
Abstract
Stilbenoids have a broad range of beneficial health effects. On the other hand, the emergence of antibiotic-resistant Staphylococcus aureus presents a worldwide problem that requires new antibiotics or nonantibiotic strategies. S. aureus produces α-hemolysin (a pore-forming cytotoxin) that has been implicated in the pathogenesis of sepsis and pneumonia. Furthermore, the biofilms formed by S. aureus constitute a mechanism of antimicrobial resistance. In this study, we investigated the hemolytic and antibiofilm activities of 10 stilbene-related compounds against S. aureus. trans-Stilbene and resveratrol at 10 μg/mL were found to markedly inhibit human blood hemolysis by S. aureus, and trans-stilbene also inhibited S. aureus biofilm formation without affecting its bacterial growth. Furthermore, trans-stilbene and resveratrol attenuated S. aureus virulence in vivo in the nematode Caenorhabditis elegans, which is normally killed by S. aureus. Transcriptional analysis showed that trans-stilbene repressed the α-hemolysin hla gene and the intercellular adhesion locus (icaA and icaD) in S. aureus, and this finding was in line with observed reductions in virulence and biofilm formation. In addition, vitisin B, a stilbenoid tetramer, at 1 μg/mL was observed to significantly inhibit human blood hemolysis by S. aureus.
Introduction
S
The Gram-positive pathogen Staphylococcus aureus is a leading causes of community- and hospital-acquired infections (Lowy, 1998). S. aureus can secrete several exotoxins, such as, α-hemolysin, enterotoxins, coagulase, TSST-1, and protein A, all of which are associated with specific diseases (Ohlsen et al., 1997). In particular, α-hemolysin (Hla; also known as α-toxin) is a key virulence factor that has been implicated in the pathogeneses of sepsis, pneumonia, and severe skin infections (Bhakdi and Tranum-Jensen, 1991; Wilke and Bubeck Wardenburg, 2010). S. aureus preferentially attaches to various surfaces using a self-produced extracellular polymeric matrix, called a biofilm, and is protected by this biofilm from antibiotics and the host immune system (Costerton et al., 1999). Accordingly, biofilm formation plays a critical role in antibiotic resistance (Stewart and Costerton, 2001).
S. aureus is an exceptionally adaptable to resist diverse antibiotics (Lowy, 1998), and the long-term usage of antibiotics have resulted in the generation of multidrug resistant S. aureus (Levy and Marshall, 2004; Cegelski et al., 2008). Therefore, a nonantibiotic strategy is required, and this could be provided by antivirulence compounds that target bacterial virulence rather than cell viability, which may be less prone to develop drug resistance (Hentzer et al., 2002; Cegelski et al., 2008). Antivirulence strategies developed against S. aureus involve the inhibitions of enterotoxins and hemolysins (Wyatt et al., 2010), the inhibition of biofilm formation (Boles and Horswill, 2008; Iwase et al., 2010), and the disruption of bacterial quorum sensing (Park et al., 2007).
Recently, it was reported that resveratrol inhibits biofilm formation by Propionibacterium acnes (Coenye et al., 2012), Vibrio cholera (Augustine et al., 2014), Escherichia coli O157:H7, and Salmonella Typhimurium (Selma et al., 2012; Lee et al., 2013b). Therefore, in this study, we investigated the abilities of diverse stilbene-related compounds to reduce the hemolytic activity of and biofilm formation by S. aureus. In addition, we used an in vivo Caenorhabditis elegans model to study the effects of stilbenes on S. aureus virulence, and transcriptional analysis to elucidate the molecular mechanisms involved.
Materials and Methods
Bacterial strain and chemicals
S. aureus strain ATCC 6538 and methicillin-resistant S. aureus (MRSA) were used in this study. Initially, S. aureus ATCC 6538 was the study focus because the strain forms robust biofilms and because the strain, unlike some MRSA strains, has a strong hemolytic activity against human red blood cells. Experiments were conducted at 37°C in Luria-Bertani (LB) medium for ATCC 6538 and LB with 0.2% glucose for MRSA. Ten stilbene-related compounds (biphenyl, catechin, dicinnamyl, diphenyl methane, oxyresveratrol, pyrocatechol, resorcinol, resveratrol, cis-stilbene, and trans-stilbene) were purchased from Sigma-Aldrich (St. Louis, MO), and five resveratrol oligomers (namely, ɛ-viniferin, suffruticosol A, suffruticosol B, vitisin A, and vitisin B) were obtained from the Korea Chemical Bank (
Red blood hemolysis assay
Hemolysis was quantified as described previously with some modification (Larzábal et al., 2010). Human red blood cell lysis efficacies were measured using whole cultures of S. aureus grown in the presence of the test compounds. Briefly, S. aureus cells were diluted at 1:100 in LB medium and cultured with or without test compounds for 16 h at 250 rpm. Cell cultures (including cells and culture supernatants) were then added to diluted human red blood cells (previously separated by centrifugation at 890×g for 2 min and washed with PBS buffer 3 times) in PBS buffer (330 μL red blood cells/10 mL PBS buffer). To determine hemolytic activities, mixtures of blood and S. aureus (100 μL of cell culture) were incubated at 37°C for 1 h at 250 rpm. Supernatants were collected by centrifugation at 16,600×g for 10 min and absorbance was measured at 543 nm.
