
Abstract
Chlorogenic acid (CA) has been reported to inhibit several pathogens, but the influence of subinhibitory concentrations of CA on virulence expression of pathogens has not been fully elucidated. The aim of this study was to explore the effect of CA on the virulence factor production of Staphylococcus aureus. The minimum inhibitory concentration (MIC) of CA against S. aureus was determined using a broth microdilution method. Hemolysin assays, coagulase titer assays, adherence to solid-phase fibrinogen assays, Western blot, and real-time reverse transcriptase-polymerase chain reaction were performed to evaluate the effect of subinhibitory concentrations of CA on the virulence factors of S. aureus. MIC of CA against S. aureus ATCC29213 was found to be 2.56 mg/mL. At subinhibitory concentrations, CA significantly inhibited the hemolysis and dose-dependently decreased coagulase titer. Reduced binding to fibrinogen and decreased production of SEA were observed with treatment of CA at concentrations ranging from 1/16MIC to 1/2MIC. CA markedly inhibited the expression of hla, sea, and agr genes in S. aureus. These data demonstrate that the virulence expression of S. aureus could be reduced by CA and suggest that CA could be potentially developed as a supplemental strategy to control S. aureus infection and to prevent staphylococcal food poisoning.
Introduction
S
Usually, antibiotics, such as ampicillin, penicillin, and erythromycin, were used to treat severe or systematic infections caused by S. aureus. However, the extensive use of antibiotics in animals and humans has prompted the emergence of multidrug-resistant S. aureus, particularly methicillin-resistant S. aureus, leaving a limited choice for their control. In recent years, there has been a trend of developing new substances to prevent and treat infections caused by these drug-resistant S. aureus strains. Because of their potent antimicrobial properties against a broad spectrum of microorganisms such as S. aureus, Salmonella, and Listeria monocytogenes, plant extracts, especially phytochemicals they contain, have gained great interest (Cowan, 1999; Carson et al., 2006). Many studies have been reported that natural extracts such as oil, licochalcone A, and eugenol have an antimicrobial function (Burt, 2004; Qiu et al., 2010a, 2010b).
Chlorogenic acid (CA) is a family of esters formed between certain trans-cinnamic acids and quinic acid and displays many biological properties such as antiphlogistic, antimutagenic, and antioxidant properties (Clifford, 2000; Lou et al., 2011). CA exists in many kinds of plant foods such as apples, pears, carrot, tomato, potato, and coffee (Upadhyay and Rao, 2013). CA has an obvious inhibitory effect on growth of Trichosporon beigelii, Malassezia furfur, Escherichia coli, Shigella dysenteriae, and Salmonella Typhimurium (Sung and Lee, 2010; Lou et al., 2011). In our previous study, CA was demonstrated to inhibit the growth of several S. aureus strains (Li et al., 2013). However, the influence of CA on virulence expression of pathogens has not been well documented. The objective of this study was to assess the influence of CA on S. aureus virulence factor production via assays for hemolysis, coagulase titer, adherence to solid-phase fibrinogen, and enterotoxin expression.
Materials and Methods
Reagents
Chlorogenic acid (≥95% [titration], CAS: 327-97-9) was acquired from the Sigma Chemical Co. (St. Louis, MO). The solutions were prepared in Luria Bertani (LB) broth (Beijing Land Bridge Technology Co., Ltd., Beijing, China) and stored at −20°C. All chemicals were of analytical grade.
Bacterial strains and culture conditions
S. aureus ATCC 29213 was obtained from the American Type Culture Collection (ATCC). The strain was maintained at −80°C in tryptic soy broth (Beijing Land Bridge Technology Co., Ltd.) containing 25% (vol/vol) glycerol. For hemolysis, coagulase titer, adherence, and Western blot assays, bacteria were incubated with and without CA in LB broth for 24 h (the postexponential phase). The supernatants were harvested by centrifugation (13,400×g, 2 min) and filtered using a 0.22-μm acetate syringe filter.
