
Abstract
Four mushroom (Flammulina velutipes) production plants were sampled to investigate the prevalence and contamination source of Listeria monocytogenes. Among 295 samples, the prevalence of L. monocytogenes was 18.6%; the contamination appeared to originate from the mycelium-scraping machinery, contaminating both the product and upstream packaging equipment. Of 55 L. monocytogenes isolates, lineages I.1 (1/2a-3a) and II.2 (1/2b-3b-7) accounted for 65.5% and 34.5%, respectively. In addition, lineage I.1 formed significantly thicker biofilms than those within lineage II.2, as determined by crystal violet staining and scanning electron microscopy. Genotype analyses of L. monocytogenes isolates using enterobacteria repetitive intergenic consensus-polymerase chain reaction, and random amplified polymorphic DNA revealed that the surfaces of mycelium-scraping machinery may serve as the main source of L. monocytogenes contamination in three of the four plants. This study was the first report to explore the potential contamination sources of L. monocytogenes in the mushroom production chain, thereby providing baseline information for adopting prophylactic measures for critical control points during production in mushroom plants to avoid L. monocytogenes contamination.
Introduction
L
In recent years, the prevalence of Listeria spp. in mushrooms has been reported in several countries (Samadpour et al., 2006; Cordano and Jacquet, 2009; Venturini et al., 2011). Although no listeriosis case associated with the consumption of fresh mushroom has been reported, several developed countries have formulated standards and policies to minimize the risk of foodborne listeriosis. Some recalls have even occurred because of the contamination of L. monocytogenes in mushroom products (Canadian Food Inspection Agency, 2011, 2012; U.S. Food and Drug Administration, 2012). In addition, a listeriosis case caused by salted mushroom heavily contaminated with L. monocytogenes has been reported in Finland (Junttila and Brander, 1989). We previously found that a ready-to-eat vegetable product containing fresh mushrooms (Fammulina velutipes) contained high numbers of L. monocytogenes (unpublished data). The contamination routes of L. monocytogenes in the mushroom production industry must be elucidated to prevent final products from being contaminated with L. monocytogenes. However, little information is available about the contamination sources of L. monocytogenes in mushroom plants.
The aim of the present study was to investigate the prevalence of Listeria spp. and explore the potential contamination sources of L. monocytogenes in mushroom plants.
Materials and Methods
Sampling and isolation of Listeria spp.
Four mushroom plants (hereafter denoted as A, B, C, and D) were investigated in this study. Plants A and C were located 15 km apart. Plants A(C), B, and D were located approximately 200 km apart from one another. Samples were collected from different phases of production including composting (compost, n=20); the inoculation room (solid spawn, sterile compost, inoculating machinery surfaces, and inoculation room air, n=76); the breeding and mycelium stimulation machinery (n=60); the growing room (atomized water, shelf surfaces, mushrooms, and air, n=83), and the harvesting room (package machinery surfaces, scales, conveyor belts, and packaged mushrooms, n=56). Mycelium stimulation involves the scraping and smoothing of the fungal mycelium in the neck of a culture bottle prior to fruiting body development and is referred to as the mycelium-scraping equipment. For environmental samples, sampling sites were chosen based on both direct contact and proximity to the mycelium or product. This included the air and surfaces of the inoculating machinery, mycelium-scraping machinery, shelves, conveyor belts, scales, and packaging machinery. Each sampling spot in the production environment was swabbed with nine sterile cotton swabs moistened with saline solution (0.85%, wt/vol). The cotton swabs were then placed in 225 mL of Listeria enrichment broth (Huankai, Guangzhou, China). Air sampling was carried out using the gravity-plate method. Five Listeria-selective agar plates (CHROM-agar, Paris, France) were placed open in each room for 30 min.
All samples were transported to the laboratory on ice in an insulated container and tested within 4 h after receipt. Enrichment and isolation procedures were performed as previous reported (Chen et al., 2014). For each positive sample, three to five presumptive colonies were confirmed as L. monocytogenes, using the Microgen ID Listeria identification system (Microgen, Camberley, UK). One of the confirmed L. monocytogenes isolated from each positive sample was submitted for further analysis.
