
Abstract
We investigated the implied health benefits of retail chicken breast labeled as “organic” or “antibiotic-free” when compared to conventional products based on frequency of contamination by Salmonella spp., Campylobacter spp., and coliform bacteria resistant to fluoroquinolones, extended-spectrum cephalosporins, or carbapenems. We purchased 231 prepackaged chicken breasts from 99 grocery stores representing 17 retail chains in Ohio, Michigan, and Pennsylvania from June to September 2012. Ninety-six (41.5%) packages were labeled “antibiotic free” and 40 (17.3%) were labeled “organic,” with the remaining 95 (41.1%) making neither label claim. Salmonella were recovered from 56 (24.2%) packages, and the recovery rate was not different between product types. Five percent of packages contained Salmonella carrying the extended-spectrum cephalosporin resistance gene bla CMY-2, representing 21.4% of Salmonella isolates. Campylobacter spp. were recovered from 10.8% of packages, with observed recovery rates similar for the three product types. Using selective media, we recovered Escherichia coli harboring bla CMY-2 from over half (53.7%) of packages, with similar recovery rates for all product types. In addition, we recovered E. coli carrying bla CTX-M from 6.9% of packages, and E. coli with QRDR mutations from 8.2% of packages. Fluoroquinolone-resistant E. coli recovered using selective media were more common (p<0.05) in conventional (18.9%) compared to organic (0) and antibiotic-free (2.1%) packages. Our results indicate that, regardless of product type, fresh retail chicken breast is commonly contaminated with enteric pathogens associated with foodborne illness and commensal bacteria harboring genes conferring resistance to critically important antimicrobial drugs.
Introduction
F
While most cases of salmonellosis and campylobacteriosis are self-limiting local infections that do not require medical attention in otherwise healthy individuals (Kumar et al., 1982), a proportion will progress to systemic infections that require antimicrobial therapy in order to prevent patient death (Stoycheva and Murdjeva, 2006). Populations at increased risk of adverse outcomes include children, the elderly, and immunocompromised individuals (Acheson and Hohmann, 2001). Salmonella and Campylobacter present in fresh retail chicken products may also be resistant to human antimicrobial drugs important for therapeutic treatment of invasive Gram-negative bacterial infections. The 2011 NARMS reported that 22.4% of C. jejuni recovered from chicken breasts were resistant to ciprofloxacin, and that 33.5% of Salmonella spp. recovered from chicken breasts were resistant to ceftriaxone (FDA, 2011). Foodborne outbreaks of ciprofloxacin-resistant Campylobacter (Engberg et al., 2004) and ceftriaxone-resistant Salmonella (Dutil et al., 2010) associated with consumption of fresh retail chicken have been reported. Resistant infections may increase the likelihood of treatment failure, resulting in increased healthcare costs (Holmberg et al., 1987) and a greater risk of patient death (Helms et al., 2002).
Some retail chicken products are marketed as “organic” or “antibiotic free” with the implication that they are healthier than similar products without these label claims. As a result, consumers may reasonably expect that they are less likely to contain pathogens or antimicrobial-resistant organisms such as Escherichia coli. Any product labeled as organic must meet specific criteria for production practices established by the US Department of Agriculture (USDA) including no use of antimicrobial drugs (USDA, 2011). However, there are no requirements for production practices that can be expected when the term “antibiotic free” is present on food product labels. The available scientific literature suggests that fresh retail meat products labeled as organic or antibiotic free are microbiologically similar to products without those label claims (Smith-Spangler et al., 2012), although this has not yet been fully established. Our objective was therefore to determine the differences in contamination with pathogens and antimicrobial-resistant organisms between retail boneless chicken breasts labeled as organic or antibiotic free when compared to conventional products without those label claims.
Materials and Methods
Sampling
Prepackaged fresh boneless chicken breast with label claims of “organic” or “antibiotic-free” and conventional packages without either label claim were purchased from supermarkets in three US states between June and September 2012. A convenience sample of retail grocery stores was selected based on product availability, and only stores with boneless chicken breasts available representing at least two of these three label claims were sampled. A single package representing each of the available product types at each store was selected from available product in retail cases, purchased, and transported or shipped overnight to our laboratory at The Ohio State University.
Bacterial culture
For bacterial culture, one breast half per package was mixed in 275 mL buffered peptone water (BPW) and incubated overnight at 37°C. The following day, the chicken rinsate was aseptically inoculated to MacConkey agar and streaked for isolation. After overnight incubation at 37°C, a single E. coli isolate from each package was confirmed indole positive and stored for further characterization.