Determination of minimum inhibitory concentration (MIC)
The MIC of CA against S. aureus ATCC 29213 was determined using a broth microdilution method described by CLSI (2005), with minor modifications. S. aureus ATCC 29213 was cultured in LB broth to an optical density at 600 nm (OD600) of 0.2, and 100 μL of the culture was aliquoted into each well of 96-well plates. CA was added to wells to obtain the final concentrations of 2.56 mg/mL, 1.28 mg/mL, 0.64 mg/mL, 0.32 mg/mL, and 0.16 mg/mL. Following addition of CA, the plate was incubated at 37°C for 24 h. MIC was defined as the lowest concentration at which no visible growth was observed.
Growth curves
The method described by Qiu et al. (2011c) was used with some exceptions. Briefly, bacteria were cultured at 37°C to an OD600 of 0.2 in LB broth, and 100 mL of the cultures was put into 250-mL sterile flasks, followed by the addition of CA at concentrations of 1/16 MIC, 1/8 MIC, 1/4 MIC, 1/2 MIC, and MIC. LB broth without CA was the control. Then bacteria were incubated at 37°C under aerobic conditions. Cell growths were measured by measuring the OD600 values at 2-h intervals.
Hemolysis assay
Hemolysis assay was performed as previously described by Qiu et al. (2012) with modifications. Briefly, 25 μL of defibrinated rabbit blood was mixed with 100 μL volume of culture supernatant in 875 μL of phosphate-buffered saline (PBS) buffer (pH 7.0). After 30 min at 37°C, the unlysed blood cells were removed by centrifugation (5500×g, 20°C, 1 min). The hemolytic activity was monitored by measuring the OD value under 543 nm. The CA-free culture served as the 100% hemolysis control, and the percent hemolysis was calculated by comparison with the CA-free culture.
Coagulase titer assay
Coagulase titers were determined based on the method of Kuroda et al. (2007), with some modification. Briefly, bacteria were grown overnight in LB broth, and the supernatants were filtered with a 0.22-μm membrane. The supernatants were twofold serially diluted with LB broth and then mixed with equal volumes (500 μL) of rabbit plasma (contained 400 μL PBS). After incubation at 37°C for 2 h, coagulase titer was determined as the maximum dilution factor of culture supernatant for coagulation.
Adherence to solid-phase fibrinogen assay
The method described by Barta et al. (1998) was followed with minor exceptions. The 96-well plates were coated with serial dilutions of fibrinogen (20 mg/L), and 50 μL of dilutions was added to each well. After incubation at 4°C for 12 h, the 96-well plates were washed twice, and then 2.5 μg of bovine serum albumin (BSA) containing 50 μL of PBS per mL was added. After incubation at 37°C for 1 h, the plates were washed three times with PBS, and 100 μL of treated cells (OD600=1) was applied to each well. Plates were maintained at room temperature for 2 h. The wells were washed four times with PBS. The bound cells were fixed with 25% formaldehyde and stained with crystal violet. The microtiter plate was read at 560 nm by an enzyme-linked immunosorbent assay reader. The background values from BSA-coated wells were subtracted from sample values.
Western blot assay
According to the method described by Qiu et al. (2010a), the supernatants of culture described earlier were used for Western blot analysis. Following boiling in loading buffer for 5 min, 20 μL of the culture supernatant was subjected to a 10% sodium dodecyl sulfate–polyacrylamide gel. Proteins were transferred to polyvinylidene fluoride membranes by a semidry transfer cell (Bio-Rad, Munich, Germany) at 10 V for 25 min. Membranes were incubated for 2 h at 25°C in 5% nonfat milk powder as a blocking reagent. Antibody to SEA was diluted to 1:500 (Sigma-Aldrich), and a horseradish peroxidase–conjugated anti-rabbit antiserum (Sigma-Aldrich) diluted to 1:1000 was used as the secondary antibody. The blots were developed using Amersham ECL Western blotting detection reagents (GE Healthcare, Buckinghamshire, UK).
