
Abstract
The antimicrobial potential of cauliflower, broccoli, and okara byproducts was assessed against Gram-positive and Gram-negative bacteria. Salmonella enterica serovar Typhimurium, Escherichia coli O157:H7, Bacillus cereus, and Listeria monocytogenes serovar 4b growth behavior was assessed under exposure to 5% vegetable byproducts added to the reference medium, buffered peptone water (0.1% [wt/vol]), at 37°C. Although the byproducts were not effective against L. monocytogenes, they were bactericidal against Salmonella Typhimurium, E. coli O157:H7, and B. cereus. The most promising results were achieved with the cauliflower–Salmonella Typhimurium combination, because the bacterial population was reduced by 3.11 log10 cycles after 10 h of incubation at 37°C as a result of 5% cauliflower addition. Further studies were carried out for this combination, at different cauliflower concentrations (0, 0.5, 1, 5, 10, and 15%) and at temperatures in the range of 5–37°C. The greatest inactivation level (6.11 log10 cycles) was achieved at refrigeration temperature (5°C) using 15% cauliflower addition. Both temperature and cauliflower concentration significantly (p≤0.05) influenced the Salmonella Typhimurium inactivation level. The kinetic parameters were adjusted to mathematical models. The modified Gompertz mathematical model provided an accurate fit (root-mean-square error (RMSE) [0.00009–0.21] and adjusted-R2 [0.81–0.99]) to experimental Salmonella Typhimurium survival curves describing inactivation kinetics of the pathogen to the antimicrobial effect of cauliflower byproduct.
Introduction
E
The valorization of agriculture and food byproducts is a requirement of the European Union (EUROSTAT, 2010) supporting sustainable development. Vegetable residues are inexpensive, available in large amounts, and characterized by high dietary fiber content (Stojceska et al., 2008). So far, some valuable applications of these agrifood wastes involve animal feedstocks, fertilizers, paper industry application, extraction of essential oils and fragrances, composting, bioconversion, and new ingredients in product formulations (Henningsson et al., 2004).
For new product formulations, vegetable byproducts could be a valuable source of nutritional and antimicrobial compounds. Among them, there are two very important plant families, Brassicaceae and Fabaceae. Broccoli and cauliflower, the main crops of the Brassicaceae family, and soybean, the main crop of Fabaceae, contain phytochemical components with reported antioxidant and anticarcinogenic properties (Tyug et al., 2010). Worldwide production of broccoli and cauliflower was 22,226,957 tons in 2009. About 75% of this production belongs to China and India (USDA, 2009). The antioxidant properties of these vegetables could have a significant impact in the field of nutraceuticals and in food-processing industry applications (Cabello-Hurtado et al., 2012), mainly because of the polyphenol and glucosinolate contents (O'Shea et al., 2012).
With regard to Fabaceae, soybean is one of the most commonly consumed legumes in the world, with 200 million tons produced per year (FAOSTAT, 2010). Currently, the main producer is the United States (32%), followed by Brazil (28%) and Argentina (21%) (Nahashon and Kilonzo-Nthenge, 2011). After extraction of water from soybeans to produce soymilk and tofu, a byproduct called okara is obtained. Consequently, scientific and industrial research is required to find potential applications of okara from environmental and food technology viewpoints (O'Toole, 1999).
In this context, and for valorization purposes, the antimicrobial effect of vegetable byproducts from the raw material of broccoli (Brassica oleracea italica), cauliflower (Brassica oleracea botrytis), and soybean (Glycine max) was evaluated against Gram-positive and Gram-negative foodborne pathogens.