For the recovery of E. coli resistant to fluoroquinolones, 4 mL of incubated chicken rinsate was inoculated to 36 mL of nutrient broth containing 16 μg/mL nalidixic acid and incubated for 18–24 h at 37°C. This broth was then streaked onto MacConkey agar containing 2 μg/mL ciprofloxacin and incubated overnight. A single lactose-positive, indole-positive E. coli isolate from each positive package was reserved for further characterization.
E. coli resistant to extended-spectrum cephalosporins and carbapenems were cultured by adding 4 mL of incubated chicken rinsate to 36 mL nutrient broth containing 2 μg/mL cefotaxime (Mollenkopf et al., 2012). After overnight incubation, this broth was streaked onto MacConkey agar supplemented with 4 μg/mL cefepime to identify isolates with a bla CTX-M phenotype, onto MacConkey agar containing 8 μg/mL cefoxitin to identify isolates with a bla CMY-2 phenotype, and to MacConkey agar with 2 μg/mL meropenem to identify phenotypic carbapenem resistance.
For isolation of Salmonella spp. we transferred a 100-μL chicken breast/BPW rinsate aliquot to Rappaport-Vassiliadis R10 broth, incubated overnight at 42°C and differentially selected on xylose-lysine-Tergitol 4 agar (XLT-4) with overnight incubation (Heider et al., 2009). From each sample, bacteria from a single black colony on XLT-4 were isolated on MacConkey agar and confirmed as Salmonella using standard biochemical reactions including triple sugar iron agar, urea broth, and polyvalent antisera. Salmonella spp. isolates were screened for extended-spectrum cephalosporin, carbapenem, and fluoroquinolone resistance by inoculation onto selective agar (Mollenkopf et al., 2012).
For isolation of Campylobacter spp., 50 mL of the chicken breast/BPW rinsate was transferred before incubation to Whirl-pak bags with 50 mL of double-strength Bolton broth and incubated at 42°C for 48 h under microaerophilic conditions. We inoculated this broth to Campy-Cefex agar and incubated under microaerophilic conditions at 42°C for 48 h. Suspect Campylobacter isolates were confirmed and speciated using multiplex polymerase chain reaction (PCR) with 16S rRNA primers specific for Campylobacter genus, as well as for C. jejuni (mapA) and C. coli (ceuE) as previously described (Linton et al., 1997; Denis et al., 1999).
Isolate characterization and antimicrobial susceptibility testing
To examine the genetic similarity of Salmonella and Campylobacter isolates, pulsed-field gel electrophoresis (PFGE) genotyping (CHEF-DRIII; Bio-Rad Laboratories, Hercules, CA) was performed on total genomic DNA. Agarose plugs prepared with the Salmonella isolates were digested using XbaI (Promega, Madison, WI) following previously reported protocols (Ribot et al., 2006). After electrophoresis, banding patterns were compared and levels of similarity assigned using generally accepted criteria (Tenover et al., 1995). Salmonella isolates were compiled into pulsotypic groups by using the Dice coefficient similarity index and the unweighted pair-group method with arithmetic averages (UPGMA) with clustering settings of 1.00% optimization and 1.00% band position tolerance via Bionumerics software (Applied Maths, Kortrijik, Belgium). Salmonella isolates that appeared clonal on XbaI PFGE were compared by PFGE following digestion using SpeI to confirm their relatedness. Agarose plugs prepared with Campylobacter isolates were digested using SmaI (Promega, Madison, WI) and imaged as described (Ribot et al., 2001; Sanad et al., 2011). Similarity and clustering analysis were accomplished using the Dice coefficient and UPGMA with 1.00% optimization and 1.50% band position tolerance.
The resistance phenotype of a single Salmonella, Campylobacter, and E. coli recovered from nonsupplemented MacConkey media to represent each positive package was characterized by minimum inhibitory concentrations (MICs). Salmonella and E. coli MICs to a standard panel of 15 antimicrobial drugs were generated using a semiautomated broth microdilution system (NARMS CMV2AGNF, TREK Diagnostic Systems, Cleveland, OH) following Clinical and Laboratory Standards Institute (CLSI) guidelines (CLSI, 2009). Campylobacter MICs were generated to a standard panel of nine antimicrobial drugs (NARMS CAMPY, TREK Diagnostic Systems) as described (Sanad et al., 2011).
