
Abstract
Several methods for the rapid and specific detection of Salmonella spp. in meat have been described. This study was conducted to evaluate the capability of the VIDAS® UP (SPT [Salmonella Phage Technology]), an enzyme-linked fluorescent immunoassay method, and fluorescence in situ hybridization (FISH) to complement the International Organization for Standardization Method 6579 (ISO) in detecting Salmonella spp. from beef, pork, and poultry meat samples. The meat was inoculated with a mixture of Salmonella spp. on three levels of contamination. It was also checked that the tests did not produce cross-reactions with other Enterobacteriaceae rods. On the basis of the results, the relative specificity, relative accordance, and relative sensitivity of the method were determined. In meat samples, Vidas UP and FISH detection results were in substantial agreement with ISO, with relative specificity, accordance, and sensitivity rates of 90%, 96.3%, and 100%, respectively, for Vidas UP and 100%, 100%, and 99.4%, respectively, for FISH. This is the first report on the evaluation of both Vidas UP and FISH compared to ISO for the rapid detection of Salmonella enterica serovars in meat.
Introduction
S
Fluorescence in situ hybridization (FISH) detects nucleic acid sequences by a fluorescently labeled 16S rRNA oligonucleotide probe, which hybridizes specifically to its complementary rRNA target sequence within the intact cell. Probes are 15–25 nucleotides in length labeled covalently at the 5′ end with a fluorescent dye. They bind specifically to rRNA in ribosomes of the target cells, thus identifying them. Such a solution significantly increases the determination sensitivity; since rRNA is an integral part of bacterial ribosome, it is found in the cell in a large number of copies. The procedure includes a few steps: fixation of the specimen, preparation of the sample, hybridization with the respective probes for detecting the respective target sequences, washing steps to remove unbound probes, mounting, and visualization (Moter and Göbel, 2000). The combination of examined material in a test tube with a complementary probe has been investigated under an epifluorescence microscope with a flow cytometer or capillary electrophoresis. The FISH technique and its modifications are now widely used in medicine and environmental analyses (Bishop, 2010; Lew et al., 2010; Liao et al., 2010; Lawson et al., 2011; Loonen et al., 2011; Stefanis et al., 2013). Food, due to its specific and diversified physical and chemical properties, is a heterogeneous material for testing with this technique. A selection of optimal preparation conditions is a prerequisite for the effective application of the FISH technique for identification of food pathogens (Cocolin, 2007).
VIDAS® UP (SPT [Salmonella Phage Technology]) tests are another series of assays introduced to the market by Biomerieux in 2011. They are based on the application of recombinant phage proteins, which are specific for Salmonella spp. and labeled with alkaline phosphatase. The VIDAS apparatus is necessary for conducting these assays. The previous tests were based on an antigen–antibody reaction (VIDAS® SLM and VIDAS® XPRES, Biomerieux). Although studies carried out with these tests have demonstrated their high sensitivity, false-positive results may be produced in assays that are overloaded with accompanying microflora which, in turn, generates costs and extends the testing time due to the need for confirmation of the results with the standard method (Chajęcka-Wierzchowska et al., 2012; Temelli et al., 2012).
VIDAS UP (SPT) tests are Association of Official Agricultural Chemists certified and have been validated for 20 different matrixes. However, due to a wide variability of food products, the manufacturer still recommends checking whether the composition of the matrix does not influence the results of the analysis.
The objective of the studies was to investigate the potential for detection of Salmonella in meat with two alternative methods: FISH and the automated enzyme-linked fluorescent immunoassay–based technique VIDAS UP (SPT), and to compare the results with the findings obtained with the reference method International Organization for Standardization (ISO) 6579.
Materials and Methods
Samples
Samples (400 g each) of beef (n=20), pork (n=20), and poultry (chicken breast) meat (n=20) were purchased from a retail market in Olsztyn (northeast Poland). Knowing that microorganisms can be spread due to Poisson distribution in meat samples, samples (400 g) were homogenized (cut into very small pieces and mixed), and homogenized meat was divided into 25-g portions (for FISH, ISO, and VIDAS) and 10 g (for the determination of total viable count [TVC], Enterobacteriaceae [EB]) (Fig. 1). One 25-g sample was secreted and was examined (PN-ISO 6579:2003) for the presence of Salmonella spp. to make sure that Salmonella spp. was absent.