Materials and Methods
Bacterial cultures and growth conditions
Pure cultures of Listeria monocytogenes serovar 4b (CECT 4032), Bacillus cereus (CECT 131), Salmonella enterica serovar Typhimurium (CECT 443) and Escherichia coli O157:H7 (CECT 5947) were provided freeze-dried by the Spanish Type Culture Collection. The B. cereus culture was rehydrated with 10 mL of brain heart infusion (BHI) broth (Scharlab Chemie, Barcelona, Spain), whereas tryptic soy broth (TSB) (Scharlab Chemie) was used for E. coli O157:H7, Salmonella Typhimurium and L. monocytogenes rehydration. After 20 min, the rehydrated culture was transferred to 500 mL of BHI broth or TSB, respectively, and incubated at 32°C for B. cereus and at 37°C for the other microorganisms, with continuous shaking (Selecta Unitronic) at 200 rpm for 14 h to obtain cells in a stationary growth stage. Growth curves were obtained by plate count (colony-forming units per mL [CFU/mL]). The cells were centrifuged (Beckman Avanti J-25) twice at 4000×g at 4°C for 15 min and then resuspended in BHI broth or TSB, respectively. After the second centrifugation, the cells were resuspended in 20 mL of BHI broth or TSB with 20% glycerol, and then dispensed in 2-mL vials to a final concentration of 108 obtained by plate count. The 2-mL samples were immediately frozen and stored at −80°C until needed for the kinetic inactivation studies.
Antimicrobial substances
Cauliflower, broccoli, and okara byproducts from vegetable raw materials were provided as leaf residues from primary production. Each raw byproduct was tested to screen its bacteriological quality. The bacteriological analysis determined the presence/absence of microbial contamination and was carried out according to the procedures described by Aycicek et al. (2006). The samples studied presented positive contamination with L. monocytogenes and B. cereus (Gram positives), chiefly in broccoli and cauliflower samples, below 5 CFU/g. In contrast, no samples were contaminated with E. coli O157:H7 or Salmonella Typhimurium (Gram negatives).
The raw byproduct was washed in sterile water to eliminate contaminating substances, dried, triturated, and homogenized using a laboratory grinder (Janke & Kunkel Ika-Labortechnik) to obtain a powder with a particle size of 40 μm, which was used to perform the experiments (Brandi et al., 2006).
Total phenolic compounds of vegetable byproducts
The total phenol contents of the vegetable byproducts were determined spectrophotometrically according to the Folin–Ciocalteu colorimetric method (Singleton and Rossi, 1965). Gallic acid calibration standards with concentrations of 0, 100, 200, 300, 400, 500, 600, 700, 800, and 1000 ppm were prepared. Three milliliters of sodium carbonate solution (2% [wt/vol]) (Sigma-Aldrich Co. LLC) and 100 μL of Folin–Ciocalteu reagent (1:1 [vol/vol]) (Sigma-Aldrich Co. LLC) were added to an aliquot of 100 μL from each gallic acid standard (Sigma-Aldrich Co. LLC) or sample tube. The mixture was vortexed (Heidolph) and allowed to stand at room temperature in the dark for 1 h. Absorbance was measured at 750 nm using a Lan Optics Model PG1800 spectrophotometer (Labolan, Spain), and the results were expressed as milligrams of gallic acid equivalents per liter.
Substrate and inoculation
Buffered peptone water (Scharlab Chemie) (0.1% [wt/vol]) was used as reference substrate in the present study (Pina-Pérez et al., 2007; Lin et al., 2010). The reference medium was supplemented with natural vegetable byproduct and later was inoculated with the stock culture to obtain a final concentration of 107 CFU/mL. In an initial research step, the antimicrobial potential of each byproduct (cauliflower, broccoli, and okara) was tested against the microorganisms studied under specific conditions: (1) 5% (wt/vol) of vegetable byproduct addition to medium, and (2) at the optimal incubation temperature for each microorganism, for 10 h. The plates of B. cereus were incubated at 30°C during 48 h on BHI agar (Scharlab Chemie); the plates of E. coli O157:H7 and Salmonella Typhimurium were incubated at 37°C during 24 h on tryptic soy agar (TSA); and the plates of L. monocytogenes were incubated at 37°C during 48 h on TSA.
A second experimental step was conducted, based on the results obtained in the first one. The vegetable byproduct with the greatest bactericidal effect was tested over a wide concentration range (0–15% [wt/vol]) against the most sensitive microorganism. Moreover, to test the influence of temperature on the antimicrobial potential of the vegetable byproduct studied, incubation was carried out at four temperatures (5, 10, 22, and 37°C).