Discrimination of resistance genes
The quinolone-resistance determining regions (QRDR) corresponding to the gyrA and parC subunits of the fluoroquinolone-resistant E. coli isolates were amplified by PCR and bidirectionally sequenced to assess DNA-binding surface amino acid substitutions (Genewiz, South Plainfield, NJ) (Kato et al., 1990; Everett et al., 1996; Heisig, 1996). Additionally, E. coli isolates expressing fluoroquinolone resistance were tested by PCR for the carriage of plasmid-mediated qnr using a global qnr primer pair (Chen et al., 2012).
The presence of bla
CTX-M and bla
CMY-2 in extended-spectrum cephalosporin-resistant E. coli and Salmonella isolates was confirmed by PCR utilizing previously reported primer sets (Mollenkopf et al., 2012). bla
CTX-M genes were bidirectionally sequenced using the corresponding PCR amplification primers and analyzed using BLAST (
Plasmid characterization
The plasmid content of each bla CTX-M E. coli isolate was visualized by electrophoresis using a standard procedure (Kado and Liu, 1981). bla CTX-M-harboring plasmids were codified according to a PCR-based replicon typing procedure that detects 18 replicon types based on incompatibility group loci (Carattoli et al., 2005, 2006; Johnson et al., 2007).
Data analysis
Prevalence of each outcome of interest was expressed on a per-package basis. Differences in the likelihood of recovering specific pathogens or antimicrobial-resistant organisms between the three package types were estimated using logistic regression mixed models with SAS Proc GLIMMIX (SAS v. 9.3, SAS Institute, Cary, NC). Variables representing the state in which the package was purchased, vacuum packaging, weight of the package, and date of purchase were assessed in the models as fixed effects, while variables representing the individual grocery store, the retail grocery store chain, and the processing plant were assessed in the models as random effects in order to investigate and control for potential confounding. Multiple pairwise comparisons of least-square means were accomplished using the Tukey-Kramer method (Hayter, 1984).
Results
We purchased 231 packages of fresh retail boneless chicken breast from 99 grocery stores belonging to 17 store chains located in 3 US states between June and September 2012 (Table 1). Most packages (61%) were purchased from stores located throughout Ohio. Another 36% of packages were purchased from stores located throughout southern Michigan, and the 8 packages purchased in Pennsylvania were from stores in the Pittsburgh metropolitan area. The packages originated from 27 chicken processing plants, although the processing plant was not identifiable on the label of 9 (4%) packages. Ninety-six (41.5%) of the packages were labeled “antibiotic free” and 40 (17.3%) of the packages were labeled “organic” (Table 1). Most (83.1%) packages were foam trays covered with clear plastic stretch wrap and the remaining (16.9%) were in vacuum packaging.
Data are presented as frequency (proportion) of packages.
Includes chicken breast packages from 10 retail grocery store chains.
Includes chicken breast packages from 14 processing plants including 9 packages for which the processing plant could not be determined.
QRDR, quinolone-resistance determining regions.
We recovered Salmonella spp. from 56 (24.2%) of the packages of boneless chicken breasts (Table 1). Over 5% of the packages contained Salmonella carrying the extended-spectrum cephalosporin resistance gene bla CMY-2, representing over 21% of all Salmonella isolates. We also recovered Campylobacter spp. from 10.8% of the packages (Table 1), most of which (72%) were C. jejuni while the remainder (28%) were C. coli. We observed little variability in pathogen recovery rates between states, but there were considerable differences between grocery store chains and between processing plants, with Salmonella recovery rates as high as 41% for one large regional chain of grocery stores. We could not detect differences in recovery rates of Salmonella, Salmonella carrying bla CMY-2, or Campylobacter spp. among conventional, organic, or antibiotic-free chicken breasts.
We recovered E. coli harboring bla CMY-2 from over half (53.7%) of the packages using selective media (Table 1). In addition, we recovered E. coli carrying bla CTX-M-1 from 6.9% of packages, all of which were located on IncI1 plasmids. We also recovered E. coli with QRDR mutations conferring fluoroquinolone resistance from 8.2% of packages using selective media (Table 1). We did not recover carbapenem-resistant isolates. We could not detect differences among product types in the probability of recovering E. coli harboring either bla CMY-2 or bla CTX-M using selective media. Using Fisher's exact test, we were able to detect differences among the product types in the probability of detecting E. coli with QRDR mutations conferring fluoroquinolone resistance. However, because no organic packages contained E. coli with these QRDR mutations, multivariable models utilizing this outcome would not converge. As a result, we were unable to control for potential confounders and so cannot draw strong conclusions regarding this observed difference.
Distributions of MICs of Salmonella spp., Campylobacter spp., and commensal E. coli recovered without antimicrobial selection pressure are summarized in Figure 1. We could not detect differences in the proportion of isolates with reduced susceptibility or in the median MIC to each of the antimicrobial drugs tested between conventional, organic, or antibiotic-free packages of chicken breasts.