Crystal-violet biofilm assay
A static biofilm formation assay was performed in 96-well polystyrene plates (SPL Life Sciences, Korea) as previously described (Lee et al., 2011). Briefly, cells were inoculated into LB medium (total volume 300 μL) at an initial turbidity of 0.05 at 600 nm and cultured with or without each test compound for 24 h without shaking at 37°C. To quantify total biofilm formation, biofilms were stained with crystal violet, dissolved in 95% ethanol, and absorbances were measured at 570 nm (A570). Cell growths in 96-well plates were also measured at 620 nm (A620). Results are the averages of at least 12 replicate wells.
Confocal laser microscopy
S. aureus ATCC 6538 cells were cultured in 96-well plates and stained with carboxyfluorescein diacetate and succinimidyl acetate (Invitrogen, Molecular Probes, Inc., Eugene, OR) (Weston and Parish, 1990). Stained S. aureus ATCC 6538 biofilms were visualized using confocal laser microscopy (Nikon Eclipse Ti, Tokyo, Japan) with an Ar laser (excitation wavelength 488 nm, and emission wavelength 500–550 nm) and a 20×objective. Color confocal images were constructed using NIS-Elements C version 3.2 (Nikon eclipse). For each experiment, at least five random positions in four independent cultures were chosen for microscopic analysis.
C. elegans killing assay
The C. elegans killing assay was performed as described previously (Lee et al., 2013a). In brief, 10 μL of an overnight S. aureus ATCC 6538 culture was spread onto nematode growth medium plates with or without a test compound, and L4/young adult fer-15; fem-1 (Murphy et al., 2003) worms (n=20) were infected by placing them on the lawns. The nematodes were incubated at 25°C and scored as alive or dead on a daily basis by gently touching them with a platinum wire. Worms that crawled onto the walls of culture plates were eliminated from the analysis. Four independent experiments were conducted. C. elegans survival was determined using the Kaplan–Meier method, and the significances of survival differences were determined using the log-rank test (STATA6; STATA, College Station, TX). Statistical significance was accepted for p values<0.05.
RNA isolation and real-time quantitative reverse transcription–polymerase chain reaction (qRT-PCR)
To examine gene expressions, S. aureus ATCC 6538 cells were incubated with and without trans-stilbene (100 μg/mL) and incubated for 8 h with shaking at 250 rpm. Before taking samples, RNase inhibitor (RNAlater; Ambion, TX) was added and cells were immediately chilled for 30 s with dry ice and 95% ethanol (to prevent RNA degradation) and centrifuged at 16,600 g for 2 min. Cell pellets were then immediately frozen in dry ice and stored at −80°C. Total RNA was isolated using a Qiagen RNeasy mini Kit (Valencia, CA). To remove all DNA, purified RNA was treated for 15 min with 30 U of DNase I. qRT-PCR was used to investigate the transcriptions of α-hemolysin gene (hla), intercellular adhesion locus (icaA and icaD), proteases genes (aur and clp), quorum-sensing gene (agrA), accessory regulator A (sarA), enterotoxin (seb), and RNA polymerase sigma factor (sigB). The primer pairs used for qRT-PCR are presented in Supplementary Table 1 (Supplementary Data are available online at
Results
Antihemolytic activities of stilbene-related compounds
Initially, 10 commercially available stilbene-related compounds were tested at a concentration of 10 μg/mL to determine their abilities to reduce human blood hemolysis by S. aureus ATCC 6538 (Fig. 1). Notably, three stilbenes (resveratrol, cis-stilbene, and trans-stilbene) markedly inhibited hemolytic activity, and four stilbene-related compounds (catechin, dicinnamyl, oxyresveratrol, and pyrocatechole) had no significant effect (Fig. 1A). trans-Stilbene and resveratrol were found to dose-dependently reduce hemolysis (Fig. 1B and C), and at 10 μg/mL to inhibit hemolysis by more than 85% and 90%, respectively.
