
Abstract
This study characterized 52 Escherichia coli isolates from distinct diseased organs of 29 broiler chickens with clinical symptoms of colibacillosis in the Southern Brazilian state of Rio Grande do Sul. Thirty-eight isolates were highly virulent and 14 were virtually avirulent in 1-day-old chicks, yet all isolates harbored virulence factors characteristic of avian pathogenic E. coli (APEC), including those related to adhesion, iron acquisition, and serum resistance. E. coli reference collection phylogenetic typing showed that isolates belonged mostly to group D (39%), followed by group A (29%), group B1 (17%), and group B2 (15%). Phylogenetic analyses using the Amplified Ribosomal DNA Restriction Analysis and pulse-field gel electrophoresis methods were used to discriminate among isolates displaying the same serotype, revealing that five birds were infected with two distinct APEC strains. Among the 52 avian isolates, 2 were members of the pandemic E. coli O25:H4-B2-ST131 clone.
Introduction
A
Use of antimicrobials to prevent or treat APEC infections has allowed a great rise in the productivity of the poultry industry. It has also favored the selection of resistant E. coli isolates, posing concerns to the poultry industry itself and also to human health, because chicken products could be a source of foodborne extraintestinal pathogenic E. coli (Johnson et al., 2012; Mellata, 2013).
To be classified as APEC, an E. coli must have been isolated from lesions of a bird with signs of colibacillosis and should harbor a set of APEC-associated virulence genes (VAGs), encoding adhesins, iron-scavenging systems, protectins, and other traits (Brito et al., 2003; Ewers et al., 2007) that can be present in a wide variety of combinations. Moreover, APEC isolates fall mainly into serogroups O78 and O2, followed by O1 (Rodriguez-Siek et al., 2005b; Ewers et al., 2007; Dziva and Stevens, 2008; Ozawa et al., 2008; Schouler et al., 2012). Still, since the APEC pathotype is not yet fully defined and less pathogenic E. coli can be isolated from ill birds, the application of Koch's postulates and in vivo assays are necessary to verify the virulence of a particular E. coli isolated from a chicken (Schouler et al., 2012; Barbieri et al., 2013).
Brazil is the largest exporter and the third largest producer of poultry meat in the world (UBABEF, 2013), and its Southern states account for ∼75% of the total Brazilian production. Thus, infections by APEC represent significant losses to the Brazilian economy. In this work we characterized 52 E. coli isolates from 29 broiler chickens with clinical symptoms of colibacillosis, which, in some cases, included E. coli isolated from different organs of the same bird. All were isolated from birds in the Southern Brazilian state of Rio Grande do Sul. The antimicrobial susceptibility, the prevalence of virulence-associated factors, and virulence in 1-day-old chickens were also assessed. Serotyping and phylogenetic analysis allowed us to ascertain whether a diseased bird could be infected with more than one distinct pathogenic strain.
Materials and Methods
Bacterial strains
Fifty-two E. coli isolates were obtained from colisepticemic lesions from 29, 7-week-old broiler chickens, at the time of their slaughter, during the sampling periods September 2007 to January 2008, and January 2010 to May 2010. Lesions from different organs, including the heart, liver, air sacs, skin, and intestine, were sampled in the same birds.
The sampled poultry represented flocks from 14 distinct farms from various locations within the Centre-East and Centre-South regions of Rio Grande do Sul (RS), Brazil. Biochemical tests (triple sugar iron, urease, and MacConkey) were performed to confirm that all isolates were E. coli, and all strains were stored at −80°C in Luria-Bertani broth with 20% glycerol until use.
Antimicrobial resistance in APEC
The antimicrobial susceptibility of all APEC isolates was examined using the disc diffusion test according to the Clinical and Laboratory Standards Institute (CLSI) guidelines, using E. coli strain ATCC 25922 as a control (Barbieri et al., 2013; CLSI, 2011). The antimicrobial agents tested included the following: ampicillin (10 μg), cephalothin (30 μg), ceftiofur (30 μg), ciprofloxacin (5 μg), chloramphenicol (30 μg), enrofloxacin (5 μg), gentamicin (10 μg), neomycin (30 μg), nitrofurantoin (300 μg), norfloxacin (10 μg), tetracycline (30 μg), sulphonamides (300 μg), trimethoprim (5 μg), and a combination of sulphonamide and trimethoprim (23.7 μg plus 1.3 μg). All antimicrobial discs were from CEFAR (São Paulo, Brazil). The breakpoints were obtained from CLSI (CLSI, 2011) for all antimicrobials, except for ceftiofur (values for bovine mastitis) and enrofloxacin (CLSI, 2013), and neomycin (Sayah et al., 2005).
DNA extraction and multiplex polymerase chain reactions (PCRs)
Bacterial DNA was obtained from whole organisms by boiling (Barbieri et al., 2012). The extracts were stored at 4°C, and the supernatants were used as templates for gene amplification.