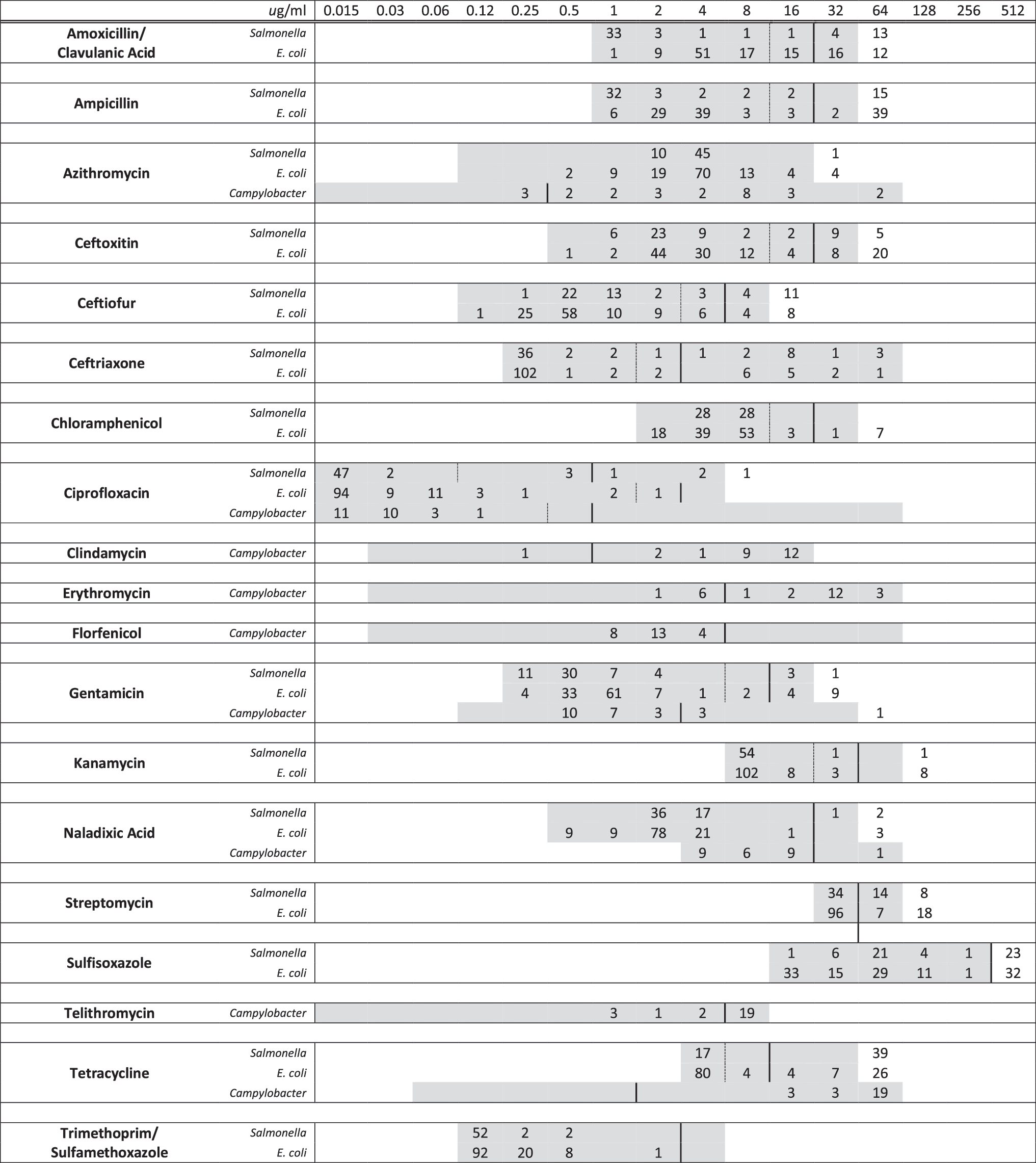
Distribution of minimum inhibitory concentrations (MICs) among Salmonella spp., Campylobacter spp., and commensal Escherichia coli recovered without antimicrobial selection pressure from 231 retail packages of fresh boneless chicken breasts purchased from grocery stores in 3 US states. (numbers of isolates are shown in the body of the table). Broken lines represent susceptible breakpoints and solid lines represent resistant breakpoints where available. Corresponding to the concentration listed at the top of each column (μg/mL), the included range of each antimicrobial is shown in gray. For amoxicillin/clavulanic acid, the indicated range refers to amoxicillin concentration; clavulanic acid was included in wells at half the amoxicillin concentration. For trimethoprim/sulfamethoxazole, the indicated range refers to trimethoprim; sulfamethoxazole was included in wells at 19 times the concentration of trimethoprim.