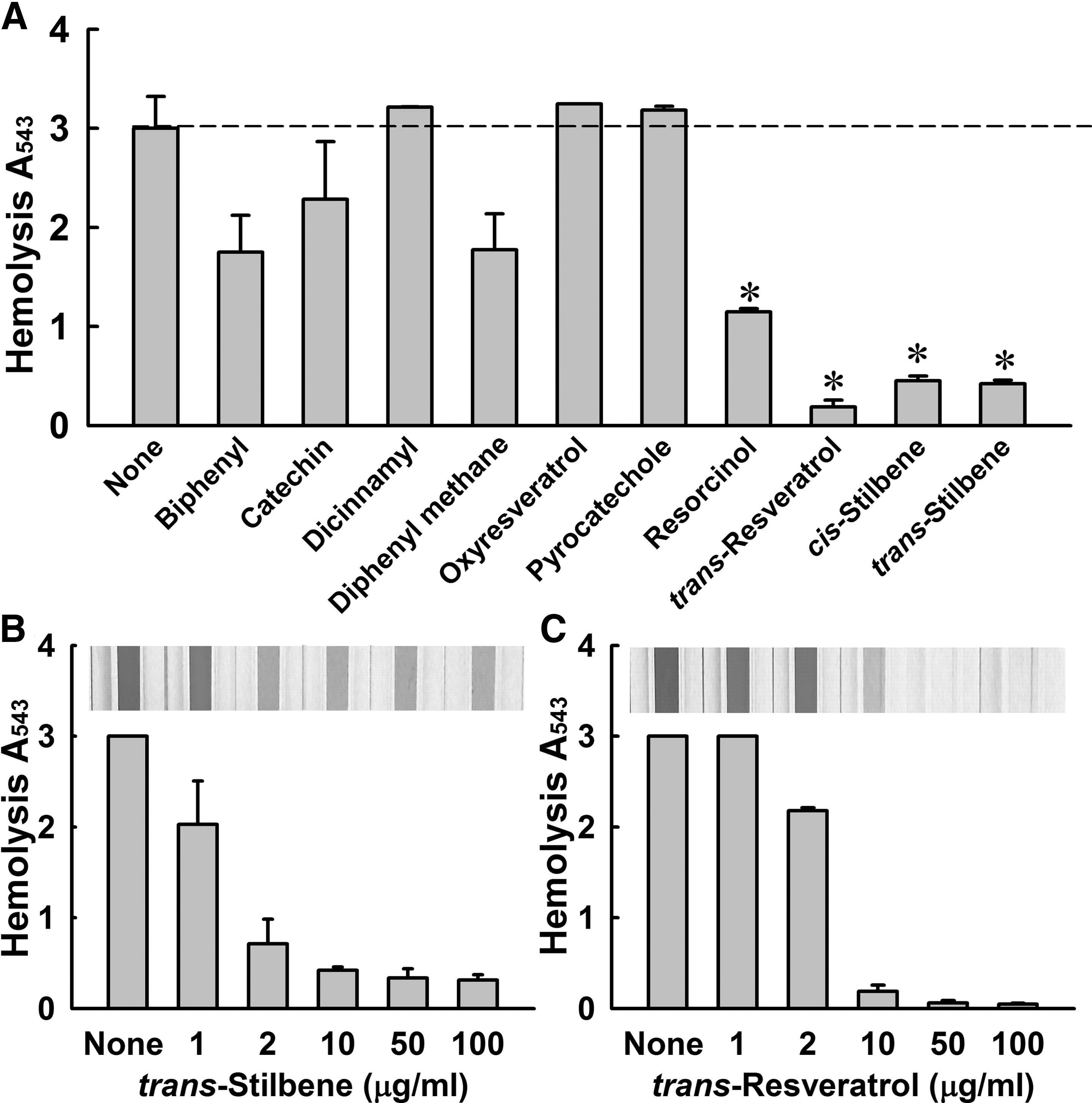
Antihemolytic activities of stilbenes. Human blood hemolysis by Staphylococcus aureus ATCC 6538 was quantified in the presence of stilbenes (10 μg/mL) after incubation for 16 h
Antibiofilm activities of stilbene-related compounds against S. aureus
The effects of 10 stilbene-related compounds (Fig. 2) were also investigated on S. aureus biofilm formation. Initially, the compounds were tested at a concentration of 100 μg/mL. Dicinnamyl, oxyresveratrol, resorcinol, and trans-stilbene significantly inhibited S. aureus biofilm formation, whereas resveratrol had no significant effect (Fig. 3A). In particular, biofilm formation was dose-dependently reduced by trans-stilbene concentrations up to 200 μg/mL (Fig. 3B). Biofilm reduction by trans-stilbene was also examined by confocal microscopy (Fig. 3D), and this examination confirmed the results of the quantitative biofilm assay. Similar to the model strain (S. aureus ATCC 6538), the biofilm formation of MRSA was dose-dependently inhibited by trans-stilbene and resveratrol, but not by cis-stilbene (Supplementary Fig. S1). Also, the hemolytic activity of MRSA was dose-dependently decreased by only resveratrol, but not by trans-stilbene and cis-stilbene (Supplementary Fig. S2). Since trans-stilbene dose-dependently reduced both biofilm formation and blood hemolysis, trans-stilbene was focused on the further study for the action mechanism.