Serotyping
The serotypes of 55 L. monocytogenes isolates and five reference strains (Supplementary Table S1; Supplementary Data are available online at
Enterobacteria repetitive intergenic consensus (ERIC)-PCR typing and random amplified polymorphic DNA (RAPD) analyses
Genomic DNA was extracted from L. monocytogenes strains using a Bacterial Genomic DNA Purification Kit (Dongsheng Biotech, Guangzhou, China) according to the manufacturer's instruction. ERIC primers (ERIC forward: 5′-ATGTAAGCTCCTGGGGATTCAC-3′, ERIC reverse: 5′-AAGTAAGTGACTGGGGTGAGCG-3′) were designed as described by Versalovic et al. (1991). ERIC-PCR typing was performed using the protocol described by Chen et al. (2014). The 10-mer primer UBC-155 (5′-CTGGCGGCTG-3′) was designed and tested in a previous study (Farber and Addison, 1994). The PCR reaction condition was carried out on the 55 L. monocytogenes isolates and 5 reference strains using the protocol described by Chen et al. (2014).
Cluster analysis was performed with NTSYS-pc (version 2.10), a numerical taxonomy and multivariate analysis software package (Rohlf, 2000), based on Dice's similarity coefficient (S D), with 1% position tolerance. The unweighted pair group method was also performed using arithmetic averages.
Quantitation of biofilm formation
Cells attached onto the well walls were quantified as previously described (Djordjevic et al., 2002), with minor modifications. Six wells per strain were filled with 150 μL of 1% (vol/vol) overnight culture of the bacterium in trypticase soy broth (TSB), and each microtiter plate included six wells with sterile TSB in control wells. The microtiter plates were tightly sealed and incubated at 25°C for 72 h. After incubation, the medium was removed from each well, and the plates were washed four times with sterile distilled water to remove loosely attached cells. The plates were then stained with 1% (wt/vol) aqueous crystal violet solution for 45 min and washed four times to completely remove unbound crystal violet. Thereafter, 150 μL of 95% (vol/vol) ethanol was added to each well, and the optical density (OD) at 595 nm was determined. The average OD from the control wells was subtracted from the OD of the test wells. The microtiter plate biofilm assays were performed three times. Comparison of biofilm formation between Listeria lineages was analyzed using one-way analysis of variance followed by a Student-Newman-Keuls post-hoc test with the level of significance of p<0.05.
Biofilm formation assay by scanning electron microscopy (SEM)
Six strains of L. monocytogenes strains were selected for this assay according to the capacity of biofilm formation as determined by crystal violet, namely, ATCC19115, ABMS-1, AGR-2, AHR-4, DBMS-3, and DGR-1. Glass discs (Φ 8 mm, WHB Biotech, Shanghai) were placed in a 48-well plate. Each glass disc was covered with 2 mL of inoculum at 107 colony-forming units/mL in TSB. After 72 h of incubation at 25°C, the growth medium was removed and the discs were washed three times with 5 mL of sterile distilled water. Thereafter, the discs were fixed with 3% glutaraldehyde for 4 h, dehydrated with gradient ethanol solution, dried with a freeze dryer (Hitachi ES2030, Japan), and coated with gold–palladium (Hitachi E1010, Japan). SEM images were taken using a S-3000N Hitachi scanning electron microscope.
Results
Prevalence of L. monocytogenes and Listeria spp. in tested samples
Table 1 shows that among the 295 samples collected from the 4 production plants, 55 (18.6%) samples were positive for L. monocytogenes (i.e., the contaminated sites including compost, surfaces of mycelium-scraping machinery, mushrooms, surfaces of package machinery, scales, products conveyor belts, and packaged mushrooms). No Listeria spp. were found from air samples. Nonpathogenic Listeria spp. (33.9%) exhibited higher frequency in the collected samples. No Listeria seeligeri and Listeria ivanovii were recovered in Flammulina velutipes plants. No sample was found to be contaminated with L. monocytogenes in plant B, while the other three plants were contaminated with L. monocytogenes in different production phases.