Analysis of the relatedness of the Salmonella isolates (Fig. 2) revealed multiple clonal or highly similar strains recovered from packages purchased in different stores located in different states, originating from different processing plants, and recovered at different times throughout the summer. In addition, multiple clonal or highly similar Salmonella strains were recovered from at least two of the three different product types. Three highly similar (>97%) Salmonella isolates were each recovered from different product types purchased from different store chains located in two states and originated from different processing plants. In two situations, clonal or highly similar Salmonella strains differed in their carriage of plasmid-borne bla CMY-2.
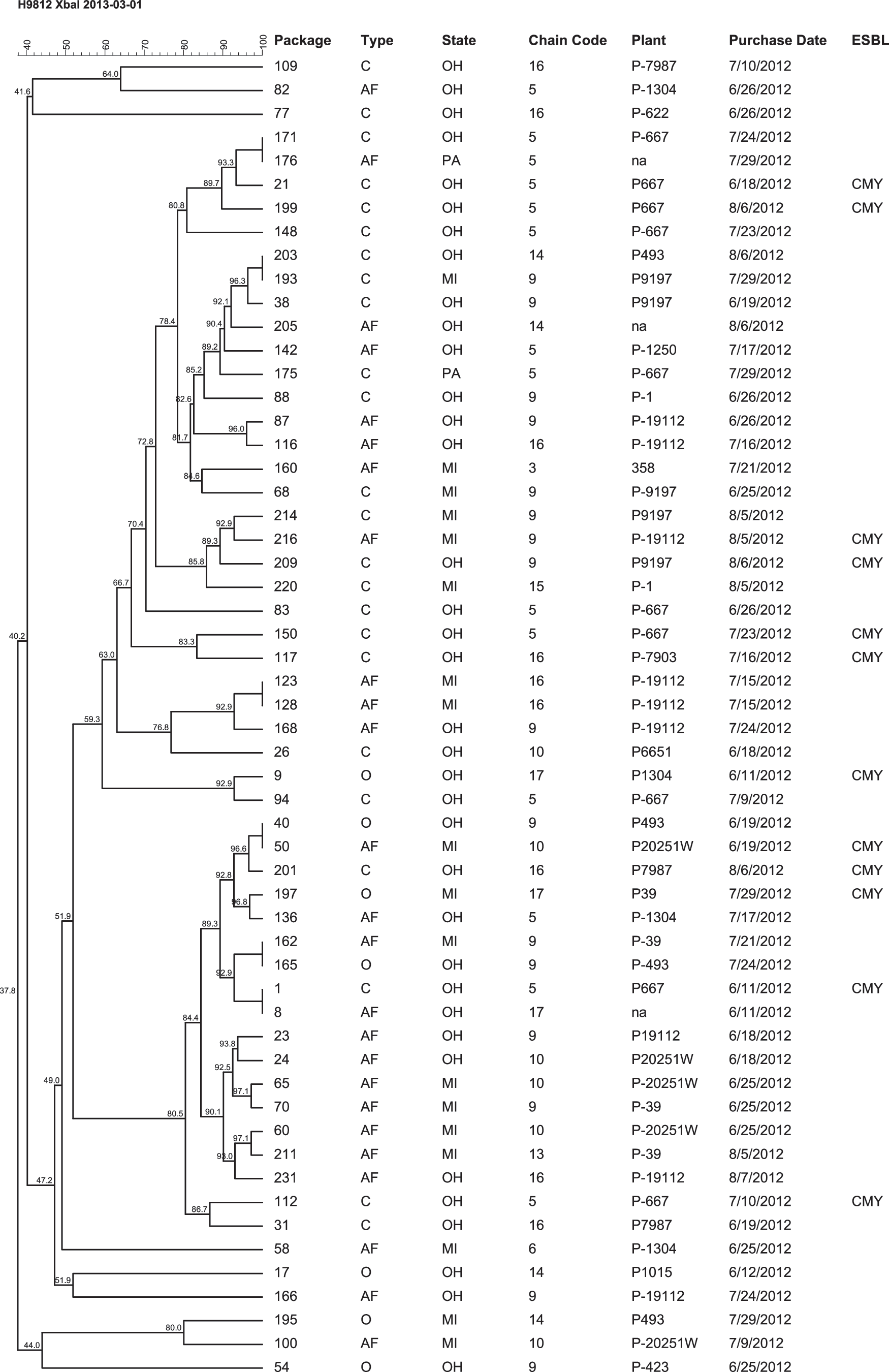
Dendrogram showing XbaI pulsed-field gel electrophoresis profiles for Salmonella spp. recovered from retail packages of fresh boneless chicken breasts purchased from grocery stores in 3 US states. For product type, C=conventional, O=organic, AF=antibiotic-free. ESBL, extended-spectrum β-lactamase.