RNA isolation and real-time reverse transcriptase RT-polymerase chain reaction (RT-PCR)
S. aureus ATCC 29213 was incubated with and without CA in LB broth for 12 h, then the total RNAs from the bacterial cultures was extracted as described by Qiu et al. (2011b) using an RNApure Bacteria Kit (Tiangen, Beijing, China). RNA was reverse transcribed into cDNA using the Takara PrimeScript™ RT reagent kit (Perfect Real Time) (Takara, Kyoto, Japan) according to the manufacturer's instructions. The primer pairs from Qiu et al. (2010c) and Qiu et al. (2011c) were used in real-time RT-PCR tests. The reagents consisted of 12.5 μL SYBR Premix Ex Taq (Takara), 1 μL of each primer (10 μmol/L) and 2 μL of sample cDNA in a final volume of 25 μL. The reactions were performed using the IQ5 software (Bio-Rad, Hercules, CA). Cycling conditions consisted of an initial denaturation step at 95°C for 30 s, followed by 40 cycles at 95°C for 5 s, 55°C for 30 s, and 72°C for 30 s. The melting curves for the PCR products were obtained by stepwise increases in temperature from 60°C to 95°C. All samples were analyzed in triplicate, and the gyrase (gyr) gene served as an internal control to normalize the expressional levels between samples. The relative expression levels were analyzed by the 2-ΔΔCT method.
Statistical analysis