The presence of 34 VAGs in the isolates was investigated using multiplex PCRs as described (Barbieri et al., 2012).
Serotyping
The identification of APEC serotypes was carried out following standard methods, using O (O1–O181) and H (H1–H56) antisera; O/H antigens were determined by a tube agglutination test (Ewing, 1986).
PCR detection of O25b-ST131 clone isolates
The multilocus sequence type of O25:H4 strains RS004 and RS005 were assessed using the PCR method described by Clermont et al. (Clermont et al., 2009). Reactions were performed in a GenePro Thermal Cycler (Bioer Technology, China).
PCR-based classification into “ECOR” phylogenetic groups
Isolates were classified into four different Escherichia coli reference collection (ECOR) phylogenetic groups (A, B1, B2, and D) based on the presence and absence of chuA, yhaA genes and tspE4.C2 fragment (Clermont et al., 2000). Reactions were performed in a GenePro Thermal Cycler (Bioer Technology, China) as follows: denaturation for 4 min at 94°C, 30 cycles of 5 s at 94°C and 10 s at 59°C, and a final extension step of 5 min at 72°C.
ARDRA phylogenetic analysis
Genetic data was obtained using the Amplified Ribosomal DNA Restriction Analysis (ARDRA) method as previously described (Barbieri et al., 2013).
Pulsed-field gel electrophoresis (PFGE) analysis
All strains were subjected to molecular subtyping using PFGE. Isolates were analyzed using the method of Ribot et al. (Ribot et al., 2006). Preparation, lysis, washing of plugs, and XbaI restriction were performed according to the PulseNet protocol. Salmonella Braenderup H9812 was used as a DNA marker. Macrorestriction patterns were compared using the BioNumerics Fingerprinting software (Ver 6.6, Applied Math, Austin, TX). The similarity index was calculated using the Dice coefficient, with a band position tolerance of 1% and an optimization of 0.5%.
Lethality and pathogenicity tests
Lethality and pathogenicity tests were performed as described (Barbieri et al., 2012, 2013). The lethality score (LS) represents the number of animals that died within 7 days. The pathogenicity score (PS) combines the times of death and clinical scores observed at necropsy; the PS for each strain represents the median PS for the 10 chicks infected with that particular strain.
Ethics statement
All animal experiments were approved by the Biosafety Committee of the Instituto de Pesquisas Veterinárias Desidério Finamor (CIB 004/08), and chickens were euthanized according to animal welfare norms. All the procedures were done in the presence of a qualified veterinarian.
Statistical analysis
PS and LS, resistance, and number of VAGs were treated as quantitative variables and described by mean±SD. Data were analyzed using nonparametric tests due to asymmetry in their distributions, except for number of VAGs. The relationship between the presence of a gene and the PS was analyzed using the Wilcoxon–Mann–Whitney test. All statistical analyses were carried out with the Statistical Package for the Social Sciences (SPSS v.18.0, IBM). Statistical significance was accepted at p≤0.05.
Results
Antimicrobial resistance among the APEC isolates
We tested 52 APEC isolates for susceptibility to 14 antimicrobial agents that are or had been commonly employed in the poultry industry in Brazil to prevent/treat infections and promote growth and performance. We observed that APEC isolates presented frequencies of resistance <25% for 9 antimicrobials with exceptions being those antimicrobials that act on the cell wall (ampicillin, with 75% resistance, cephalothin, 65%, and ceftiofur, 56%); and antimicrobials that have been on the market for a long time, such as tetracycline (69.4% resistance) and sulphonamides (59.7%) (Fig. 1).