Lmo, Listeria monocytogenes; Lin, Listeria innocua; Lwe, Listeria welshimeri; Lgr, Listeria grayi.
Serotyping
Figure 1 shows the lineages of 55 isolates and the sources from which they were recovered. In samples from plants A, C, and D, 19 (34.5%) of 55 isolates were lineage II.2 (1/2b, 3b, and 7), and 65.5% (36/55) were lineage I.1 (1/2a and 3a).
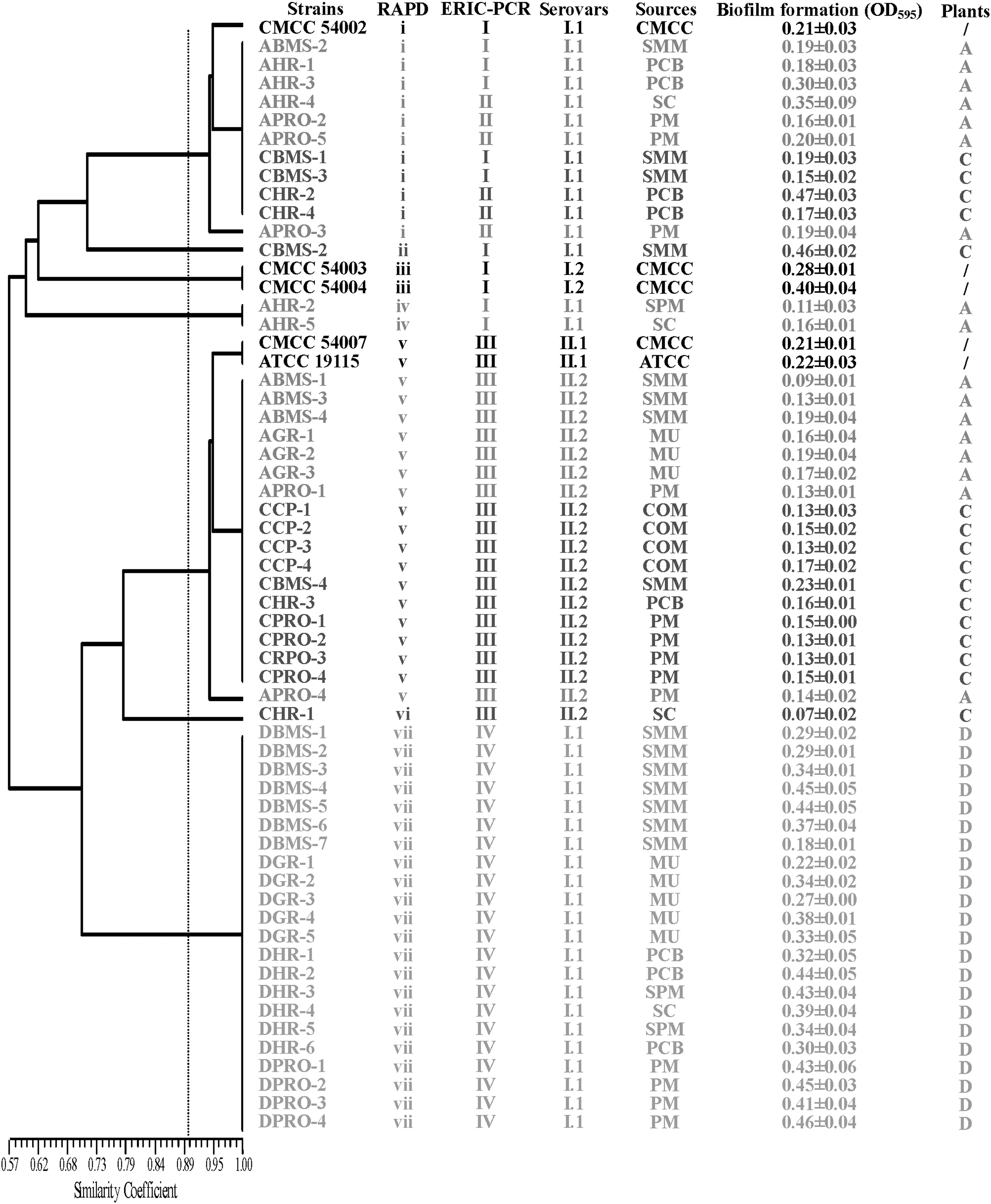
The characterization of Listeria monocytogenes isolates from Flammulina velutipes plant environment samples. RAPD, random amplified polymorphic DNA; ERIC-PCR, Enterobacteria repetitive intergenic consensus–polymerase chain reaction; OD, optical density; CMCC, China Medical Culture Collection; ATCC, American Type Culture Collection; SMM, surfaces of mycelium-scraping machinery; PM, packaged mushrooms; PCB, products conveyor belt; SC, scales; COM, compost; MU, mushrooms; SPM, surfaces of package machinery.