All experiments were performed in triplicate. Statistical analysis was performed with the SPSS 19.0 statistical software (IBM, Armonk, NY). Statistical differences were examined using the independent Student t-test, and p<0.05 was considered to be statistically significant.
Results
Influence of chlorogenic acid on S. aureus growth
We found that the MIC of CA against S. aureus ATCC 29213 was 2.56 mg/mL. The results demonstrated that at the concentrations of 1/16MIC, 1/8MIC, and 1/4MIC, CA had no obvious effects on the growth of S. aureus (Fig. 1). However, at the concentration of MIC or 1/2MIC, the growth of S. aureus was significantly inhibited. The OD600 value of the culture treated with CA at the concentration of 1/2MIC was 19.60% of the control after 24 h.
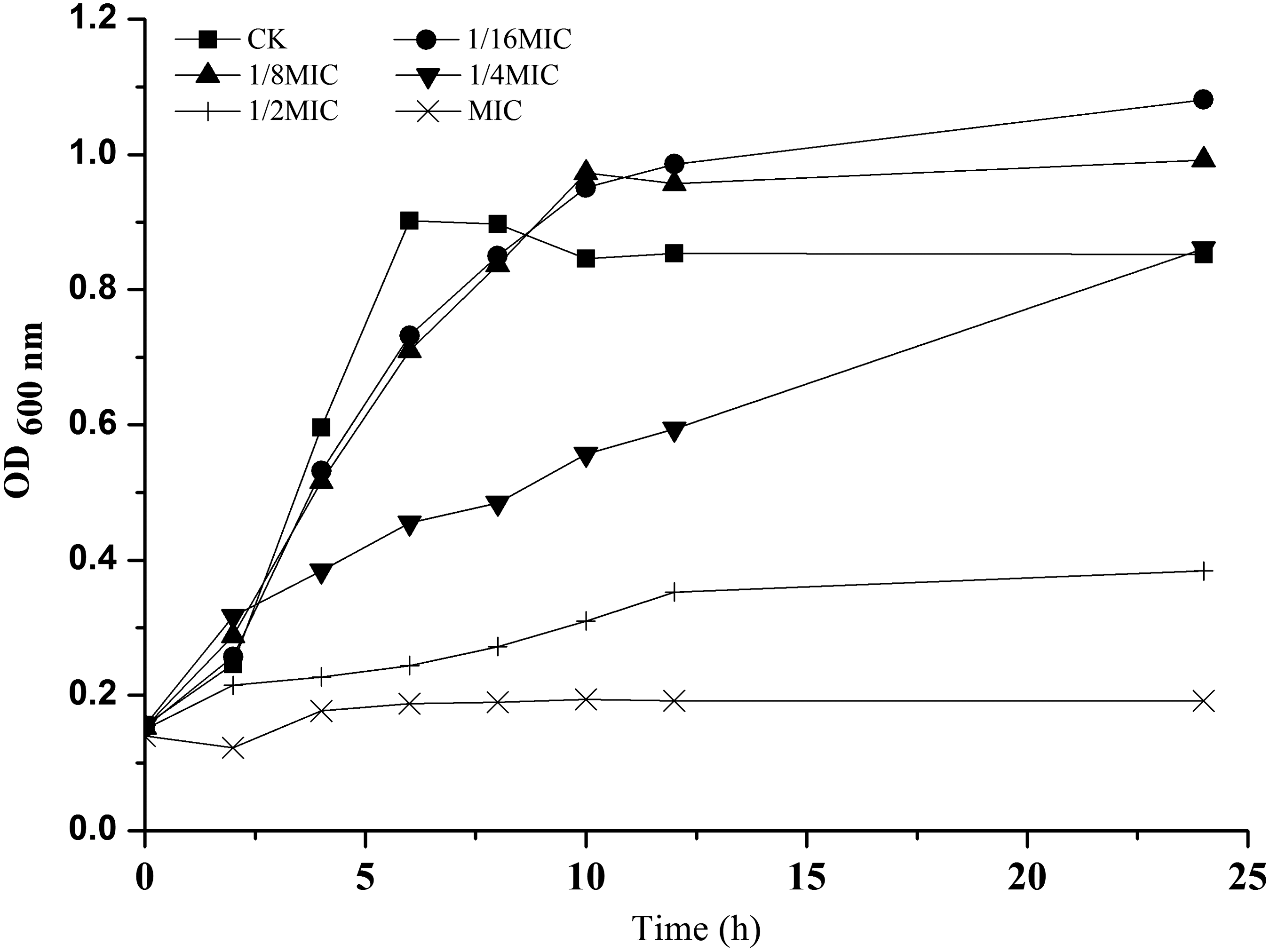
Growth curve for Staphylococcus aureus strain ATCC 29213 in the presence of different concentrations of chlorogenic acid. These curves represent the average values of three reproducible experiments. CK, control; MIC, minimum inhibitory concentration; OD, optical density.
Chlorogenic acid inhibited hemolytic activity of S. aureus culture supernatants
CA led to a significant inhibition of S. aureus hemolysis (Fig. 2). The hemolysis values of culture treated with CA at 1/16MIC, 1/8MIC, 1/4MIC, and 1/2MIC were 48.18%, 19.72%, 6.14%, and 3.89% of the control, respectively. And a dose-dependent (from 1/16 MIC to 1/2MIC) attenuation of hemolysis was observed in S. aureus ATCC 29213.