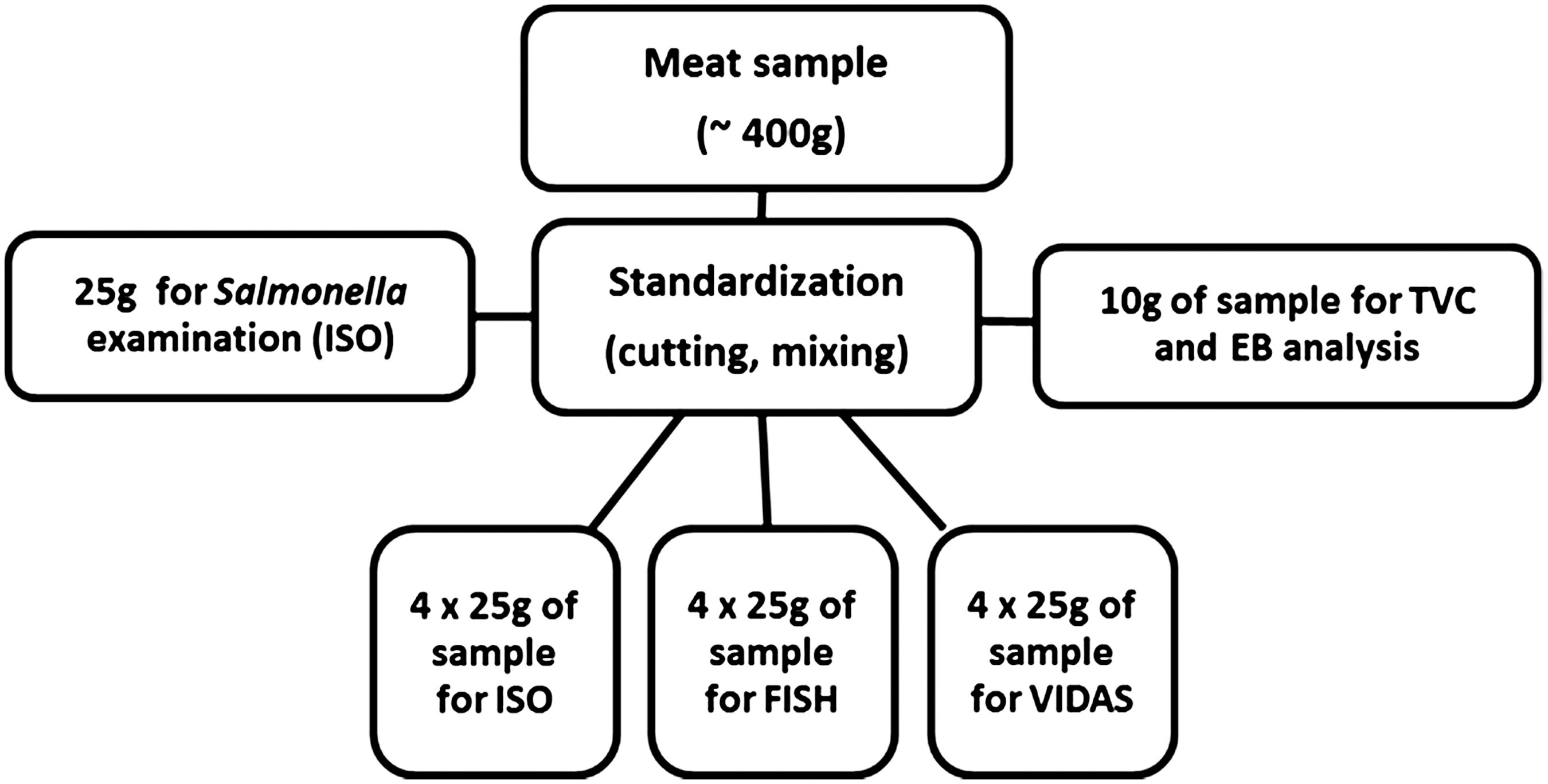
Schematic diagram of samples preparation. ISO, International Organization for Standardization; TVC, total viable count; EB, Enterobacteriaceae; FISH, fluorescence in situ hybridization.
The TEMPO (Biomerieux) automated system was used for enumeration of TVC and EB in meat samples as described previously (Kłębukowska et al., 2013) The goal was to assess the so-called “product matrix.”
Portions of meat (25 g) were artificially contaminated with Salmonella cells on three levels: high (1000–5000 cells per 25 g), medium (100–500 cells per 25 g), and low (10–50 cells per 25 g). For FISH and ISO analysis, samples were supplemented with 225 mL of buffered peptone water (Merck), and for VIDAS analysis they were mixed with buffered peptone water with a Vidas UP supplement. The pre-enrichment phase was carried out at 41.4°C/24 h, and further analyses were then conducted in accordance with PN-ISO-6579:2003 with a mini-Vidas immunoanalyzer and the FISH technique.