Viable microorganism count
At regular time intervals (hours), the cell suspension for each sample was evaluated by plate count after serial dilution with 0.1% (wt/vol) buffered peptone water. Each dilution was plated in duplicate. Experiments were carried out in triplicate and the plate counts were used for enumeration (CFU/mL).
Modeling of microorganism inactivation
Microbial behavior was fitted to a modified Gompertz equation to mathematically describe the bacterial inactivation kinetics under the intervention of the most effective vegetable byproduct at different concentrations and temperatures (Linton et al., 1995):
where N is the cell concentration at time t (CFU/mL), N0 is the initial cell concentration (CFU/mL); C is the difference between upper and lower value of asymptote; B is the relative death rate at time M, and M being the time at which the absolute death rate is maximal. Minus sign before C signifies the microbial inactivation.
Subsequently, with B, C and M obtained values, the maximum death rate (μmax) can be calculated as follows:
Data analysis and model evaluation
The statistical analysis was performed with STATGRAPHICS Centurion XV (version 15.1.03; STATGRAPHICS, Warrenton, VA). This analysis included average and standard deviation calculations for the three replications and an analysis of variance analysis to test significant differences depending on incubation conditions. The goodness of fit of the model was assessed by using adjusted regression coefficient (adjusted-R2) and RMSE (López et al., 2004).
Results and Discussion
Antimicrobial effect of vegetable byproducts against Gram-positive and Gram-negative bacteria
The antimicrobial effect of broccoli, cauliflower, and okara byproducts was evaluated. Figure 1 shows the survival curves obtained for (a) Gram-positive and (b) Gram-negative bacteria in 0.1% buffered peptone water supplemented/not supplemented with vegetable byproduct at 5% concentration. As can be seen in the figure, the vegetable byproducts studied exerted a bactericidal effect with reduction of the bacterial population over time when incubation was carried out at optimal growth temperature for each microorganism, with the exception of L. monocytogenes. Among the Gram-positive bacteria, L. monocytogenes was not affected by exposure to the vegetable byproducts (5%, 37°C, 10 h), with no significant reduction of the initial load (p>0.05); in contrast, the B. cereus population was reduced in the range [0.61–2.32] log cycles by vegetable byproduct intervention under the same conditions. In the case of the Gram-negative bacteria, Salmonella Typhimurium was highly sensitive compared with E. coli O157:H7 under exposure to Brassicas, but was more resistant to the antimicrobial effect of okara than E. coli O157:H7.
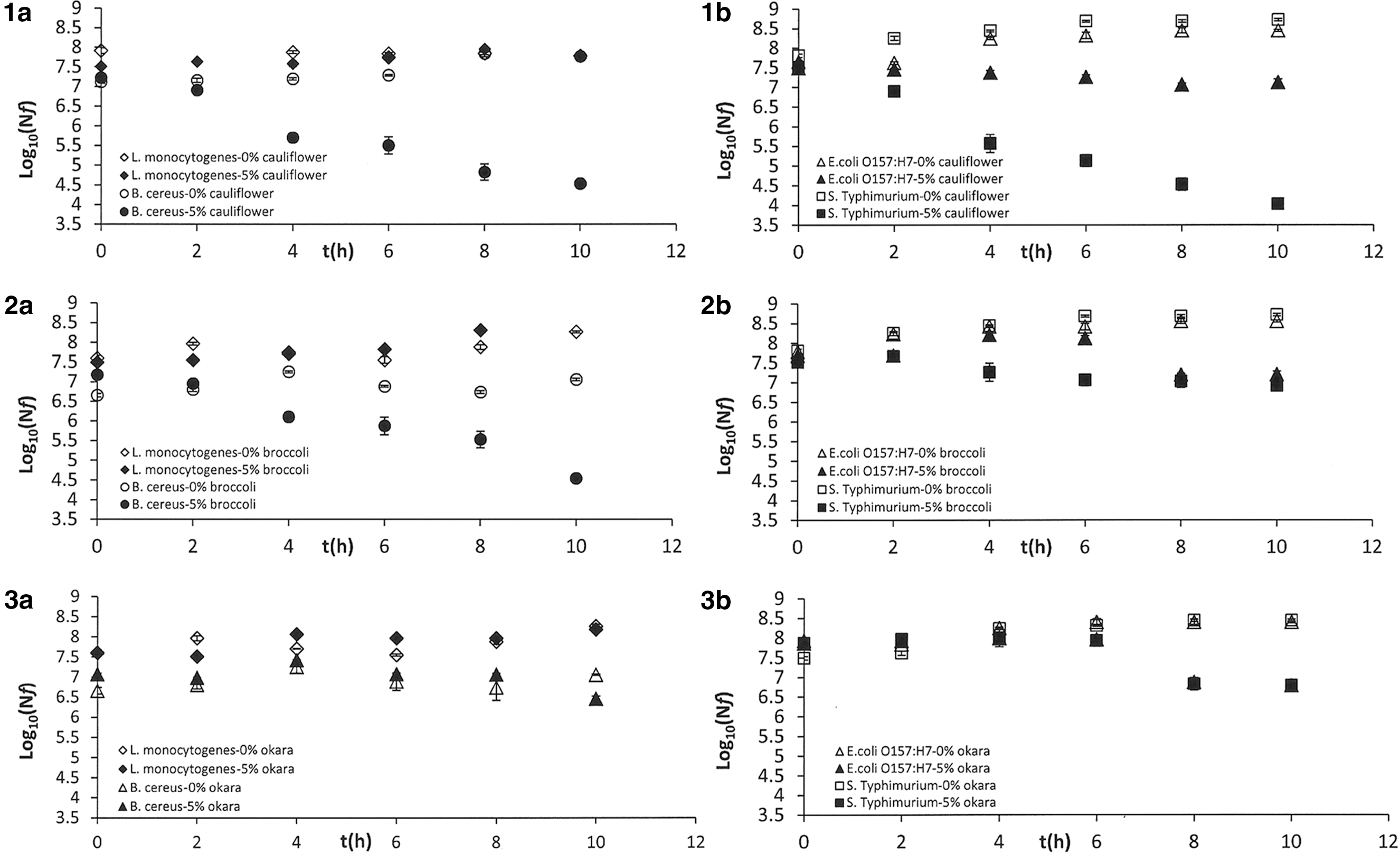
Survival curves of Gram-positive bacteria (Listeria monocytogenes and Bacillus cereus)
Many studies have supported that Gram-positive bacteria were more susceptible to antimicrobial effect of plant essential oils and extracts than Gram-negative bacteria (Smith-Palmer et al., 1998; Jayaprakasha et al., 2003). In contrast, other studies have found more sensitivity against other natural extracts or essential oils in Gram-negative than Gram-positive bacteria (Hu et al., 2004; Di Pasqua et al., 2005).
Figure 2 shows the inactivation levels achieved for each by product–microorganism combination after the complete incubation period. Under the conditions studied, the bactericidal effect against E. coli O157:H7, Salmonella Typhimurium, and B. cereus achieved a minimum value of 0.48±0.05 log10 cycles reduction under broccoli intervention against E. coli O157:H7, and a maximum value of 3.11±0.50 log10 cycles reduction by the effect of cauliflower against Salmonella Typhimurium. From the results obtained, it is possible to establish a ranking based on the sensitivity of each microorganism to the antimicrobial effect of the byproducts studied. With regard to the microorganisms' susceptibility to the antimicrobial effect of cauliflower, the ranking can be established as follows: Salmonella Typhimurium (3.11±0.15 log10 cycles reduction)>B. cereus (2.31±0.025 log10 cycles reduction)>E. coli O157:H7 (0.53±0.09 log10 cycles reduction). For broccoli, the sensitivity of the microorganisms studied can be ordered as follows: B. cereus (2.25±0.05 log10 cycles reduction)>Salmonella Typhimurium (0.49±0.225 log10 cycles reduction)>E. coli O157:H7 (0.48±0.05 log10 cycles reduction). With regard to the sensitivity of the microorganisms studied to the antimicrobial effect of okara, E. coli O157:H7 and Salmonella Typhimurium seemed to produce similar results (1.15±0.05 and 1.07±0.025 log10 cycles reduction, respectively), followed by B. cereus (0.61±0.03 log10 cycles reduction).