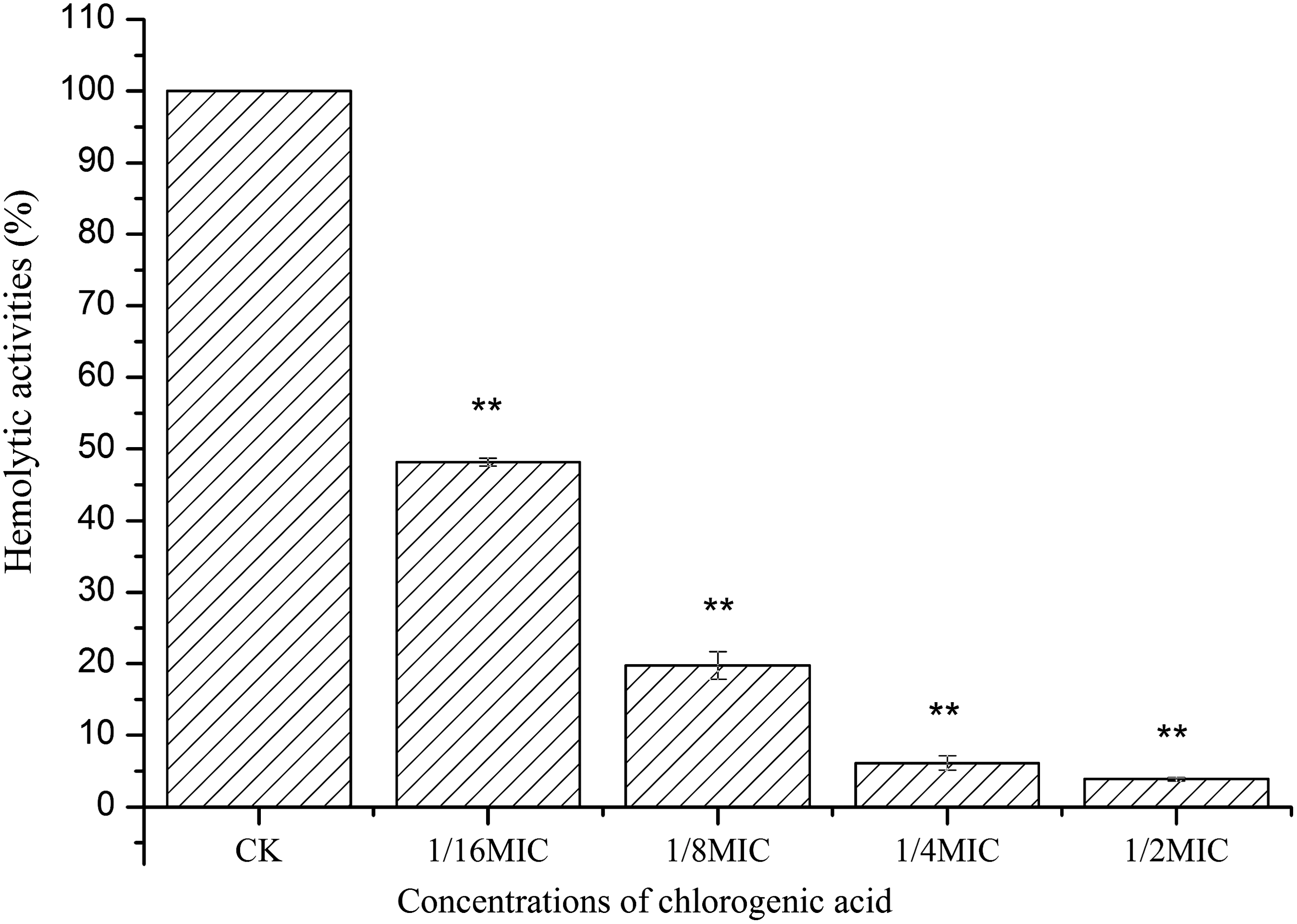
Hemolytic activities of the supernatant of Staphylococcus aureus grown at different concentrations of chlorogenic acid. Bars represent the standard deviation (n=6). Student t-test was used to compare each culture with chlorogenic acid to the untreated culture (**p<0.01). CK, control; MIC, minimum inhibitory concentration.
Chlorogenic acid reduced secretion of coagulation by S. aureus
Coagulase titer, mainly represented by the extracellular proteins, decreased with the addition of CA at the concentration ranging from 1/16 MIC to 1/2 MIC (Table 1). No coagula was obtained at 1/2 MIC.
The titer is the highest dilution of sample causing coagulation.
MIC, minimum inhibitory concentration.
CA decreased S. aureus adherence to solid-phase fibrinogen
S. aureus adherence to solid-phase fibrinogen is usually promoted by a clumping factor and coagulase. With the increasing of CA concentrations from 1/16MIC to 1/2MIC, reduced binding of S. aureus (from 76.94% to 21.71% of the control) was observed (Fig. 3). Percentage binding of cells grown with CA at 1/2MIC and 1/4MIC was significantly different from the control (p<0.01).
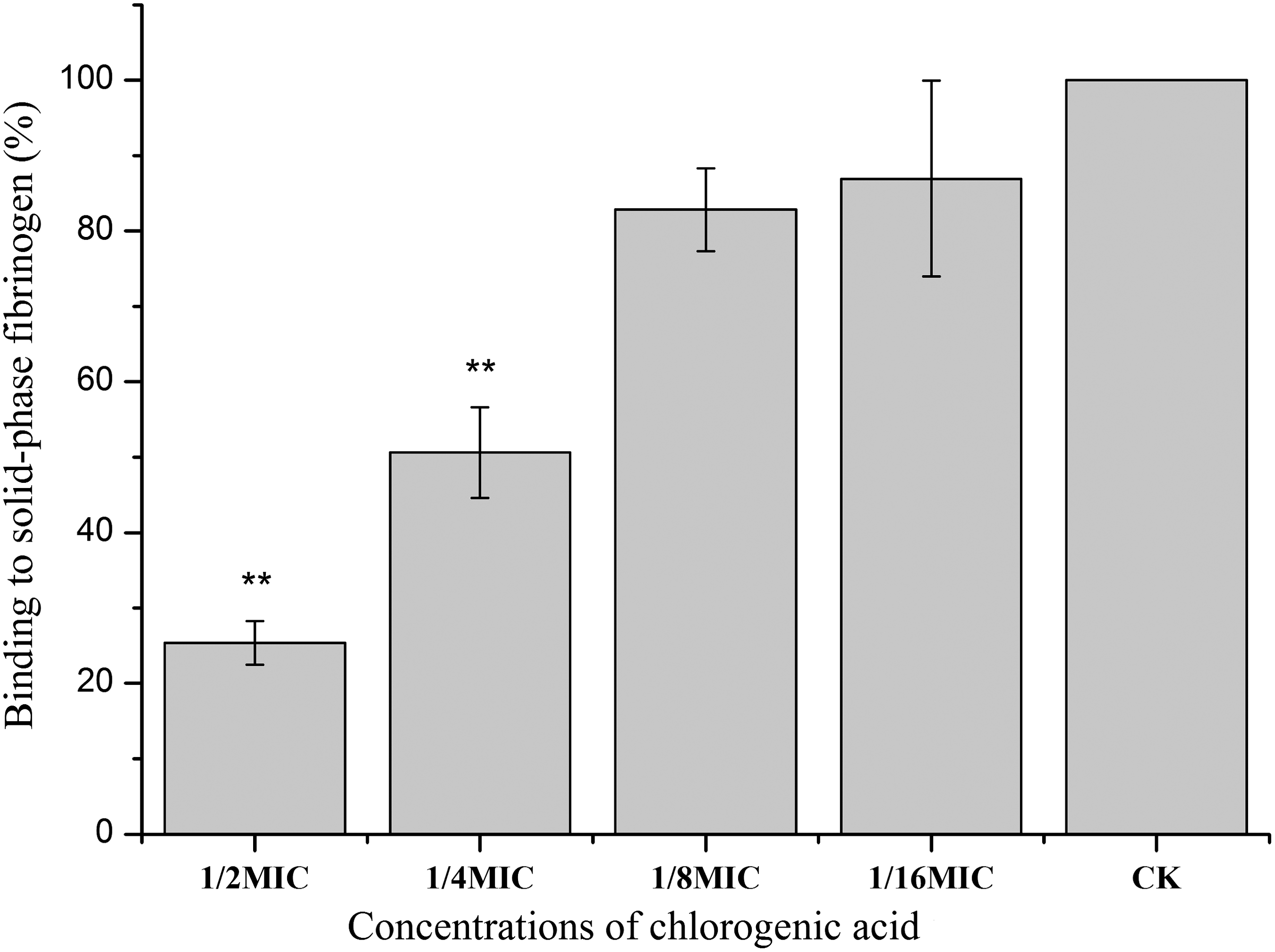
Binding of Staphylococcus aureus to solid-phase fibrinogen with different concentrations of chlorogenic acid. Bars represent the standard deviation (n=6). Student t-test was used to compare each culture with chlorogenic acid to the untreated culture (*p<0.05; **p<0.01). CK, ; MIC, minimum inhibitory concentration.
CA reduced the SEA secretion by S. aureus
The culture supernatants were used for Western blot to detect the release of SEA. As shown in Figure 4, CA caused a dose-dependent reduction in SEA. Growth with 1/16MIC of CA led to a remarkable decrease in SEA secretion, whereas at 1/2MIC, no SEA could be detected in the S. aureus ATCC 29213.

Western-blot analysis of SEA production by strain ATCC 29213 after growth with different concentrations of chlorogenic acid. MIC, minimum inhibitory concentration.
CA represses sea, hla, and agrA transcription
To determine whether the reduced production of virulence factors in the culture supernatants was due to a decrease in hla and sea transcription, real-time RT-PCR was performed to determine the relative expression levels of the investigated genes in S. aureus after treatment with CA. In addition, because the expression of these genes is positively regulated by the agr locus (Cheung and Zhang, 2002), the transcription level of agrA was also detected. The results demonstrated that the expression levels of these genes in S. aureus were markedly repressed by CA (Fig. 5). When cultured with 1/8MIC of CA, the expression levels of sea, hla, and agrA were decreased by 2.1-, 5.9- and 1.6-fold of the control, respectively.