Probes and their verification
Three oligonucleotide probes were used: a Salmonella enterica probe, Sal3 (Nordentoft et al., 1997); the universal bacteria probe EUB338; and NON338 (Christensen et al., 1999). Oligonucleotide probes labeled with fluorescent dyes (Cy5, Cy3) at 5′ ends (i.e., EUB338–Cy5, NON338–Cy5, Sal3 –Cy3 [Bionovo], respectively), were used in the studies.
The verification of probe binding was made to optimize the protocol for our research to ensure that the stage of cell preparation for hybridization was properly conducted. The verification was carried out with monocultures and mixed cultures of strains isolated from food at the Department of Industrial and Food Microbiology: Salmonella spp. SLMX1, Salmonella spp. SLM3, and the reference strains: Salmonella enterica subsp. anatum ATCC® 9270, Salmonella Typhimurium ATCC® 14028, Salmonella Enteritidis ATCC® 13076, Escherichia coli ATCC® 11229, Proteus mirabilis ATCC® 25933, P. vulgaris ATCC® 33420, Citrobacter freundii NCTC® 9750, and Enterobacter aerogenes ATCC® 13048. These assays were aimed at testing the correctness of Sal3 probe binding only with the genetic material of Salmonella spp. cells (Nordentoft et al., 1997). In the tests of Salmonella spp. monocultures, total superimposition of the signal of the cells visualized with a specific Cy3 fluorescence-labeled probe and a universal Cy5-labeled EUB338 probe and with a DAPI intercalator (4′,6-diamidyne-2-phenylindole) was assumed as the correct result. The observed superimposition of all signals indicates correct permeabilization of Salmonella spp. cells and the binding of all cells found in the solution with Sal3 probe. In the assays with mixed cultures, total superimposition of the signals of the cells bound to EUB338 and DAPI and fluorescence of the selected cells (Salmonella) labeled with a Sal3 probe were assumed as the correct result. Nonspecific binding of oligonucleotide was verified with a NON338 probe that constituted a negative control. In addition, superimposition of the signals from fluorophores Cy3 and Cy5 with DAPI fluorophore was confirmed.
Analysis with FISH
The volume of 1000 μL of 24-h strain culture of the pre-enriched sample was collected and centrifuged at 12,000 r/10 min. The supernatant was decanted and the sediment was washed twice with 1000 μL of phosphate-buffered saline (PBS) (Sigma). The cells were suspended in 4% paraformaldehyde (Sigma), stored at 4°C for 4 h, and then washed twice in ethanol–PBS solution (1:1). The cells were suspended in 500 μL of ethanol–PBS mixture and 10 μL of the prepared solution was then placed onto 8-well polytetrafluoroethylene (Teflon) glasses (Thermo Scientific) and dried. The glasses were then washed in a Hellendhal-type container for 3 min in 50%, 80%, and 96% ethanol. Ten microliters of hybridization buffer (900 mM NaCl, 20 mM Tris/HCl, 0.01% sodium dodecyl sulfate [SDS], ddH2O) containing 5 ng of Sal3 probe/μL and 5 ng of EUB338 probe/μL was then applied to the dried preparations. Hybridization was performed in humid chambers in a hybridization oven for 3 h at 45°C. After incubation, the glasses were washed in a washing-buffer solution (900 mM NaCl, 20 mM Tris/HCl, 0.01% SDS, ddH2O) at 45°C for 15 min. Vectashield HardSet Mounting Medium oil with DAPI (Vector Labs, Burlingame, CA) was then placed onto the glasses that were kept at 4°C for 15 min. The FISH methodology modified by Vieira-Pinto et al. (2008) was used. The preparations were analyzed with an epifluorescent Olympus BX51 microscope (Olympus Corp.) equipped with a 100-W mercury burner, a color digital XC10 camera (for observations in infrared light) cooled with the Peltier system with a CCD photosensitive element and appropriate filter sets for fluorophores: Cy3 (U-M41007 within the range of 530–560 nm), Cy5 (U-M41008 within the range of 590–650 nm), and DAPI (U-MNU2 within the range of 360–370 nm). The image analysis was performed with a CellSens Dimension (Olympus) plus Solution Multi Channel 5D software. The images of the preparations were taken at a constant exposure time.