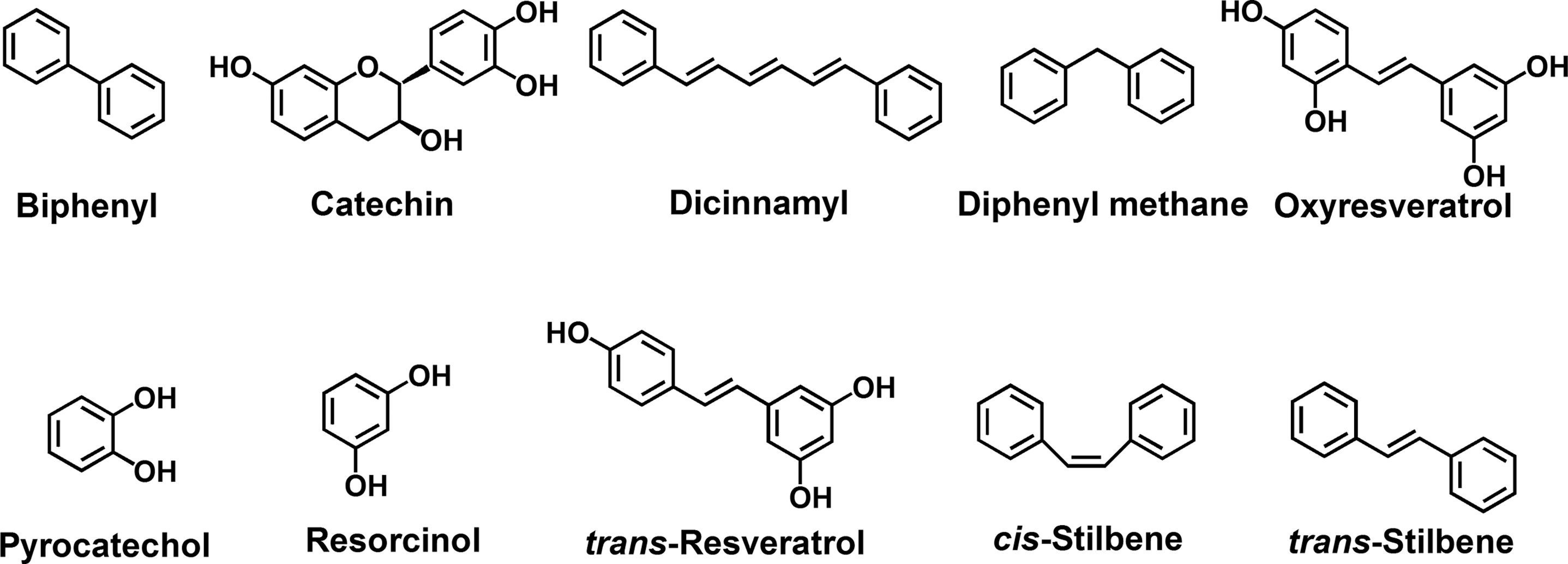
The stilbene-related compounds used in this study.
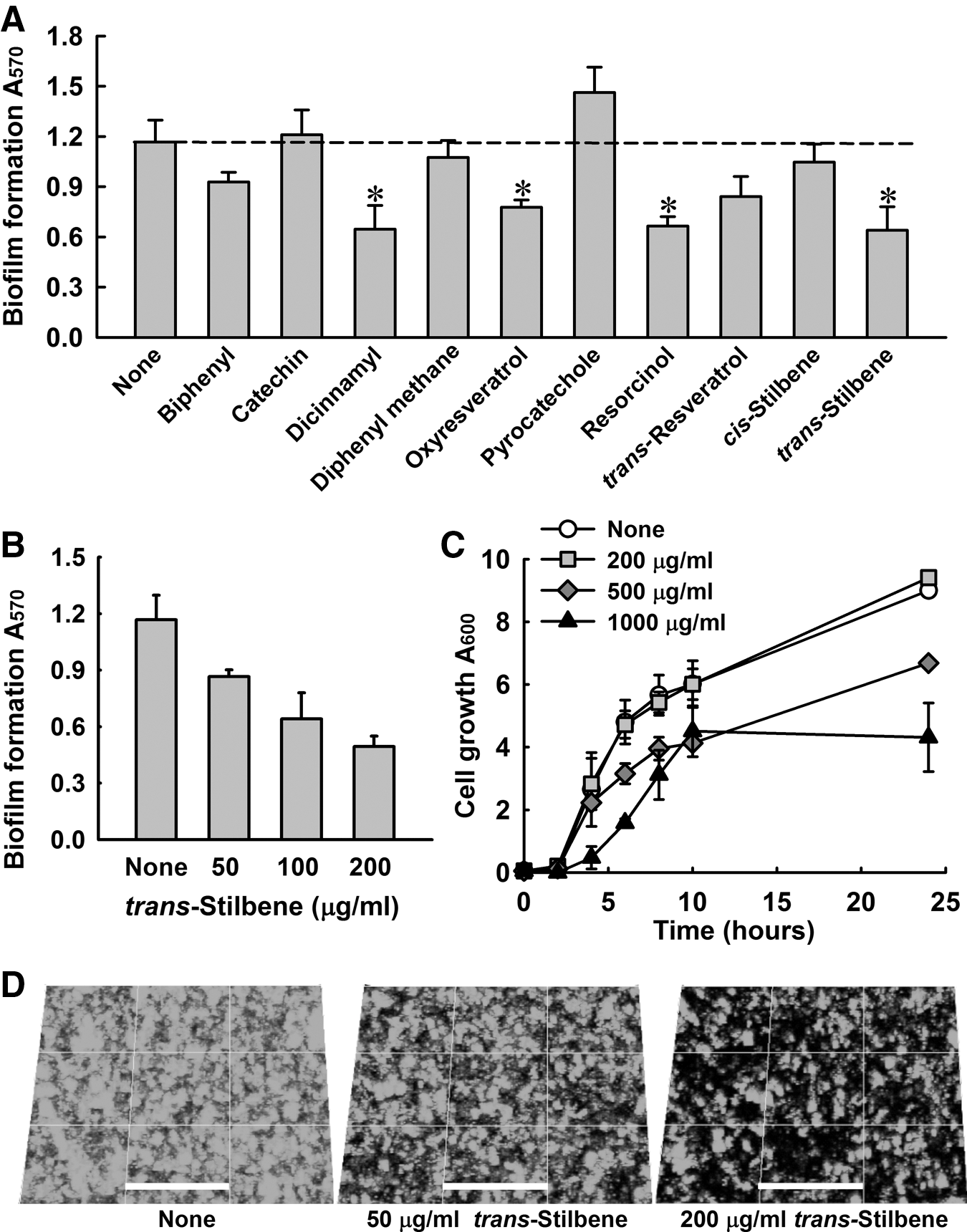
Effects of stilbenes on Staphylococcus aureus biofilm formation. Biofilm formation by S. aureus ATCC 6538 was quantified in the presence of stilbenes (100 μg/mL) after 24 h in 96-well plates
The cell growth of S. aureus was investigated because we wanted to investigate the antivirulence activity of trans-stilbene in the absence of bactericidal activity. trans-Stilbene at concentrations of up to 200 μg/mL did not reduce S. aureus ATCC 6538 cell growth, but at 1000 μg/mL delayed cell growth (Fig. 3C). This result suggests that the observed reductions in biofilm formation caused by trans-stilbene were due to its antibiofilm activity and not to its antimicrobial activity.
trans-Stilbene and resveratrol increased the survival of C. elegans exposed to S. aureus
Since S. aureus kills the nematode C. elegans (Sifri et al., 2003), a C. elegans killing assay was used to determine the effects of trans-stilbene and resveratrol on the survival of infected C. elegans. Both compounds were found to dose-dependently prolong C. elegans survival (Fig. 4), indicating that trans-stilbene and resveratrol reduce the virulence of S. aureus ATCC 6538. Additionally, C. elegans survival was investigated with E. coli OP50, which is a common food source to the nematode. As expected, there was no harmful effect of trans-stilbene (100 μg/mL) on the nematode (data not shown).
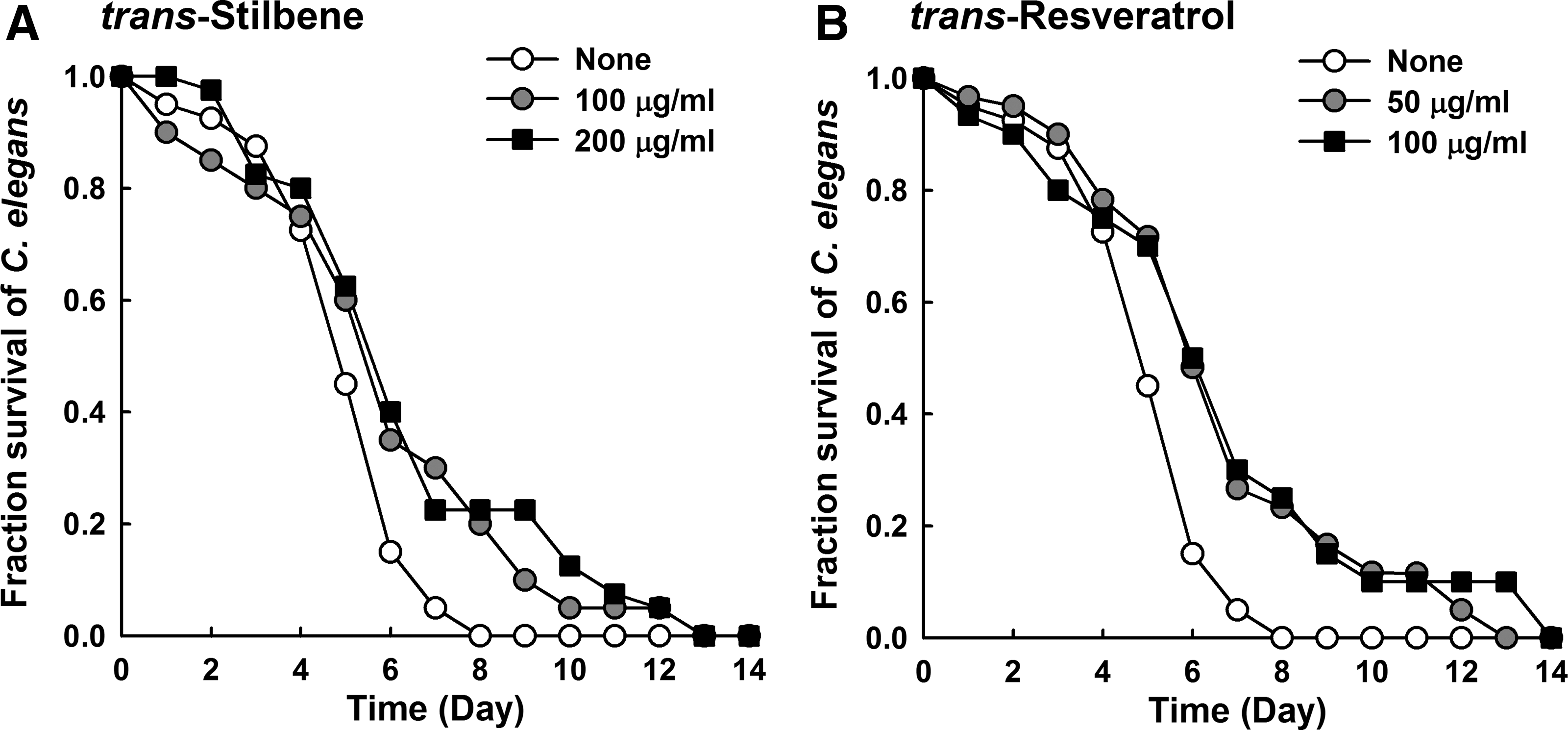