Effect of 5% cauliflower, broccoli, and okara concentration on inactivation levels of Listeria monocytogenes, Escherichia coli O157:H7, Salmonella enterica serovar Typhimurium, and Bacillus cereus cells after 10 h at 37°C. *Statistically significant inactivation (p-value<0.01).
According to the results obtained, under the conditions studied (1) okara was the most bactericidal byproduct against E. coli O157:H7 (1.15 log10 cycles); (2) cauliflower and broccoli showed the highest antimicrobial effect against B. cereus (2.25 and 2.31 log10 cycles, respectively), and (3) cauliflower was the most effective vegetable byproduct against Salmonella Typhimurium (3.11 log10 cycles). So it is possible to conclude that addition of cauliflower at 5% achieved the greatest reduction in bacterial levels, showing the most bactericidal capability among the vegetable byproducts studied.
Although the antioxidant capacity of Brassicaceae and Fabaceae is widely known and has been attributed mainly to their polyphenol contents (Tyug et al., 2010; O'Shea et al., 2012), so far the antimicrobial effect of these plants has scarcely been studied (Hu et al., 2004). With respect to the soybean byproduct, okara, there are previous studies indicating antimicrobial effect of soybean derivatives (O'Toole, 1999; Roubos-Van den Hil et al., 2010).
As far as we know, the antimicrobial capability of these raw agrifood byproducts has not previously been reported. To our knowledge, only qualitative studies of the antimicrobial potential of other vegetable byproducts have been carried out, establishing a correlation between functional properties attributed to these vegetables and their polyphenol contents (Fattouch et al., 2007; Roubos-Van den Hil et al., 2010). The observed bactericidal capability of cauliflower, broccoli, and okara byproducts against the foodborne pathogens studied could be due to the effect of their high polyphenol content.
As can be seen in Table 1, cauliflower extract had the highest polyphenol content (milligrams gallic acid per liter), followed by extract of broccoli and finally, okara extract. A similar order followed for their antimicrobial action as well. Therefore, it is possible to establish a relationship between the polyphenol content of tested byproducts and their antimicrobial activity.
Milligrams gallic acid per liter.
Effect of temperature and concentration of cauliflower against Salmonella Typhimurium
In view of the results obtained, an intensive study was conducted on the antimicrobial effect of cauliflower (the most effective vegetable byproduct), based on its effect on the most sensitive microorganism, Salmonella Typhimurium. Various concentrations of cauliflower (0, 0.5, 1, 2, 5, 10, and 15%) were assessed against Salmonella Typhimurium at different temperatures: 5°C (refrigeration), 10°C (abuse in refrigeration), 22°C (room temperature), and 37°C (optimal temperature).
For all the temperatures, the higher the concentration of cauliflower, the greater the reduction of cell population (p≤0.05). Figure 3 shows the concentration effect of cauliflower at 5°C. As can be seen graphically, the Salmonella Typhimurium cell population was reduced by 0.6, 0.93, 1.88, and 2.86 log10 cycles at concentrations of 0, 0.5, 1, and 2%, respectively, and about 6 log10 cycles at concentrations of 5, 10, and 15%, reaching a maximum reduction level (6.11 log10 cycles) at the highest cauliflower concentration (15%). These results are in agreement with studies conducted by Brandi et al. (2006) on the antimicrobial potential of Brassica leaf juice in reference media. The influence of concentration level on the antimicrobial capability of Brassicaceae species was reported previously with the disc diffusion method (Sousa et al., 2008; Blazevic et al., 2010).