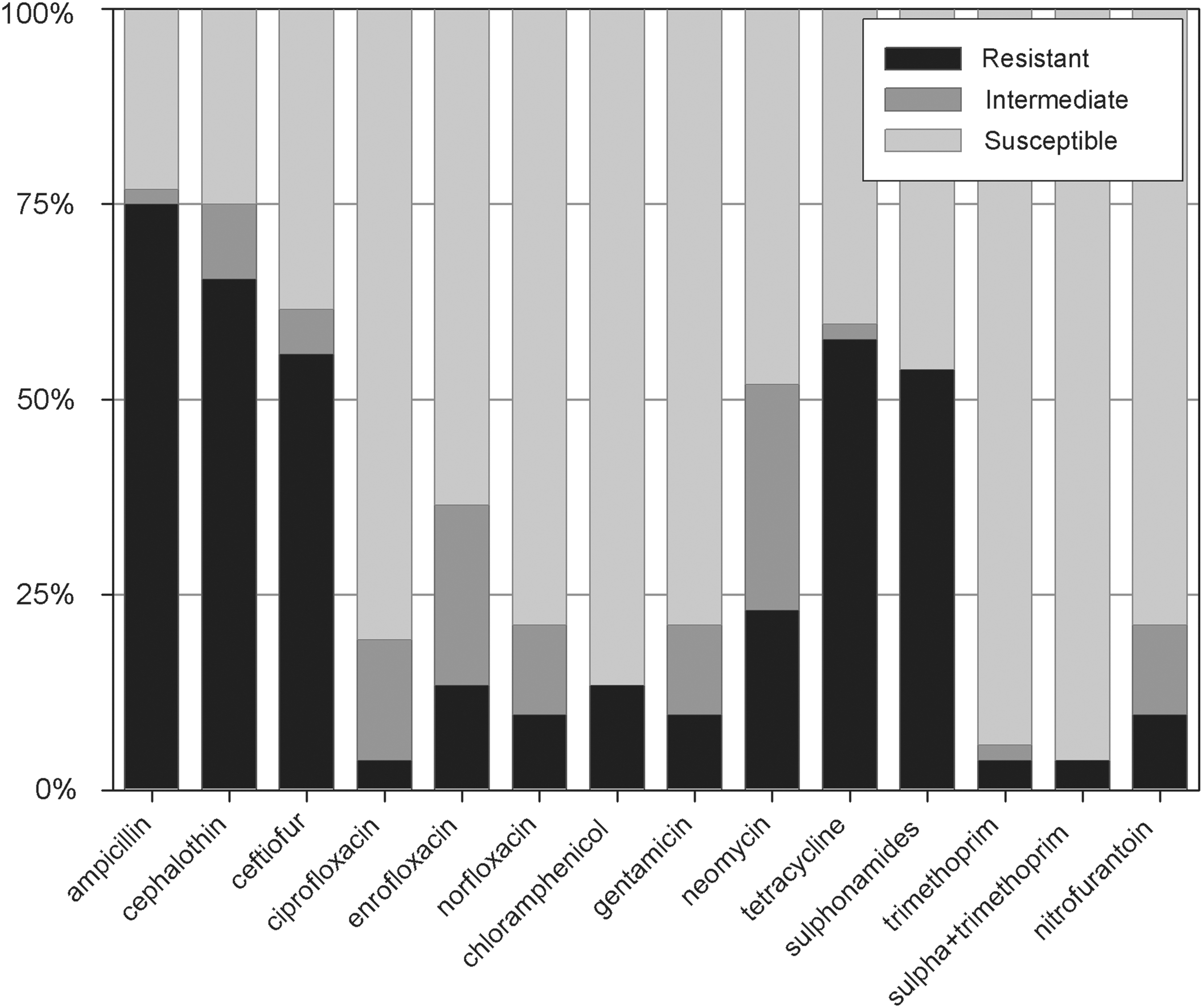
Antimicrobial susceptibility of colisepticemic isolates. The susceptibility of 52 avian Escherichia coli isolates from colisepticemic chickens to 14 antimicrobials was tested individually using disc diffusion tests.
Considering the site of antibiotic action, 77% of the APEC isolates were resistant to at least one of the aforementioned antimicrobials that act on the cell wall; 15% were resistant to at least one of the antimicrobials that inhibit nucleic acid synthesis (ciprofloxacin, enrofloxacin, norfloxacin); 65% were resistant to at least 1 of the antimicrobials that inhibit protein synthesis (chloramphenicol, gentamicin, neomycin, tetracycline; but only 33% if tetracycline is excluded); and 56% were resistant to at least 1 of the antimicrobials that are antagonistic to metabolic activity (sulphonamides, trimethoprim, sulpha+trimethoprim; but only 6% if sulphonamides are excluded).
Concerning the multiresistance pattern, 23% of APEC isolates were resistant to at least 1 class of antimicrobial agents; 25% of APEC isolates were resistant to 2; 37%, to 3; and 8%, to all 4 classes. It is noteworthy that there was no antimicrobial agent to which all 52 APEC strains were susceptible.
Genotyping by multiplex PCR
Prevalence of VAGs among APEC isolates is shown in Figure 2 (shaded area). The genes fimC, csgA, and crlA were present in all isolates, and afa/dra was not detected in any isolate. Figure 3 specifies the profile of VAGs of each isolate. Overall, the isolates had an average of 14.2 VAGs (range from 9 to 23 VAGs).