Effects of trans-stilbene and resveratrol on the survival of nematodes infected with Staphylococcus aureus. Solid killing assays of Caenorhabditis elegans strain fer-15; fem-1 infected with S. aureus ATCC 6538 in the presence of trans-stilbene
trans-Stilbene modulated the expressions of virulence and biofilm related genes in S. aureus
To investigate the mechanism underlying the antivirulence and antibiofilm activities of trans-stilbene, real-time qRT-PCR was used to examine the differential expressions of virulence factor–related genes and important regulatory genes in S. aureus (Fig. 5). Of several regulatory genes, three (hla, icaA, and icaD) were significantly repressed by trans-stilbene. In particular, the repression of α-hemolysin gene (hla) supported the observed reduction in hemolysis by S. aureus (Fig. 1). In addition, it appeared that trans-stilbene inhibited S. aureus biofilm formation by specifically downregulating intercellular adhesion locus genes (icaA and icaD), rather than other biofilm-related genes (protease genes, agrA quorum-sensing gene, and other regulatory genes) (Fig. 5).
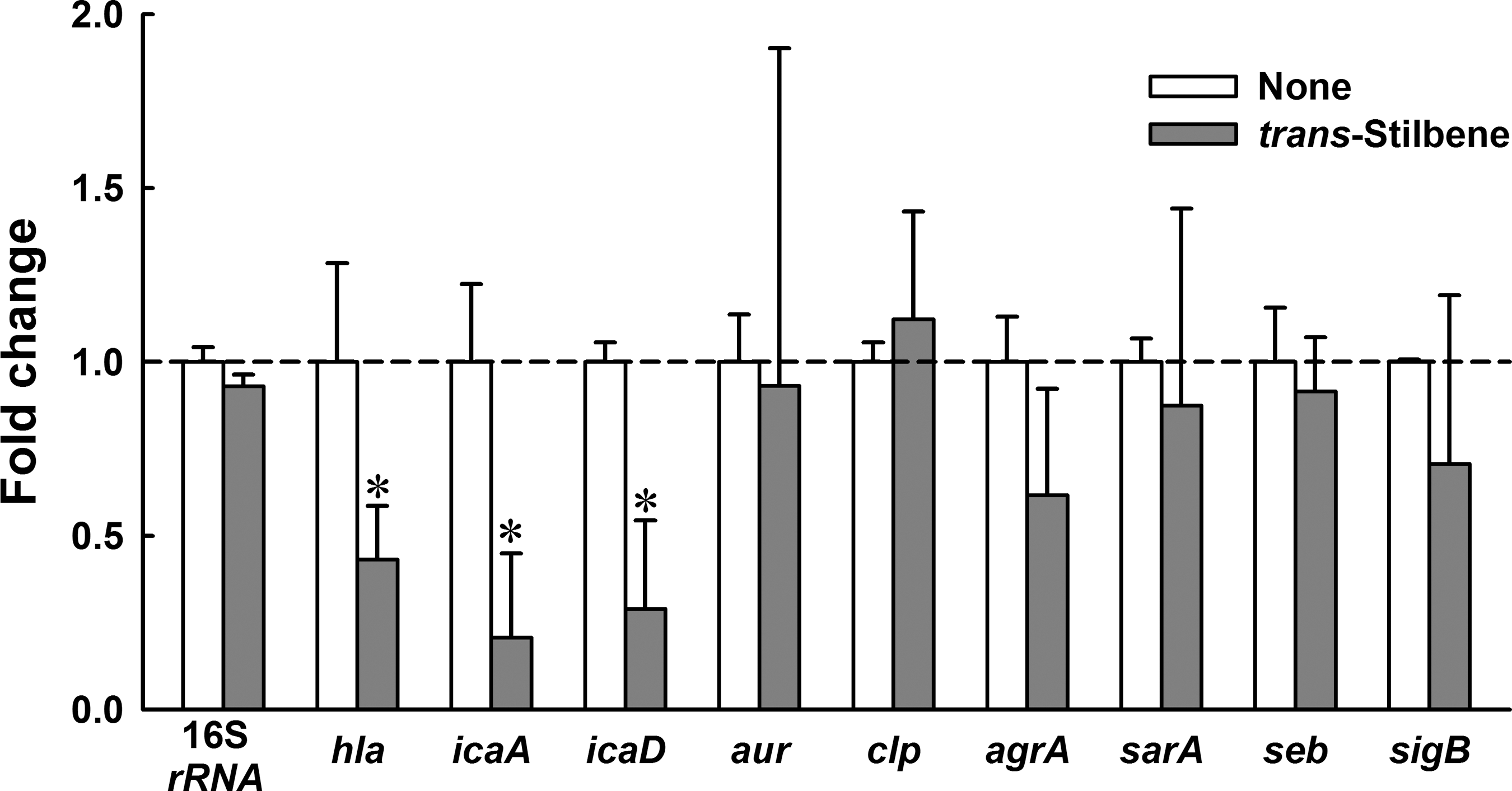
Transcriptional profiles of Staphylococcus aureus cells treated with or without trans-stilbene. S. aureus ATCC 6538 was cultivated to an A600 of 1 and then incubated with or without trans-stilbene (100 μg/mL) for 8 h. Transcriptional profiles were measured by real-time quantitative reverse transcription–polymerase chain reaction. Fold change represents changes (n-fold) in transcription of treated S. aureus versus untreated S. aureus (value 1.0). The experiment was performed in duplicate. *p<0.05 versus the untreated control.