Analysis of the relatedness of the C. jejuni (Fig. 3) and C. coli (Fig. 4) isolates also revealed highly similar strains recovered from different product types, different store chains, and originating from different processing plants. Two highly similar (>90%) C. jejuni isolates were recovered from conventional and antibiotic-free package types in different store chains located in Ohio and Michigan that originated from different processing plants. Highly similar (>95%) C. coli isolates were also recovered from different product types purchased from different store chains, and originating from different processing plants.
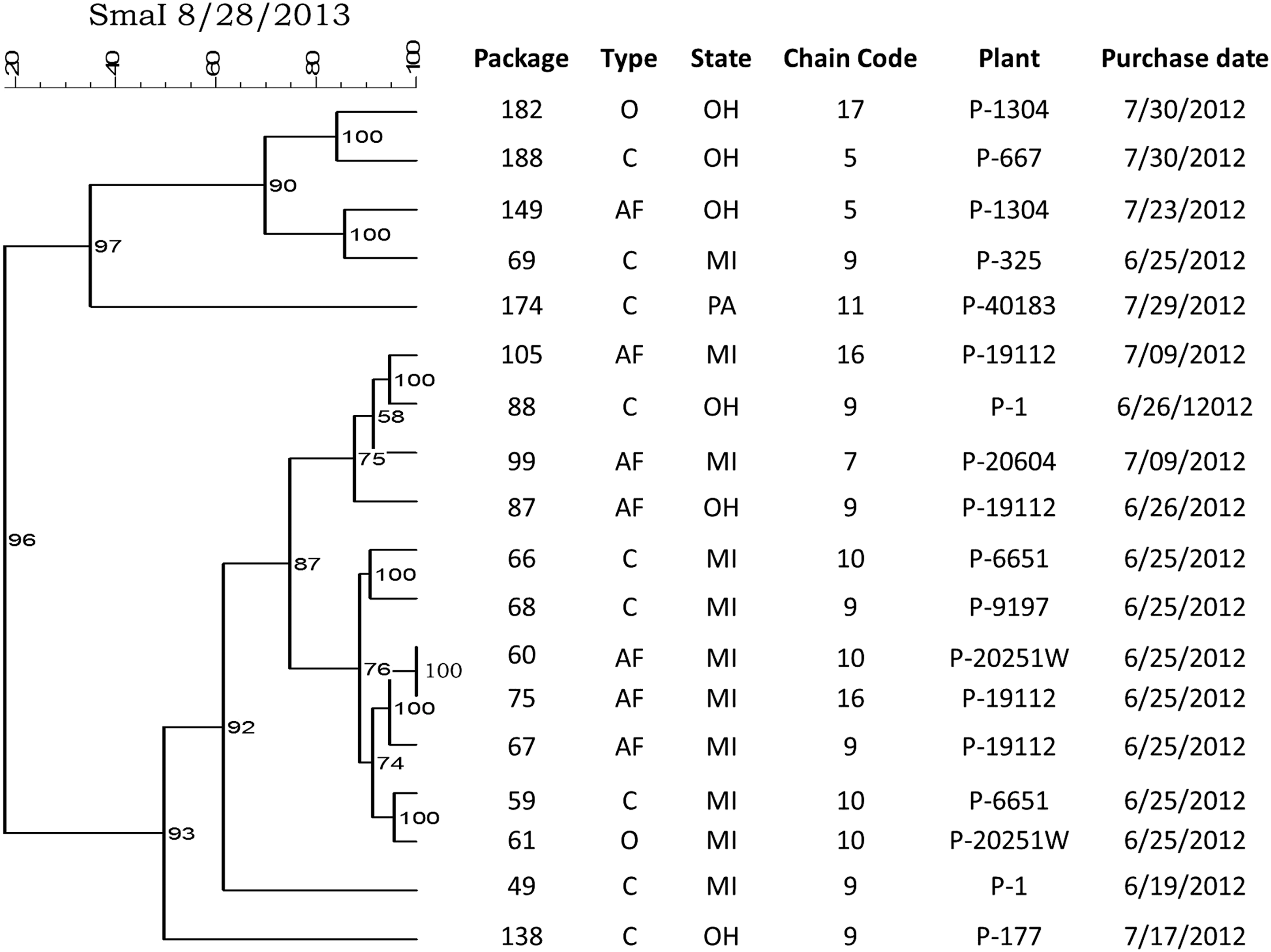
Dendrogram showing SmaI pulsed-field gel electrophoresis profiles for 18 Campylobacter jejuni recovered from retail packages of fresh boneless chicken breasts purchased from grocery stores in 3 US states. For product type, C=conventional, O=organic, AF=antibiotic-free.