ERIC-PCR typing
The numbers of L. monocytogenes isolates submitted for genotyping are listed in Supplementary Table S1. ERIC-PCR fingerprints showed 5–10 bands ranging from approximately 190 bp to 2300 bp in size. At the coefficient of 70%, 55 isolates fell into three clusters according to the different lineages. At a relative similarity coefficient of 90%, 55 L. monocytogenes isolates were grouped into four clusters designated as I, II, III, and IV (Fig. 2).
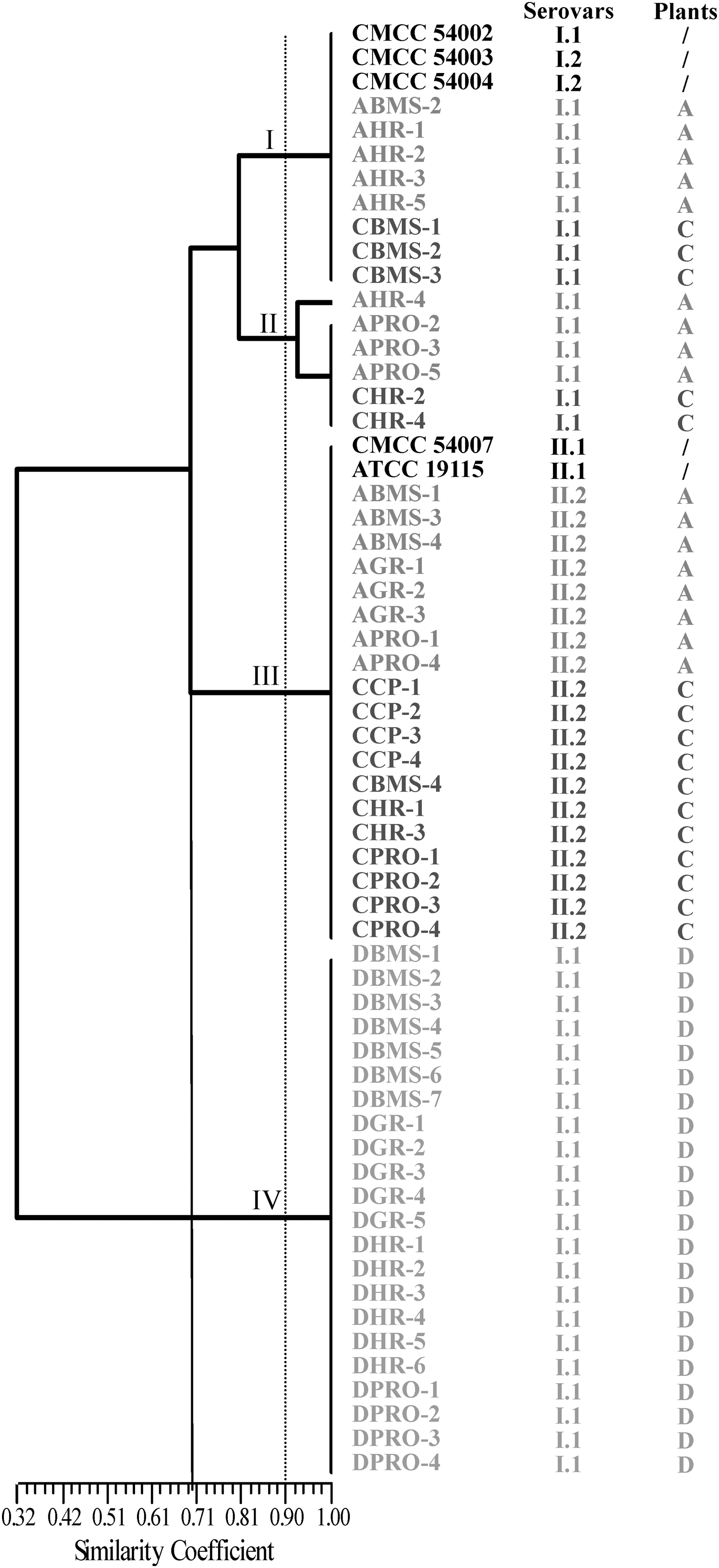
Dendrogram of Listeria monocytogenes isolates recovered from Flammulina velutipes plant samples based on Enterobacteria repetitive intergenic consensus (ERIC)-polymerase chain reaction. L. monocytogenes isolates were named according to the source of isolates; details are shown in Supplementary Table S1. CMCC, China Medical Culture Collection; American Type Culture Collection.
As shown in Figure 2, cluster III predominated in the samples collected from plant A, where 47% of the isolates were recovered from the surfaces of mycelium-scraping machinery, mushrooms, and packaged mushrooms. The remaining nine isolates were divided into two clusters: cluster I and cluster II. Cluster I included one isolate recovered from the surfaces of mycelium-scraping machinery and four isolates recovered from harvesting room including scales, conveyor belts, and surfaces of package machinery. Cluster II comprised one isolate recovered from conveyor belts and three isolates from packaged mushrooms.