Antihemolytic activity of stilbenoid oligomers
Because stilbenoid oligomers are widespread (Sotheeswaran and Pasupathy, 1993; Zgoda-Pols et al., 2002), we investigated the antihemolytic activities of five resveratrol oligomers (namely, ɛ-viniferin, suffruticosol A, suffruticosol B, vitisin A, and vitisin B). As expected based on the result in Figure 2, 3 of the 5 exhibited substantial antihemolytic activity at 10 μg/mL (Fig. 6). Vitisin B, a stilbenoid tetramer, was the most active of the five, and at 1 or 2 μg/mL inhibited hemolysis by more than 70% or 95%, respectively.
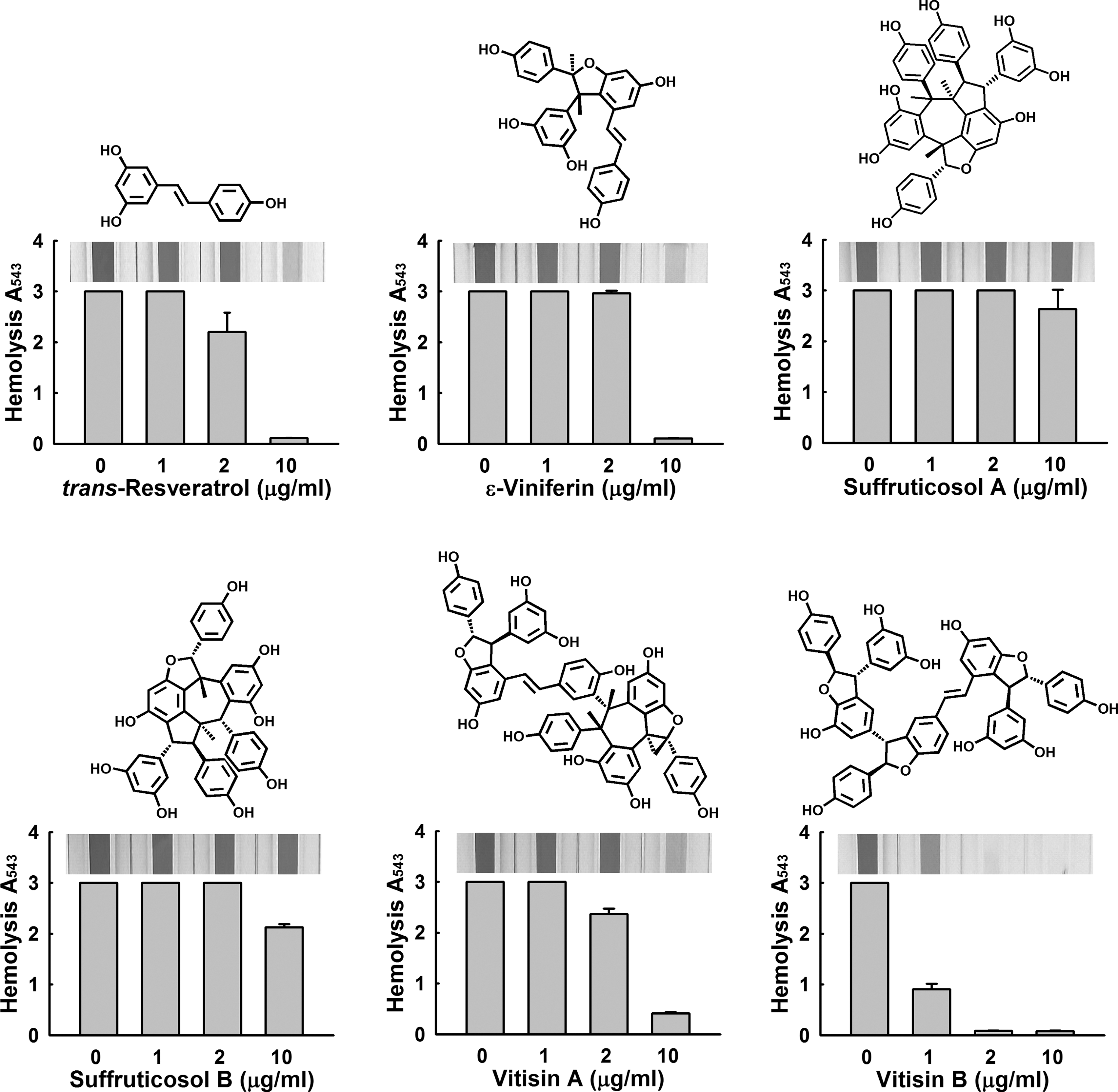
Antihemolytic activity of resveratrol oligomers. Human blood hemolysis by Staphylococcus aureus ATCC 6538 was quantified in the presence of five resveratrol oligomers (0, 1, 2, and 10 μg/mL) after incubation for 16 h. Pictures of spectrophotometer cuvettes are shown.
Discussion
In this study, we performed dual screenings using diverse synthetic and natural stilbene-related compounds to determine their abilities to inhibit the hemolytic activity of and biofilm formation by S. aureus. This study demonstrates for the first time that trans-stilbene exhibits antihemolytic, antibiofilm, and antivirulence activities in S. aureus in a dose-dependent manner.
One of objectives of this study was to identify the structural motif present in trans-resveratrol responsible for antivirulence activity in S. aureus. Hemolysis data suggests that the stilbene backbone is important for reducing hemolysis by