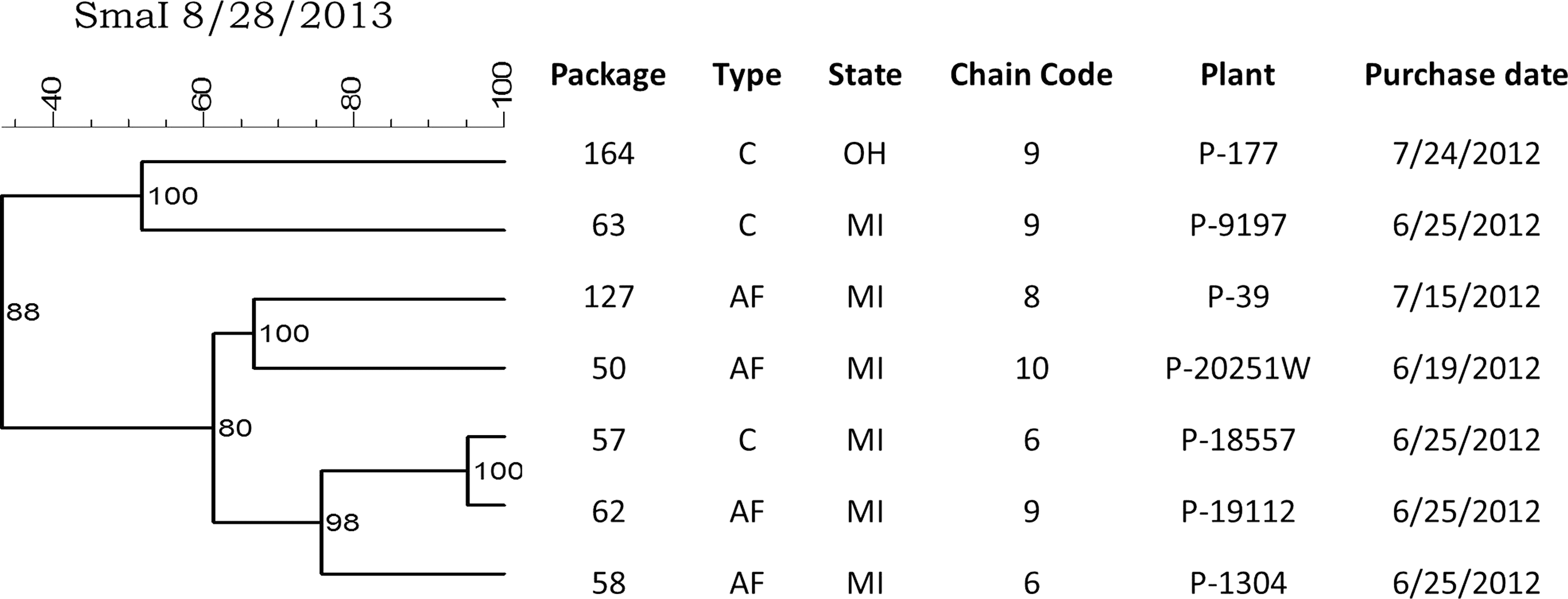
Dendrogram showing SmaI pulsed-field gel electrophoresis profiles for seven Campylobacter coli recovered from retail packages of fresh boneless chicken breasts purchased from grocery stores in 3 US states. For product type, C=conventional, AF=antibiotic-free.
Discussion
We found little difference in the microbiological quality of fresh boneless chicken packages labeled as organic or antibiotic free compared to conventional packages without those label claims. Salmonella spp., including multidrug-resistant strains, frequently contaminated all three product types. Commensal E. coli carrying bla CMY-2 and bla CTX-M on transferable plasmids were also frequently recovered for all three package types. A recent review of the scientific literature pertaining to the safety of organic foods also concluded that bacterial contamination of retail chicken is common but unrelated to organic status (Smith-Spangler et al., 2012). However, they did find evidence that the risk of isolating bacteria resistant to three or more antibiotics was higher in conventional than in organic chicken, although they did not consider specific resistance genotypes (Smith-Spangler et al., 2012). In addition, they did not include antibiotic-free products in their review. Our results provide little evidence of differences in the microbiological quality of fresh retail chicken labeled organic, antibiotic free, or conventional without either label claim.
