the tested isolates (86%) belonged to one lineage (CC828), which infers a common ancestry. This result is consistent with a previous investigation, which indicated that 83.3% of Chinese C. coli isolates from chicken meat belonged to CC828 (Wang et al., 2010).
There have been reports showing that pigs are a common source of C. coli, and they have been previously considered to be the most probable source of human infections (Nielsen et al., 1997; Harvey et al., 1999); however, there are also studies that have demonstrated poultry to be the primary reservoir for both C. jejuni and C. coli bacteria (Siemer et al., 2005; Wingstrand et al., 2006; Zorman et al., 2006). In the present study, both MLST, porA, and flaA-SVR analysis have indicated that the majority of isolates collected from diarrheal patients share identical genotypes with chicken isolates. In addition, the universal C. coli prevalence in chickens also suggests that chickens may be a dominant source of human C. coli infections in China (Wang et al., 2010).
Campylobacter infections are usually self-limiting and do not need antibiotic treatment. However, for severe and prolonged cases, antimicrobial treatment is required. Currently, fluoroquinolones and macrolides are commonly used for the treatment of human campylobacteriosis in China. However, our results indicate that all of the tested isolates are almost 100% resistant to fluoroquinolones and 66% of the human isolates are resistant to erythromycin. The clinical treatment for C. coli-associated campylobacteriosis should therefore be carefully reconsidered. Previous reports have demonstrated that poultry is a major source of fluoroquinolone-resistant Campylobacter-resulting human infections (Iovine et al., 2004a, 2004b; Smith et al., 2010). The judicious use of fluoroquinolone in food-producing animals might be one effective way in which the spread of drug-resistant Campylobacter be reduced in China. It should be noted that the resistance ratios of isolates from diarrheal patients was much higher than has been previously reported (Uzunovic et al., 2013).
The present study indicated the resistant profile of the tested C. coli isolates was quite different from the strains isolated in the United States, which were commonly resistant to tetracycline (66.2%) and less resistant to fluoroquinolone (1.5%) (Thakur et al., 2010). The resistant frequencies were higher than those of isolates from other counties (Lutgen et al., 2009; Dohne et al., 2012; Obeng et al., 2012). High frequencies of multidrug resistance were noted in C. coli isolates from different sources (99%). No specific cluster was found among the 51 multidrug-resistant isolates according to both the PFGE and MLST results. These isolates showed variant patterns, which indicates that they are genetically diverse. The increase in the number of highly resistant C. coli will not only bring complications with regard to disease treatment, but these bacteria will also form a resistant reservoir in the intestines of animals. A resistance integron was recently found in C. coli isolates isolated in China (Qin et al., 2012).
Conclusions
In summary, most C. coli isolates from diarrheal patients shared identical genotypes and antibiotic resistance patterns with isolates from chickens, and high drug resistance was found among isolates from different sources. The data obtained in this study will benefit the established strategy for Campylobacter monitoring and may also improve food safety in China and other parts of the world.
Footnotes
Acknowledgments
This work was supported by the Major State Basic Research Development Program (2013CB127204) and the General Program of National Natural Science Foundation of China (81271789).
Disclosure Statement
No competing financial interests exist.