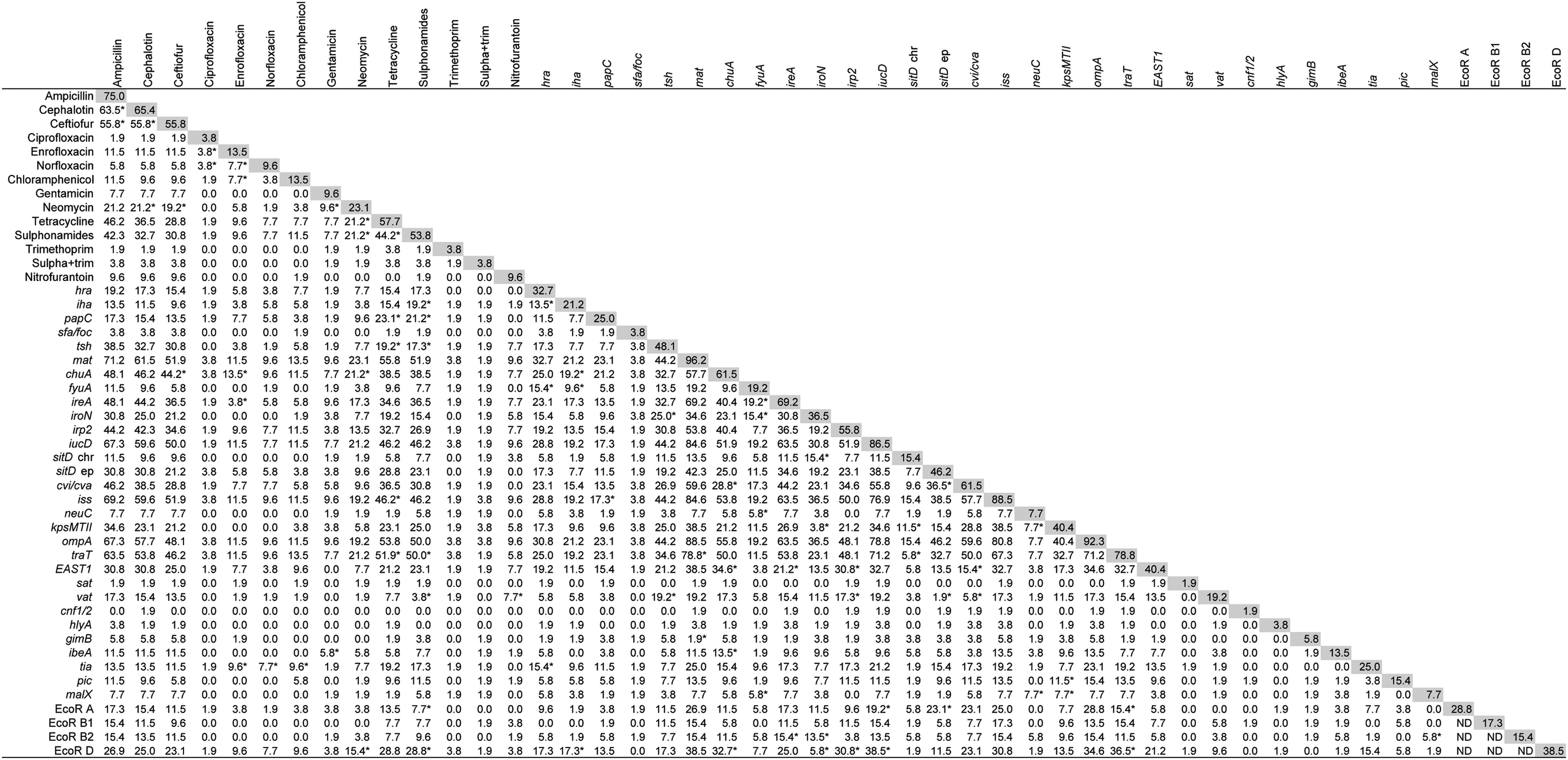
Association between presence of 30 avian pathogenic Escherichia coli (APEC)–associated virulence genes (VAGs) among 52 APEC isolates. Numbers indicate the percentage of isolates that have both traits, while numbers in shaded boxes indicate the percentage of isolates that have the corresponding trait; *p≤0.005 using chi-square test. ND, not determined. afa was not included, since it was absent in all strains, and fimC, crlA, and csgA were not included, since they were present in all strains.