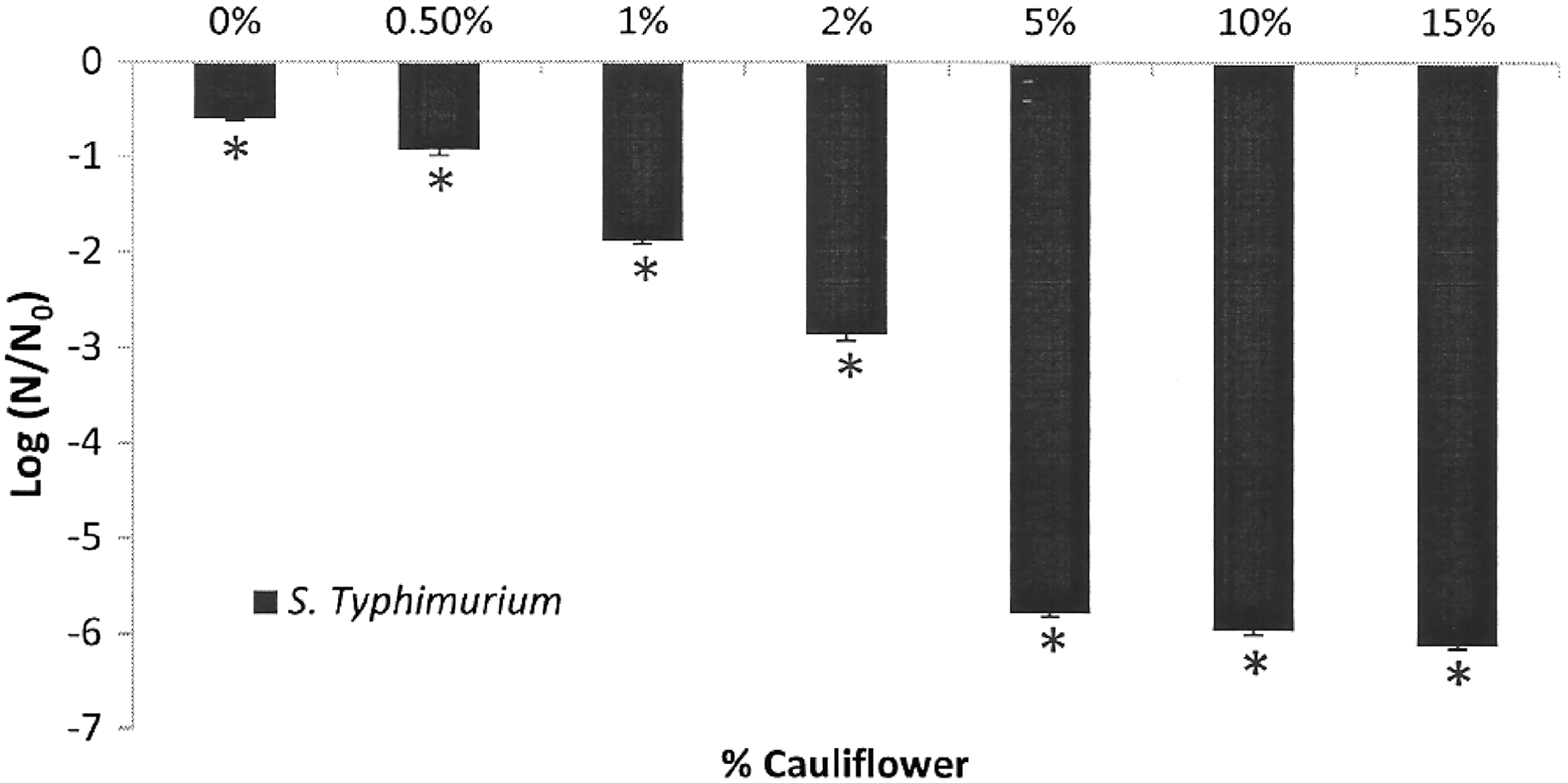
Effect of 0.5, 1, 2, 5, 10, and 15% cauliflower concentrations on inactivation levels of Salmonella enterica serovar Typhimurium in reference medium after 432 h at 5°C. *Statistically significant inactivation (p-value<0.01).