We recovered Campylobacter spp. from 11% of the packages, and we could not detect a difference in recovery between product types. Our Campylobacter recovery rate was lower than expected based on previous reports from fresh retail chicken products (Williams and Oyarzabal, 2012; Trimble et al., 2013). We believe that our recovery of Campylobacter spp. may have been reduced by our practice of transporting the fresh packages of chicken breasts at ambient temperature from the stores to the laboratory during summer months. Temperature has been shown to impact the survival of Campylobacter in fresh meat products (Park, 2002). This type of nondifferential misclassification of Campylobacter contamination status is generally expected to introduce bias toward the null hypothesis of no difference between product types (Chen et al., 2013). However, it has been previously reported that recovery of Campylobacter is similar between conventional and antibiotic-free chicken products (Price et al., 2005).
The AmpC bla CMY-2 was commonly present in E. coli and Salmonella recovered from boneless chicken breast packages. bla CMY-2 has been previously reported in fresh meat products including beef, pork, and chicken (Mollenkopf et al., 2011; White et al., 2001; Dutil et al., 2010; Chen et al., 2004; Folster et al., 2012). In addition, bla CMY-2 has been reported to be widely disseminated in food animal populations (Tragesser et al., 2006; Heider et al., 2009; Mollenkopf et al., 2012). The extended-spectrum β-lactamase bla CTX-M has only recently been detected in E. coli and Salmonella from livestock in the United States (Wittum et al., 2010, 2012; Mollenkopf et al., 2012). In addition, an E. coli isolate carrying bla CTX-M was recovered from retail chicken in Pennsylvania that was identical to a human clinical isolate (Doi et al., 2010). Extended-spectrum cephalosporins are not approved for use in broiler production in the United States, although ceftiofur sodium is approved for use in day-old chicks for the control of early mortality associated with E. coli. However, the frequency of ceftiofur use in chickens has not been reported.
We recovered clonal or highly similar Salmonella strains from packages representing different grocery store chains and different processing plants. Because broiler chickens are raised in grower/finisher barns using all in/all out production systems, our result suggests that these common Salmonella strains must originate from a common source earlier in the production system such as a common hatchery or common parent stock. Hatcheries have previously been associated with the widespread dissemination of Salmonella in chicks (Gaffga et al., 2012; Wilkins et al., 2002). However, the role of broiler chicken parent stock in the maintenance and dissemination of Salmonella strains has not been reported. We also recovered highly similar C. jejuni and C. coli from diverse sources, suggesting a common source prior to broiler barns. Field research to identify common sources for Salmonella and Campylobacter spp. dissemination and to develop effective interventions might result in reduced zoonotic foodborne transmission of these pathogens.
We observed considerable variability in the contamination rates of retail meat products associated with the specific processing plant from which the product originated, which resulted in highly variable contamination rates among individual stores and between store chains. The large observed differences between processing plants may be due to differences in live bird infection rates, or in management and process control factors in place at individual plants resulting in different contamination rates. This result suggests that retail stores can directly influence the contamination rate of fresh chicken products by modifying their purchasing practices to take differences in plant contamination rates into account. Unfortunately, this information may not be readily available to individuals making purchasing decisions for retail grocery stores. All poultry processing plants in the United States must monitor Salmonella contamination rates of their fresh meat products as part of required Hazard Analysis Critical Control Point surveillance programs (FDA, 2014). However, this information is not available in a format that is accessible to consumers or that can be used by retailers to inform their purchasing decisions.
Conclusions
Retail meat products labeled “organic” or “antibiotic free” are marketed with an implied health benefit, and consumers of these products may reasonably expect that they be less likely to contain pathogens or antimicrobial-resistant organisms. Our results indicate that, regardless of product type, fresh retail chicken breast is commonly contaminated with enteric pathogens associated with foodborne illness and commensal bacteria harboring genes conferring resistance to critically important antimicrobial drugs. Consumers, regardless of product type, will need to practice appropriate kitchen hygiene in order to reduce their risk of foodborne illness.
Footnotes
Acknowledgments
The authors gratefully acknowledge the technical support of Joshua B. Daniels and Kehinde Olugbenga of The Ohio State University. This work was supported by the USDA NIFA award no. 2010-65201-20598.
Disclosure Statement
No competing financial interests exist.