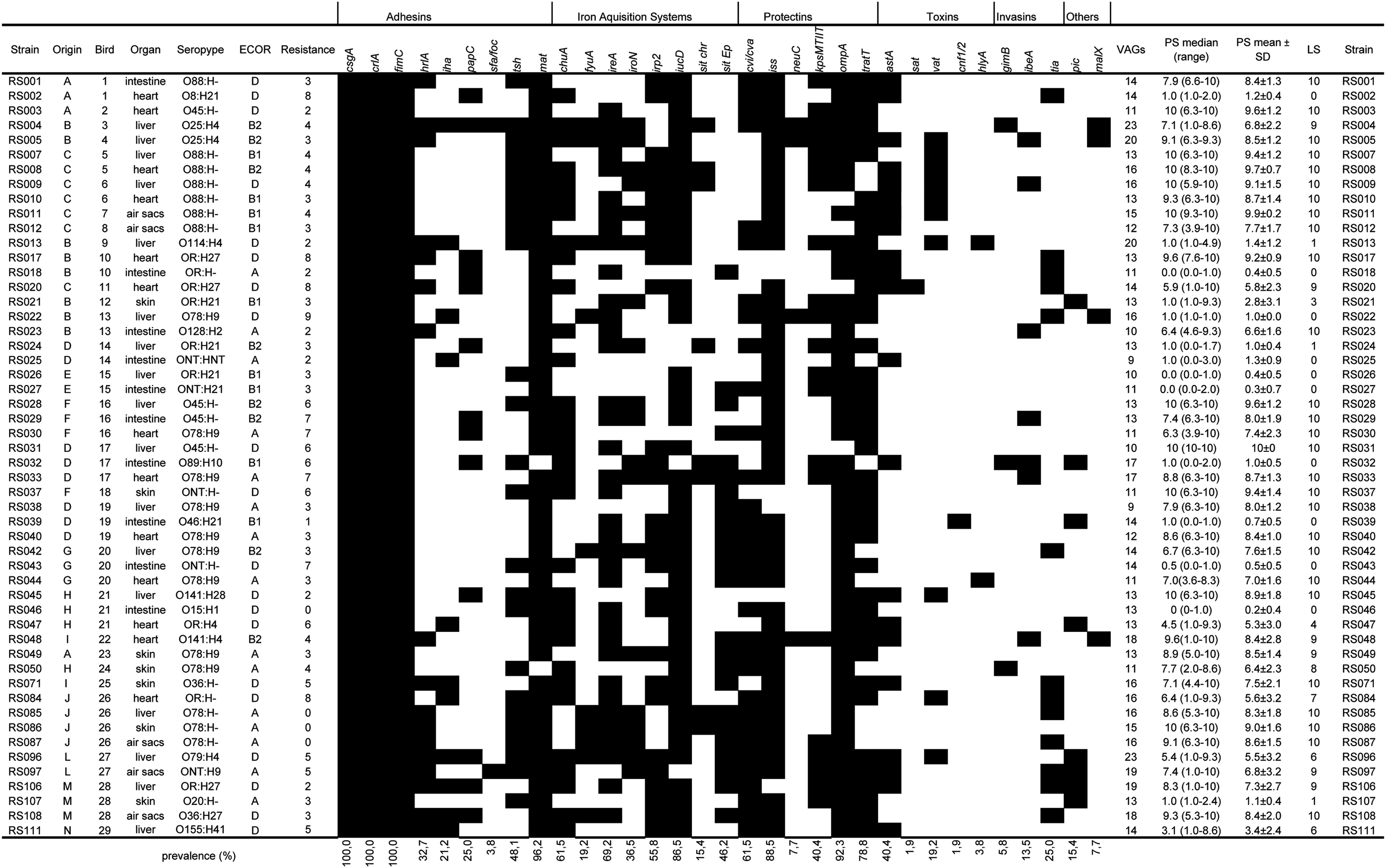
Characterization of 52 avian pathogenic Escherichia coli (APEC) isolates. Columns from left to right: Strain, isolate designation; Origin, source of the isolate, with A to N designating each of the 14 farms from which the isolates were collected; Bird, same numbers designate the same animal; Organ, organ of bacteria isolation; Serotype; ECOR, ECOR phylogenetic group; Resistance, number of antimicrobials to which the isolate was resistant to (out of 14 tested); the subsequent columns depict the polymerase chain reaction results for all APEC-associated virulence genes (VAGs) tested, with presence indicated in black and absence indicated in white; no. VAGs, total number of VAGs in each isolate; PS median, median pathogenicity score (range); PS mean±SD, mean pathogenicity score ± standard deviation; LS, lethality score.
Virulence factors related to adhesion, iron acquisition, and serum resistance were present in all strains. ColV plasmid-associated genes (cvi/cva, iroN, iss, iucD, sitD, traT, tsh) occurred in the majority (from 88% to 36%) of isolates, and cvi/cva itself was present in 61% of the isolates (Fig. 3). The isolates harbor an average of 4.8 genes associated with adhesion (range from 3 to 8 out of 9 tested), 3.9 associated with iron acquisition systems (range from 1 to 7 out of 8 tested), 3.7 associated with serum resistance (range from 3 to 6 out of 6 tested), 0.7 associated with toxins (range from 0 to 2 out of 5 tested), and 0.4 genes associated with invasins (range from 0 to 2 out of 3 tested).
Serotyping
Among 52 serotyped APEC strains, 39 (75%) could be classified into a single O-type, 5 isolates were nontypeable with the antisera used, and 8 isolates displayed a rough phenotype (Fig. 3). Most of the typeable strains belonged to O-types O78 (12; 23%), O88 (7; 13%), and O45 (4; 8%), while 16 (31%) were distributed among 13 different O-types. Considering the H antigen, 31 (60%) APEC isolates could be classified into a single H-type. Isolates belonged to H-types H9 (10; 19%), H4 (6; 12%), H21 (6; 12%), and H27 (4; 8%). Only 1 strain was nontypeable with any of H-anti-serum used, whereas 20 isolates were nonmotile.