S. aureus, because only three of the ten compounds tested (resveratrol, cis-stilbene, and trans-stilbene) had a significant effect (Fig. 1A). Furthermore, the stilbenoid dimer ɛ-viniferin and its tetramers vitisin A and vitisin B also exhibited high antihemolytic activity (Fig. 6).
Staphylococcal biofilms are encased in an extracellular matrix composed of proteins, polysaccharides, and extracellular DNA, and polysaccharide intercellular adhesion is due to linear β-1,6-linked glucosaminylglycans, which are synthesized by enzymes encoded by the ica operon. Hence, the ica operon is a key component of S. aureus biofilm formation (Cramton et al., 1999). Our transcriptional analysis supports the notion that trans-stilbene downregulates the ica operon, rather than downregulating agrA quorum-sensing or other regulatory systems (Fig. 5). It would be interesting to investigate the effect of stilbenes on S. aureus quorum sensing and how stilbene analogs enter and work in S. aureus cells.
The level of α-hemolysin (Hla) has been reported to be correlated with S. aureus virulence (Bubeck Wardenburg and Schneewind, 2008), and thus, the inhibition of hemolysis is an accepted antivirulence strategy (Wyatt et al., 2010). Furthermore, biofilm formation contributes to the virulence of S. aureus infection because it is associated with inherent resistance to antibiotics and to host immune response (Stewart and Costerton, 2001). It has also been reported that an hla deletion mutation significantly attenuates C. elegans killing by S. aureus (Sifri et al., 2003), and that staphylococcal