In plant C, the 16 isolates were distributed among three clusters. Eleven isolates from compost, surfaces of mycelium-scraping machinery, and packaged mushrooms belonged to cluster III. The three isolates recovered from the surfaces of mycelium-scraping machinery and two isolates recovered from harvesting room belonged to cluster I and cluster II, respectively.
For the samples from plant D, 22 isolates recovered from the surfaces of mycelium-scraping machinery, mushrooms, scales, conveyor belts, and packaged mushrooms exhibited the same ERIC-PCR fingerprints and lineage. At a relative coefficient of 90%, 22 isolates were grouped into cluster IV (Fig. 2).
RAPD typing
A total of 55 isolates collected from plants A, C, and D and 5 reference strains were typed with UBC-155 primer. At a relative coefficient of 90%, 55 isolates were grouped into 5 clusters and 2 singletons (i–vii). The isolates recovered from plant A were mainly grouped into clusters i and v. Five isolates belonging to cluster i were isolated from the surfaces of mycelium-scraping machinery, harvesting room, and packaged mushrooms (Fig. 1). Cluster v included seven isolates recovered from plant A, containing three isolates from the surfaces of mycelium-scraping machinery, three isolates from mushrooms, and one isolate from packaged mushrooms. Isolates recovered from the packaged mushrooms belonged to two lineages and had RAPD patterns identical to those of isolates from the mycelium-scraping machinery and mushrooms, respectively. In plant C, 16 isolates were distributed among the four clusters. Seven isolates recovered from compost, surfaces of mycelium-scraping machinery, mushrooms, and packaged mushrooms belonged to cluster v. Twenty-two isolates recovered from plant D belonged to cluster vii, which was consistent with the result of ERIC-PCR typing.