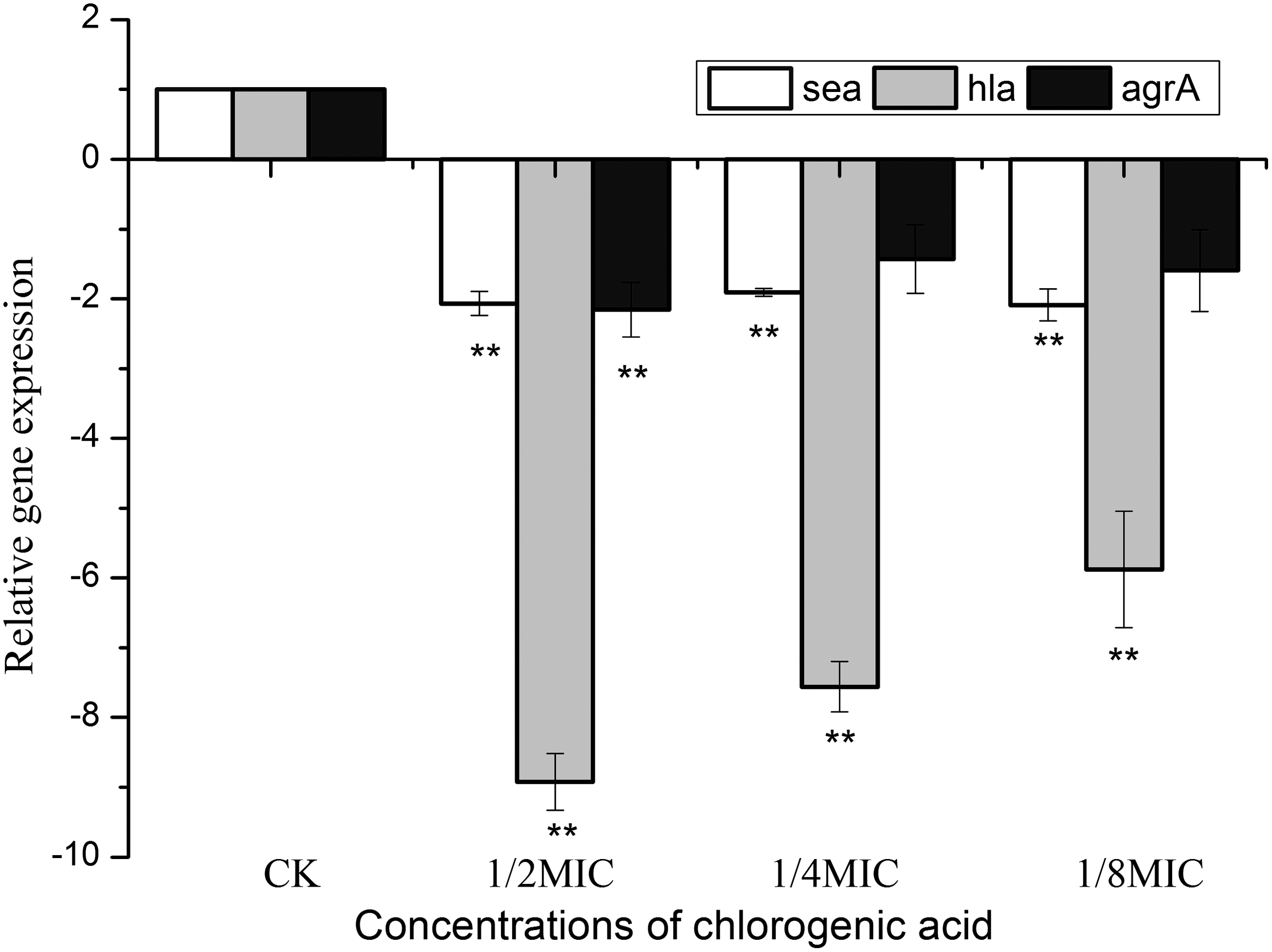
Effect of chlorogenic acid on the transcription of sea, hla, and agrA in Staphylococcus aureus. Data are expressed as the means±standard deviation for three independent experiments. Student t-test was used to determine statistical differences. (**p<0.01). CK, control; MIC, minimum inhibitory concentration.
Discussion
Studies reported CA has the ability to inhibit both Gram-negative and Gram-positive bacteria growth (Lou et al., 2011), but the influence of subinhibitory concentrations of CA on virulence expression of the pathogens remained obscure. In this study, we demonstrated that CA could attenuate virulence-associated factors of enterotoxigenic S. aureus strains for the first time.