Analysis with a miniVIDAS immunoanalyzer
VIDAS UP Salmonella is a combination of a two-step assay using a cocktail of monoclonal antibodies, and recombinant phage proteins were used. They target both somatic and flagellar antigens, allowing detection of both motile and nonmotile strains. The miniVIDAS apparatus reads the results based on the intensity of fluorescence (ELFA [enzyme-linked fluorescence assay]) generated during the reaction of a product. The individual stages of the analysis were performed in accordance with the methodology specified by the manufacturer (Biomerieux) (User's Manual, 2011).
Statistical analysis
On the basis of the results, the relative specificity, accordance, sensitivity, and the level of detection of the method were determined. To define these parameters, all experiments were repeated separately for beef, pork, and poultry meat in 80 repetitions for each of them. After analyzing each sample, the positive and negative accordances were determined as the number of samples with the same positive or negative result. Negative discordance, understood as the number of expected positive samples for negative samples for the alternative method, and positive discordance, understood as the number of expected negative samples for positive samples for the alternative method, were determined. Relative selectivity (SP) was defined as the percent of negative samples showing a proper negative result:
where NA stands for negative accordance and PD stands for positive discordance.
Relative sensitivity (SE) was determined as the percent of positive samples showing a proper positive result:
where PA stands for positive accordance and ND stands for negative discordance.
Relative accordance correctness (AC) was calculated as a degree of accordance of the expected results and the results obtained by the alternative methods:
where PA stands for positive accordance, NA stands for negative accordance, and N stands for total sample number.
The results were also complemented with Cohen's κ test to evaluate the significant similarities between the (1) expected results and ISO results, (2) expected results and VIDAS results, and (3) expected results and FISH results using PQStat Software (version 1.4.6).
Results
The microbiological analysis of meat with a TEMPO apparatus demonstrated a diverse level of contamination in the meat samples. The highest level of contamination was detected in poultry meat (Table 1). The total number of mesophilic aerobic microflora ranged from 3.1 log colony-forming units (CFU)/g to 6.2 log CFU/g, while the number of Enterobacteriaceae ranged between 2.2 log CFU/g and 3.2 log CFU/g. In the beef and pork samples, the total number of mesophilic aerobic microflora did not exceed log 3.6 CFU/g and for Enterobacteriaceae—log 1.8 CFU/g (Table 1). Salmonella spp. were not detected in any of the purchased meat samples.
Results for relative detection level assays: Number of samples negative or positive for each method at each concentration level.
ISO, International Organization for Standardization; FISH, fluorescence in situ hybridization; CFU, colony-forming units; TVC, total mesophilic count; EB, Enterobacteriaceae; NC, noncontaminated meat samples; FP, false-positive results.
The results of the tests for the presence of S. enterica in the samples of contaminated meat depended on the level of introduced inoculum as well as on the number of accompanying microflora. The least-satisfactory results were recorded for the samples of poultry meat, in which the total number of mesophilic aerobic microorganisms and the number of Enterobacteriaceae were the highest. It constituted a substantially wide matrix of the product and, at the same time, a considerable competition to S. enterica inoculated into the sample. Since the VIDAS tests revealed four positive results in the control samples (noncontaminated), the specificity of this method was 80%. False-positive results were also identified in two samples of beef and, in this case, the specificity was 90%. Regardless of the type of investigated meat, no false-negative result was recorded and, thus, the sensitivity of this method was 100%. The level of introduced contamination did not exert any impact on the results of testing with this method. In all samples into which the pathogen was introduced, the apparatus demonstrated a positive result (Table 2).
FISH, fluorescence in situ hybridization; ISO, International Organization for Standardization; SP, relative selectivity; Ac, accordance correctness.
The verification of probe binding indicated that the probe Sal3 hybridized to all used Salmonella strains. No cross-reaction to other strains of the Enterobacteriaceae family was observed. The application of the FISH technique did not yield any positive result in the samples that were not contaminated with S. enterica. The specificity of this method was thus 100%, whereas the relative sensitivity was 98.3%, as Salmonella was not detected in one contaminated sample (Table 2).
Of the 180 samples of meat contaminated with S. enterica, the presence of these bacteria was detected with a VIDAS test in all samples. The application of the FISH technique detected Salmonella in all except for one sample of poultry meat, while the standard method did not demonstrate these bacteria in five samples. The false-negative results determined in accordance with the ISO standard were recorded in one sample of beef contaminated with Salmonella at a low level (10–50 cells per 25 g) and in four samples of poultry meat, of which two false-negative results were detected in meat contaminated at a low level, one false-negative result in the sample contaminated at a medium level (100–500 cells per 25 g), and one false-negative result in one sample contaminated at a high level (1000–5000 cells per 25 g) (Table 1). The FISH technique yielded a false-negative result in the sample of chicken meat that was artificially contaminated with Salmonella spp. at a low level (10–50 cells per 25 g).