Comparison between typing methods
At the relative similarity coefficient of 70%, the coincidence rate between lineage and ERIC-PCR was 100%; we also observed a good agreement between lineage and RAPD. At the relative similarity coefficient of 90%, when comparing ERIC-PCR clusters and RAPD clusters, a good agreement was observed between ERIC-PCR and RAPD (i.e., cluster v and vii typing by RAPD belonged to cluster III and IV of ERIC-PCR, respectively).
Biofilm formation
The abilities of 55 L. monocytogenes isolates and 5 reference strains to form biofilm were tested at 25°C in TSB. As shown in Figure 1, the quantity of biofilm obtained after 72 h highly depended on the strain lineage. The OD values of lineage I.1 (serotypes 1/2a and 3a) were significantly higher than that of lineage II.2 (serotypes 1/2b, 3b, and 7) (p<0.01). Surprisingly, several strains belonging to the same cluster as grouped by ERIC-PCR and RAPD had different abilities to form biofilm. Six strains were selected to further confirm the ability of biofilm formation by SEM. Four strains, namely, ATCC 19115, AHR-4, DBMS-3, and DGR-1, formed a three-dimension, organized, dense structure (Figs. 3G and J–L) with individual L. monocytogenes cells surrounded by an exopolymeric substance (Figs. 3A and D–F). Strains ABMS-1 and AGR-2 developed limited biofilms, and a few attached cells were scattered on the glass surfaces (Figs. 3B, H, C, and I).

Micrographs of Listeria monocytogenes biofilms grown in trypticase soy broth at 25°C for 72 h on glass discs by scanning electron microscopy.
Discussion
Recent studies have explored the transmission route of Listeria spp. in the small-scale Agaricus bisporus plant in the United States (Viswanath et al., 2013; Weil et al., 2013). In addition, our laboratory found a high occurrence of L. monocytogenes in retail-level fresh F. velutipes products in China (unpublished data). These data may indicate that conventional disinfection procedures in the production process did not effectively eliminate Listeria spp., and this phenomenon may cause cross-contamination in mushroom plants. In the present study, we specifically focused on the contamination patterns of L. monocytogenes in four mushroom plants and associated environments. Among 295 collected samples, 18.6% were positive for L. monocytogenes. Only plant B was not contaminated with L. monocytogenes. These results indicated that the contamination appeared to originate from the mycelium-scraping machinery and was found on upstream processing equipment and the packaged mushrooms. These data may indicate that cross-contamination of L. monocytogenes occurred during the production process of mushrooms. About 33.9% of the collected samples were positive for nonpathogenic Listeria spp., including L. innocua, L. welshimeri, and L. grayi. No L. ivanovii and L. seeligeri isolates were found in these four plants. L. innocua was predominant over L. monocytogenes in the plants and associated samples, which was consistent with a previous finding (Aguado et al., 2004). Other studies have reported that the presence of L. innocua may reduce the detectability of L. monocytogenes by both overgrowth and the production of inhibitory compounds (Cornu et al., 2002; Zitz et al., 2011). Therefore, the presence of L. innocua may be considered as an indicator of contamination with L. monocytogenes (Greenwood et al., 2005; Pagadala et al., 2012), indicating that a strict sanitation measure should be adopted during the production process.
Serotyping is a useful tool for the epidemiological investigation and tracing of contamination sources. L. monocytogenes strains were divided into four lineages (Roberts, et al., 2006; Ward et al., 2008; Orsi et al., 2011), while Doumith et al. (2004b) grouped L. monocytogenes into three lineages and further defined them into five distinct phylogenetic groups by rapid multiplex PCR, each correlated with the following lineages: I.1 (1/2a-3a), I.2 (1/2c-3c), II.1 (4b-4d-4e), II.2 (1/2b-3b-7), and III (4a-4c). Given that serotypes 3a, 3b, 4d, and 4e are relatively rare in foodborne L. monocytogenes, lineages I.1, II.1, and II.2 were considered as serotypes 1/2a, 4b, and 1/2b, respectively (Doumith et al., 2004a). Previous study reported that 95% of isolates recovered from foods and clinical samples are serotypes 1/2a, 1/2b, 1/2c, and 4b (Pontello et al., 2012). In this study, the specific lineage I (1/2a-3a) and II (1/2b-3b-7) of L. monocytogenes dominated in mushrooms plants, which was consistent with the previous findings on contamination patterns in different food-processing environments while only few 4b serotype has been isolated from facilities (Kathariou, 2002), including those for ready-to-eat foods, cull cow and bull processing plants (Gilbreth et al., 2005; Guerini et al., 2007). Conversely, serotype 4a was found only in a small-scale mushroom-production facility (Viswanath et al., 2013).