PCR detection of O25b-ST131 clone isolates
O25:H4-B2 strains RS004 and RS005 belong to multilocus sequence type 131, as verified by amplification of the pabB gene (Clermont et al., 2009) (results not shown). Both strains were resistant to ampicillin, cephalothin, and ceftiofur, and strain RS004 was also resistant to sulphonamides.
Lethality and pathogenicity tests
We assessed the virulence of the APEC isolates in day-old chickens, and results for each strain are presented in Figure 3. Twenty eight isolates (54%) killed 10 chickens within 7 days (LS=10), and 10 (19%) isolates did not kill any chickens. Among these 10 isolates with LS=0, 7 had been isolated from intestinal lesions (RS18, RS25, RS27, RS32, RS39, RS43, and RS46); thus, they likely represent nonpathogenic, commensal E. coli. Overall, the APEC isolates had a mean LS of 6.9.
For the PS, which also considers the clinical signs and how quickly the infection kills birds, 10 (19%) isolates had a median PS of 10 (i.e., most chicks died in the first day postinfection), 20 (38%) had a 9.9 >PS >7.0, 8 (15%) had a 6.9 >PS >3.0, and 14 (27%) had a median PS <3 (which include the 10 isolates with LS=0, Fig. 3). Overall, the APEC isolates had a mean PS of 6.1.
When we correlated the presence of VAGs to PS (Table 1), we found that a higher PS was significantly associated (p≤0.05) with cvi/cva and vat, but negatively associated (p≤0.05) with pic.
Genes that occurred in none (afa/dra), only one (cnf1/2, sat), or all (fimC, crlA, csgA) isolates are not listed.
The relationship between the presence of a gene and the pathogenicity score was analyzed using the Wilcoxon–Mann–Whitney test, by comparing the scores in isolates with and without this particular gene. Exact p values.
p≤0.05.
ECOR grouping
The distribution of APEC isolates among the four phylogenetic groups is shown in Table 2. Most strains (38%) belonged to group D.
Mean number of antimicrobials to which the isolates were resistant, excluding tetracycline and sulphonamides.
Table 2 also shows the mean number of antimicrobials to which strains were resistant, mean number of VAGs, PSs, and LSs of strains according to different ECOR groups. It was observed that, although not statistically significant, isolates from group B2 had a higher number of VAGs per strain and higher PS and LS, whereas isolates from group B1 had lower PS and LS.
Phylogenetic analysis
The ARDRA analysis revealed 49 different profiles among the 52 isolates examined (Fig. 4). Three pairs of isolates (RS007 and RS008, RS004 and RS005, and RS021 and RS022) had identical restriction patterns. The isolates separated into 2 major clusters at a similarity level of 70% or more (Fig. 4), with the majority of isolates belonging to cluster B. It was also found that cluster B isolates had a higher number of VAGs (average of 15.1) and a higher PS (average of 6.5) when compared to cluster A (average of 13.3 VAGs and an average PS of 5.8).