Antivirulence therapy in the treatment S. aureus infections has gained an increasing interest, which offers opportunities to inhibit pathogenesis without placing immediate life-or-death pressure on the target microorganism. S. aureus produces various exotoxins that contribute to its pathogenicity (Kokan and Bergdoll, 1987; Dinges et al., 2000; Qiu et al., 2010a). Therefore, the clinical performance of antibiotics is evaluated as follows: one is their bactericidal or bacteriostatic activity, and the other is their influence on the release of virulence factors. Several antibiotics such as linezolid and clindamycin affect the secretion of exotoxins by S. aureus, especially at subconcentrations (Stevens et al., 2007). These antibiotics are known as protein-synthesis-inhibitor because they significantly inhibit the production of several virulence factors in S. aureus at subinhibitory concentrations. However, antibiotics affect the cell wall, inducing the expression of α-hemolysin, enterotoxins, and toxic shock syndrome toxin-1 (TSST-1) through a stimulatory effect on other exoprotein synthesis (Herbert et al., 2001). Several studies have reported that plant extracts, different from mechanisms used by antibiotics, can reduce production of exotoxins at subinhibitory concentrations (Xiang et al., 2010; Leng et al., 2011; Qiu et al., 2012). Qiu et al. (2011b) observed that costus oil decreased the production of a-toxin, TSST-1, and enterotoxins A and B in S. aureus. Qiu et al. (2010a) reported that subinhibitory concentrations of thymol reduced enterotoxins A and B and α-hemolysin production in S. aureus. Li et al. (2011) demonstrated that the production of S. aureus exotoxins was decreased by subinhibitory concentrations of peppermint oil in a dose-dependent manner. Those studies all indicate that natural compounds could be utilized for the treatment of S. aureus infections through interfering with virulence factor expression.
S. aureus produces a wide variety of toxins, in which enterotoxins are known as a leading cause of food poisoning (Pereira et al., 2009). It is shown that SEA, associated with other staphylococcal enterotoxins, is most frequently involved in food poisoning outbreaks worldwide, followed by SEB, SEC or SED, SEE and other identified new SEs (Argudin et al., 2010). Currently, chemical preservatives are commonly used in foods to control microbial contamination and extend shelf life of food. However, with the demanding for the safety, natural, nutritional and functional of food additives, people pay more and more attention on natural preservatives. With the ability of CA to decrease SEA production, together with its antiphlogistic, antimutagenic, and antioxidant properties (Clifford, 2000; Lou et al., 2011), CA has the potential to be developed as a food additive or preventive agent for food poisoning.
In S. aureus, global regulators such as agr and SarA control the secretion of virulence factors during growth (Cheung and Zhang, 2002). Previous studies have reported that subinhibitory concentrations of natural products can affect the translation of certain regulatory gene products in S. aureus, which, in turn, alter the transcription of toxin-encoding genes. Those products included allicin (Leng et al., 2011), eugenol (Qiu et al., 2010a), luteolin (Qiu et al., 2011a), and essential oils (Smith-Palmer et al., 2004). Qiu et al. (2010b) found licochalcone A markedly decreased the transcription of sea, seb, and agrA in S. aureus ATCC 29213. Lee et al. (2012) reported that flavone clearly repressed the transcription of hla but elevated agrA transcription in S. aureus. To illuminate whether the SEA and HLA expressions were related to the agr locus, an RT-PCR assay was performed to assess the impact of CA on the expression level of agr in S. aureus. The results indicated that the expression level of agrA was significantly decreased when S. aureus was cultured with 1/8MIC of CA. Therefore, we presume that the reduced secretion of these toxins may partly depend on the downregulation of the agr gene induced by CA.
In summary, we found that subinhibitory concentrations of CA significantly affected virulence factors production in S. aureus. CA could be promisingly used as a supplemental agent to control S. aureus infection and to prevent staphylococcal food poisoning through inhibiting the growth of S. aureus as well as reducing the production of exotoxins. Further research is required to evaluate the toxicity of CA prior to its real application.
Footnotes
Acknowledgments
This study was supported in part by China Postdoctoral Science Foundation (No. 2011M501483), National Natural Science Foundation of China (No. 31271858), Twelve-Five Science and Technology Support Program (2012BAH30F03), New Century Excellent Talent Support Plan (NCET-13-0488), and the International Science and Technology Collaboration Research Fund (No. A213021202) in Northwest A&F University.
Disclosure Statement
No competing financial interests exist.