With regard to temperature effect, Figure 4 shows the reduction in the growth of the cell population due to temperature with respect to the growth behavior at 37°C, when cauliflower was added to the medium at 5%. When the temperature was reduced from 37 to 22°C, a reduction level of 0.20 log10 cycles was observed; reducing the temperature from 37 to 10°C achieved a reduction of 0.51 log10 cycles; and when the temperature was reduced from 37°C to refrigeration level (5°C), the reduction of bacterial counts was 1.19 log10 cycles. Therefore, under exposure to the same concentration of cauliflower byproduct, the lower the incubation temperature, the higher the bacterial reduction. Figure 4 also shows the concentration effect at different temperatures. It can be seen that cauliflower exerted a higher concentration effect at 5°C and 37°C than at 10°C and 22°C, with slightly more bactericidal effect of 5% cauliflower against Salmonella Typhimurium at 5°C than at 37°C. Therefore, the antimicrobial effect of cauliflower byproduct was higher at optimal growth temperature (37°C), such as occurs in other studies such as Brandi et al. (2006), and at refrigeration temperature (5°C), such as the results conducted by Cava et al. (2007), where the authors found that the antimicrobial activity of clove and cinnamon essential oils against L. monocytogenes also increased at lower temperature (7°C). A similar effect also was found by Smith-Palmer et al. (1998), Bahk et al. (1990), and Beuchat et al. (1994) with other plant extracts. Thus, they raise the possibility of applying these extracts in combination with refrigeration storage temperatures. Meanwhile, the enhanced antimicrobial capability of natural ingredients against several foodborne pathogens has been observed previously at refrigeration temperature in other studies (Ferrer et al., 2009; Iturriaga et al., 2012), which are in agreement with the present results.
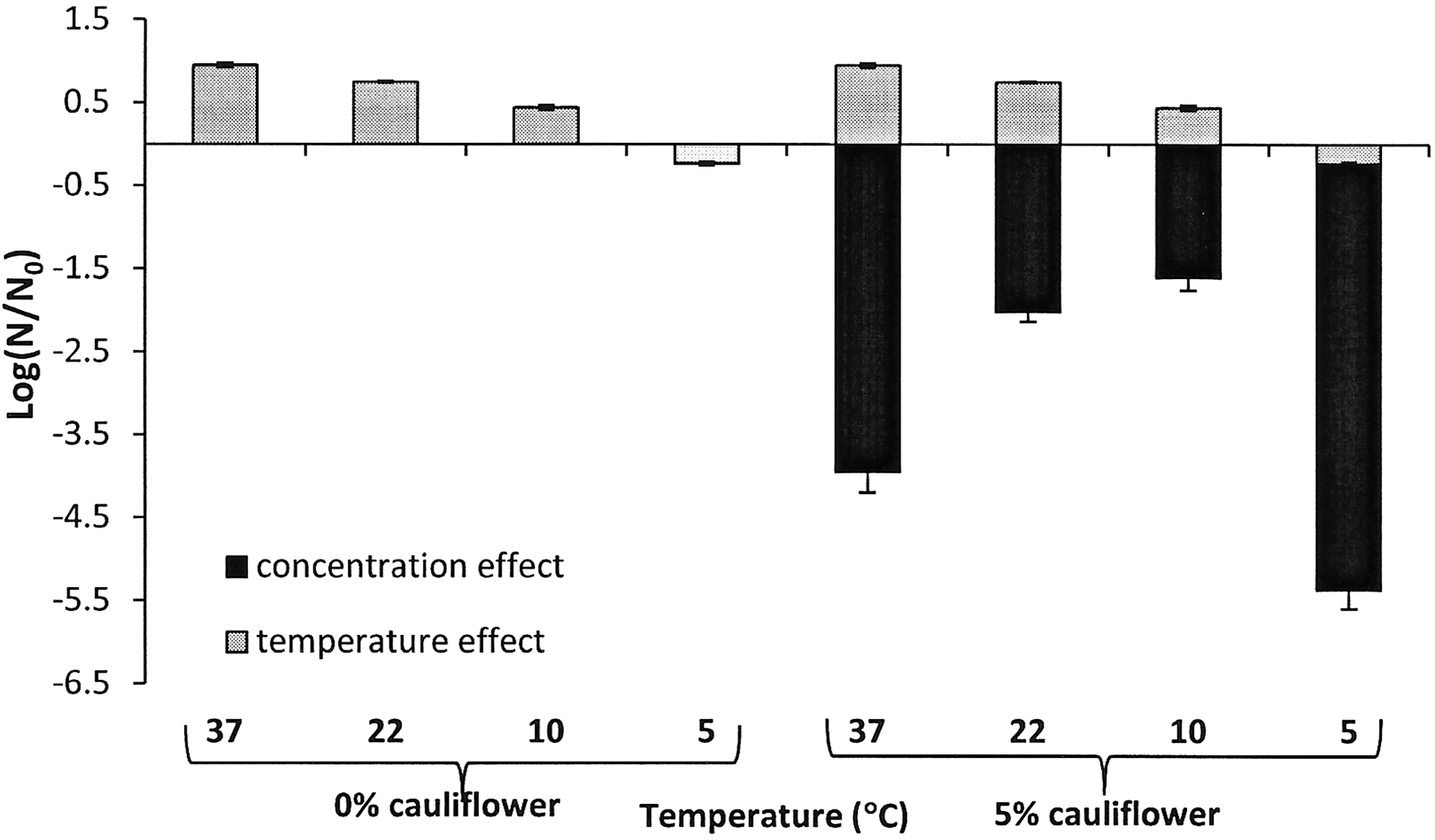
Effect of temperature on reduction in growth of initial cell population of Salmonella enterica serovar Typhimurium with respect to growth behavior at 37°C and cauliflower concentration effect at various temperatures studied.
Cauliflower extract added to reference medium at a concentration of 5% and incubated at optimal growth temperature (37°C) for 10 h not only inhibits Salmonella Typhimurium growth, but also reduces the microbial load levels by 4 log10 cycles. These results are in agreement with the results obtained by Brandi et al. (2006) against Salmonella spp. and E. coli spp. by adding 20% cauliflower extract (leaf juice) at the same temperature (37°C).