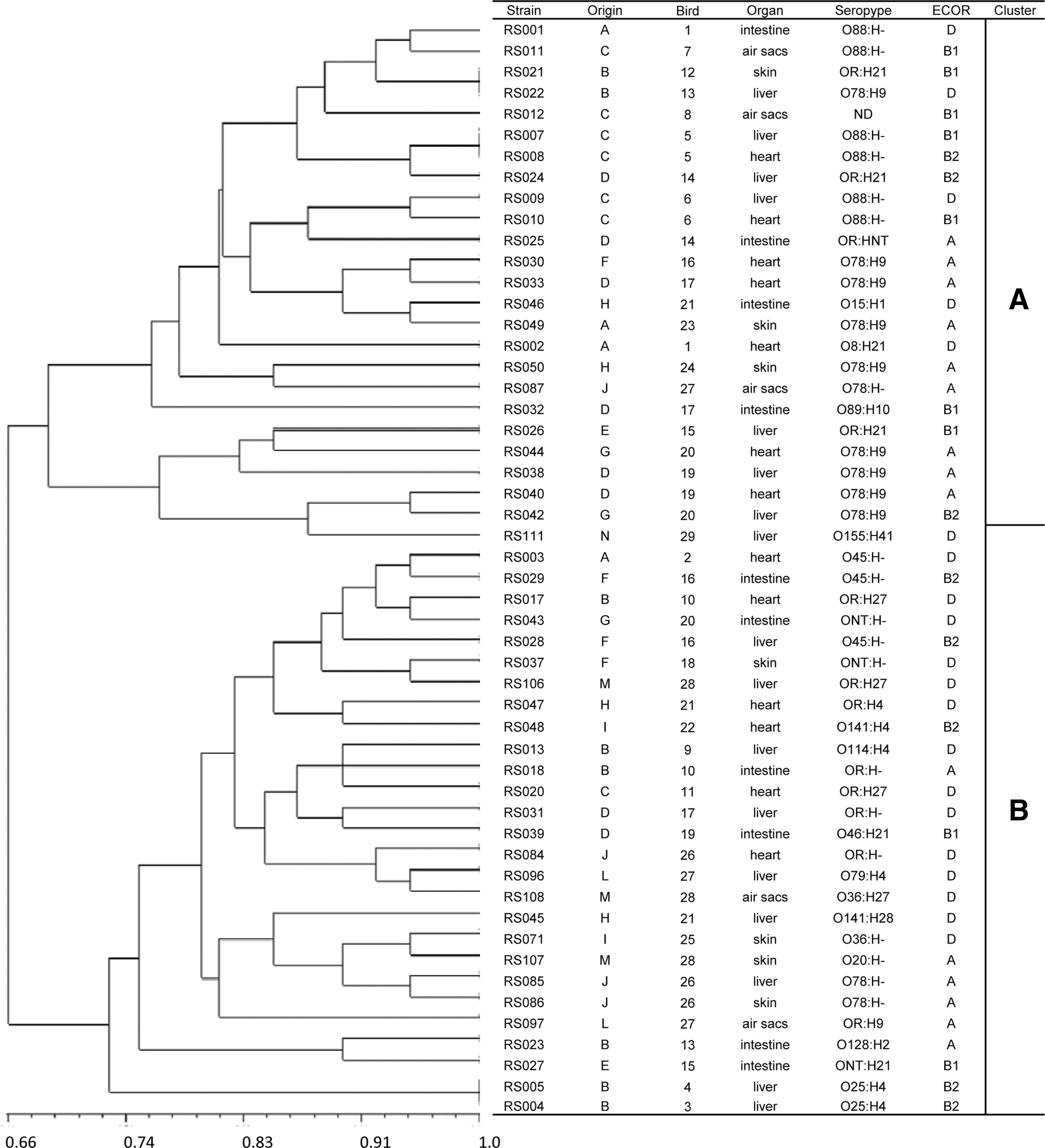
Amplified Ribosomal DNA Restriction Analysis (ARDRA) profile of 52 avian pathogenic Escherichia coli isolates. The ARDRA dendrogram was constructed by unweighted-pair group method with arithmetic mean based upon enzyme restriction digestion of amplified 16-23S DNA intergenic spacer regions. The column Strain shows isolate designation; the column Origin, source of isolate, with A to N designating each of the 14 farms from which the isolates were collected; Bird, same numbers designate the same animal; Organ, organ of bacteria isolation; Serotype; ECOR, ECOR phylogenetic group; Cluster designates the 2 genotypic clusters (A and B) into which strains with 70% similarity were grouped. E. coli ATCC25922 was analyzed as a reference strain.
The ARDRA analysis also revealed that, among the 15 birds from which 2 to 4 isolates were collected, 5 birds were infected with 2 distinct APEC isolates (Fig. 4), namely, birds 16, 17, 21, 26, and 27 (column “bird” in Fig. 3).
To confirm that some birds were indeed infected with phylogenetically distinct APEC strains, we performed phylogenetic analysis using PFGE. The results with the PFGE method were essentially the same as those with ARDRA, confirming that the five aforementioned birds were infected with two distinct APEC strains (Fig. 5). Moreover, 42 different PFGE profiles were found among the 48 isolates (Fig. 5). From the dendrogram generated, 3 major clusters could be grouped at a similarity level of 60% or more, with the majority of isolates belonging to cluster B. It was also found that cluster B isolates had a higher PS (average of 6.4) and a higher LS (average of 7.7) when compared to clusters A and C.

Pulse-field gel electrophoresis (PFGE) profile of 52 APEC isolates. The PFGE dendrogram was constructed by the unweighted-pair group method with arithmetic averages. The scale indicates levels of similarity within this set of isolates based upon XbaI enzyme restriction digestion of total bacterial DNA. The column Strain shows isolate designation; the column Origin, source of isolate, with A to N designating each of the 14 farms from which the isolates were collected; Bird, same numbers designate the same animal; Organ, organ of bacteria isolation; Serotype; ECOR, ECOR phylogenetic group; Cluster designates the 3 genotypic clusters (A to C) into which strains with 60% similarity were grouped. Escherichia coli ATCC25922 was analyzed as a reference strain. The XbaI could not digest the strains RS026, RS027, RS028, and RS043.
Discussion
The purpose of this study was to characterize avian E. coli isolates from diseased organs of 29 colisepticemic chickens. From 14 chickens, we collected 1 isolate per bird, and from 15 chickens we collected from 2 to 4 isolates from distinct organs, totaling 52 isolates. We found that a diseased bird can be infected with two distinct pathogenic strains.