biofilm protects against C. elegans immune defense (Begun et al., 2007). The present study shows that trans-stilbene attenuates S. aureus virulence in the nematode C. elegans (Fig. 4), as evidence by its inhibition of hemolysis (Fig. 1) and biofilm formation (Fig. 3), and the repression of the hla gene and ica operon (Fig. 5).
Conclusions
In the wake of rising antimicrobial resistance among S. aureus strains, novel strategies are required to control this pathogen. The results of the present study show that stilbene, and its natural derivatives, such as resveratrol and vitisins, inhibit S. aureus hemolytic activity and biofilm formation. As resveratrol is a well-characterized natural compound with known beneficial effects in humans, we propose that stilbene and its derivatives be regarded potential starting points for the development of novel antivirulence strategies. We envisage that these compounds could be used alone or in combination with other antimicrobial agents to prevent and treat infectious diseases caused by persistent S. aureus.
Footnotes
Acknowledgments
The five stilbenoid derivatives used in this study were kindly provided by the Korea Chemical Bank at the Korea Research Institute of Chemical Technology. This work was supported by the 2013 Yeungnam University Research Grant and by the Human Resources Development program (No. 20124030200100) of the Korea Institute of Energy Technology Evaluation and Planning (KETEP) grant funded by the Korea government Ministry of Trade, Industry and Energy.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.