The results obtained were adjusted to the modified Gompertz distribution function, which has been used by other authors to provide accurate fits to microbial behavior under exposure to natural antimicrobials (Gammariello, 2008; Belda-Galbis, 2013). The Gompertz kinetic parameters are presented in Table 2, which shows negative values because they are kinetics of inactivation, with the exception of the 0% cauliflower −22°C combination, which has a positive value because the microorganism grows under these conditions. The μmax values, calculated with C, B, and M parameters, give us information about the maximum growth/dead rate. As can be seen in Table 2, μ values are generally higher at higher temperature and cauliflower extract concentration. Therefore, both temperature and concentration lead to an increase of the inactivation rate, contributing to the antimicrobial effect. To our knowledge, no previous studies have reported the mathematical modeling of microbial inactivation/survival using raw cauliflower extract. There are few studies on the antimicrobial effect of Brassicaceae vegetables, and most of them evaluated the antimicrobial activity qualitatively, using inhibition zones (Hu et al., 2004; Sousa et al., 2008; Blazevic et al., 2010).
R 2 and mean squared error (MSE) values are indicators of goodness of fit.
Conclusions
According to the results presented in this research work, cauliflower extract can be considered as a potential material with antimicrobial properties with important economic and food safety applications. However, animal feed supplementation, or development of new additives based on the bactericidal effect of this extract for vegetable creams and ready-to-eat meals, which are pasteurized and stored at refrigeration temperature (5°C), present important challenges to food processors.
Footnotes
Acknowledgments
M. Sanz-Puig is grateful to Consejo Superior de Investigaciones Científicas (CSIC) for providing a contract as a researcher working actively on an INNPACTO project entitled “Nuevos Productos para Alimentación, Obtenidos a Partir de la Valorización de Subproductos Hortofrutícolas” with reference: IPT-2011-1724-060000. M.C. Pina-Pérez is grateful to the CSIC for providing a Doctorate contract. The present research work has been funded by Ministry of Economy and Competitiveness and with Fondo Europeo de Desarrollo Regional (FEDER) funds.
Disclosure Statement
No competing financial interests exist.