L. monocytogenes biofilms can tolerate high concentrations of disinfectants and sanitizers, thereby hampering the decontamination of surfaces (Carpentier and Cerf, 2011; da Silva and De Martinis, 2013). Hence, we were not surprised that the strains belonging to lineage I.1 produced significantly thicker three-dimensional biofilms than those belonging to lineage II.2. This observation was consistent with previous reports that L. monocytogenes strains have different abilities to form biofilms (Borucki et al., 2003; Harvey et al., 2007; Takahashi et al., 2009; Kadam et al., 2013). In addition, L. monocytogenes strains belonging to the same serotype differ in their ability to form biofilms depending on environmental factors (Takahashi et al., 2009). Further studies are thus necessary to elucidate the characteristics of specific serotypes of L. monocytogenes that dominate production environments and are resistant to cleaning and sanitation measures.
Packaged fresh mushrooms stored at low temperatures also allow the growth and survival of L. monocytogenes (Gonzalez-Fandos et al., 2001; Folsom et al., 2006). Thus, L. monocytogenes contamination in final products must be controlled by tracing the pathogen's sources and routes of transmission. ERIC-PCR and RAPD are widely used to trace contamination sources in various food-processing plants (Fonnesbech Vogel et al., 2001; Vogel et al., 2001; Chen et al., 2010; Keeratipibul and Techaruwichit, 2012). In the present study, seven isolates recovered from plant A were grouped into cluster III by ERIC-PCR subtyping at a relative similarity coefficient of 90%. These isolates were recovered from the surfaces of mycelium-scraping machinery, mushrooms, and packaged mushrooms. Samples collected from the surfaces of mycelium-scraping machinery were also positive for L. monocytogenes in plants C and D (Fig. 1). Surprisingly, only a unique fingerprint was found for the isolates recovered from plant D using both ERIC-PCR and RAPD. These results strongly suggested that the surfaces of mycelium-scraping machinery may serve as the primary contamination sources of L. monocytogenes in F. velutipes plants. Among the four plants, L. monocytogenes isolates were recovered from compost in plant C, suggesting that raw materials may serve as the origin source of contamination. Similarly, substrate ingredients were the likely primary contamination source in A. bisporus production facility (Viswanath et al., 2013). Long-term comprehensive studies are needed to trace sources of contamination, and molecular subtyping methods are a useful measure for identifying persistent L. monocytogenes strains at critical control points in mushrooms plants.
Conclusions
Lineage I (1/2a-3a) and II (1/2b-3b-7) were predominant in F. velutipes plants. Genotype and phenotype analyses revealed that L. monocytogenes may spread across the entire production process of mushrooms plant, from compost to the packaged mushrooms. The surfaces of mycelium-scraping machinery may serve as the primary contamination source of L. monocytogenes. Strict rules and sanitation measures for controlling critical contamination points are necessary to avoid jeopardizing the safety of final products.
Footnotes
Acknowledgments
We thank the staff of the microscopy laboratory at the Guangdong Institute of Microbiology for assisting in our scanning electron microscopy studies. We gratefully acknowledge the financial support of the National Natural Science Foundation of China (No. U1031003), the Science and Technology Cooperation Projects of China (No. 2013DFH30070), and Key Projects in the National Science & Technology Pillar Program during the Twelfth Five-year Plan Period (2013BAD16B05).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.