Discussion
Identification of Salmonella in food consists of the detection of these bacteria in a specific quantity of product, usually in 25 g (25 mL) (Eijkelkamp et al., 2009). Both in the typical method (ISO-6579:2003) and in its modifications, the first stage is the phase of nonselective multiplication. This stage is very important and aims at regenerating bacterial cells, since food production is associated with technological processing, such as heating, which may cause sublethal cellular damage (Taskila et al., 2012; Rathnayaka, 2011). Omitting the stage of pre-enrichment and culturing the examined material directly onto a solid medium may produce false-negative results.
Of the 180 meat samples contaminated with S. enterica, all samples were detected positively with the VIDAS UP test, while 179 samples and 175 samples were positive with the FISH and the standard method, respectively (Table 2). The FISH technique yielded a false-negative result in one sample. Moller et al. (2013) indicated that it might be possible to limit Salmonella spp. growth with accompanying microflora. Perhaps due to the substantial count of accompanying microflora and its antagonism and competition for nutrients, Salmonella spp. may have been unable to multiply to the number constituting the threshold of detection. The higher number of false-negative results (in five samples) generated with the standard method was probably caused by a high count of accompanying microflora, which yielded abundant growth on the plates with selective media, which significantly hindered interpretation of the results. Some of the Proteus spp. rods grow on XLD medium in colonies that are typical of Salmonella spp. The erroneous selection of the colonies for further confirmation tests may have been the cause of the false-negative results. Some authors have suggested that the multistaged nature of testing and subjective readings based on visual color changes may prompt mistakes in interpreting the results (Tomás et al., 2009). Özer and Kimiran-Erde (2013) and Vieira-Pinto (2005) had a different explanation for the higher number of positive results generated by the FISH technique in comparison with the standard method. They explained the significant differences in the number of positive samples by the potential presence of sublethally damaged cells or VBNC cells. These authors believe that the possibility of obtaining false-negative results with the standard method may be associated with the specific environment (i.e., food) (due to diverse content of salt, preservatives, pH, etc.). Bacterial cells exposed to such stress conditions may not grow on solid media. The quoted authors did not, however, verify the correctness of the protocol and probe binding. Such discrepancies in the results may be related to nonspecific probe binding. In our studies, meat was artificially contaminated with live, active, and cultured Salmonella cells. Therefore, the reason for the higher number of positive cells obtained with the alternative methods, as compared to the reference method, may be explained by the higher specificity of these methods rather than with a capacity to detect the cells in the VBNC state.
Assuming the time of analysis both in FISH and in the immunoenzymatic method, the presence of Salmonella during 20–26 h can be excluded. With the standard method, this stage takes about 70 h. The clear advantage of the VIDAS method is the lack of false-negative results, although false-positive results also extend the time period and generate costs. Some other studies have also indicated the possibility of obtaining false-positive results with a VIDAS immunoanalyzer (Vieira-Pinto et al., 2007; Chajęcka-Wierzchowska et al., 2012; Zadernowska et al., 2014a). This may result from cross-reactions with other Enterobacteriaceae present in the matrix (meat). Numerous authors have suggested a high correlation between the VIDAS technique and other culture methods. De Medici et al. (1998) indicated full agreement between VIDAS immunoconcentration of Salmonella spp. and ISO; McMahon et al. (2004) indicated that VIDAS SLM performed as well as the U.S. Food and Drug Administration (FDA) method; Reiter et al. (2007) found an even higher percentage of Salmonella spp.–positive samples by VIDAS SLM versus the FDA method.
Conclusions
Both the application of the FISH technique and Vidas UP tests allow for a substantial reduction of waiting time for the results, which is particularly important for analyses of raw materials and products with a short shelf-life. Both methods have high sensitivity and specificity. The analysis of food with the FISH technique and with the miniVIDAS apparatus requires a financial input for the necessary equipment. When a laboratory is appropriately equipped, the costs of single assays with the standard method (ISO), FISH, and miniVIDAS are comparable. The analyses with the FISH technique are relatively complicated and require substantial experience in laboratory work, and methodical errors may easily occur. The procedure with the miniVIDAS analyzer, however, is less complicated and may be performed by a less-experienced laboratory worker.
Footnotes
Acknowledgments
This study was financially supported with the resources of the National Science Centre within the framework of project No. N 312 491340.
Disclosure Statement
No competing financial interests exist.