The genotyping of our collection of colisepticemic E. coli showed that all 52 isolates carry genes encoding adhesins, iron-scavenging systems, and protectins, thought to enable APEC to resist host defenses. The fact that APECs possess several genes for adhesins, iron-scavenging systems, and protectins underscores the importance of these activities in the successful establishment of extraintestinal infections. On the other hand, 14 isolates were avirulent to 1-day-old chickens (PS <1) but were indistinguishable from the virulent ones in terms of APEC VAGs, once more showing that mere carriage of APEC VAGs does not necessarily mean that an E. coli will cause colibacillosis (Ewers et al., 2009). Of these 14 avirulent strains, 7 had been isolated from intestinal lesions and are likely commensal E. coli that nevertheless carry APEC genes (Kemmett et al., 2013). Three isolates from intestinal lesions, in contrast, were highly virulent to 1-day-old chickens (RS001, RS023, and RS029, Fig. 3) and are probably APEC.
We could not isolate virulent strains from birds 9 (RS013), 12 (RS021), 13 (RS022), 14 (RS024 and RS025), and 15 (RS026 and RS027), so the disease may be a result of causes other than APEC infection (Barbieri et al., 2012).
The presence of genes encoding colicin and Vat in E. coli from colisepticemia was correlated with higher PS (Table 1). This is in agreement with the observation that large conjugative ColV plasmids and the associated genes for colicin (cvi/cva) are well known virulence markers of APEC (Rodriguez-Siek et al., 2005a, 2005b; Skyberg et al., 2008). Vat is a serine protease autotransporter whose gene is located on a pathogenicity island in extraintestinal E. coli and has been associated with virulence in APEC (Parreira and Gyles, 2003; Dziva et al., 2013). In contrast, the presence of another serine protease autotransporter, pic, was associated with a lower PS, in agreement with what we have reported previously for cellulitis isolates (Barbieri et al., 2013). Although pic had been associated with virulence in uropathogenic E. coli (Heimer et al., 2004), our data suggest that it is not associated with virulence in APEC.
To verify how closely related the isolates were, we performed phylogenetic analyses using ARDRA and PFGE. Both methods showed that 5 chickens were infected with genetically distant (<60% PFGE similarity) APEC isolates (Fig. 4 and Fig. 5). The serotypes confirmed that these isolates are distinct. To our knowledge, this is the first report showing that a colisepticemic chicken was infected with more than one APEC strain.
When we further analyzed the clonal relationships, we found that closely related strains (>85% PFGE identity) displayed the same serotype, and this result is more evident in the PFGE (Fig. 5) than in the ARDRA method (Fig. 4). Dziva and colleagues have previously observed that APEC O78 strains χ7122 and IMT2125 were more closely related to ETEC O78 than to APEC O1 (Dziva et al., 2013). It is worth mentioning that phylogenetically similar strains with the same serotype can belong to distinct ECOR groups (Fig. 5).
Type O78 was the most prevalent O serogroup among our strains, in agreement with data from the United States (Rodriguez-Siek et al., 2005b), France (Schouler et al., 2012), Germany (Ewers et al., 2007), Japan (Ozawa et al., 2008), and China (Chen et al., 2012), followed by O88. However, we did not find any strain belonging to serogroup O1 or O2. The serotyping revealed that isolates RS004 and RS005 belong to serotype O25:H4. We also found 10 O25:H4 isolates (unpublished results) among our collection of 144 APEC cellulitis isolates (Barbieri et al., 2013). Both RS004 and RS005 are ECOR B2, carry genes for capsule, and were positive for ST131. To our knowledge, this is the first time members of E. coli O25:H4-B2-ST131 clone have been detected among avian isolates in Brazil. This is a reason for concern, because this clone frequently carries genes encoding extended-spectrum β-lactamases, which are emergent bacteria causing disease among humans, livestock, and companion animals worldwide (Ewers et al., 2010; Mora et al., 2010; Ahmed et al., 2013).
Our strains displayed low levels of resistance for most antimicrobials, but exhibited high levels of resistance to tetracycline and sulphonamides and for antimicrobials that act on the cell wall (ampicillin, cephalothin, and ceftiofur). Tetracycline has been used for a long time to promote the growth of poultry, and high levels of resistance to tetracycline and sulphonamides (≥75%) have been reported many times previously (Yang et al., 2004; Zhao et al., 2005; Ahmed et al., 2013). However, antimicrobial use for growth promotion and prevention has been banned in the Brazilian poultry industry since 2009 (MAPA, 2009); thus, studies should be conducted in the near future to determine whether this ban is having the desired effect in reducing antimicrobial resistant infections. With regard to antimicrobials that act on the cell wall, although high levels of resistance have been reported for ampicillin (Yang et al., 2004; Ozawa et al., 2008), we observed a higher resistance levels to cephalothin and ceftiofur in comparison to our previous and other works (Zhao et al., 2005; Johnson et al., 2012; Barbieri et al., 2013).
Conclusions
By characterizing and using in vivo virulence studies of a collection of 52 E. coli isolates from colisepticemic chickens, we have shown that 1 chicken can be infected with more than 1 pathogenic strain simultaneously. We also report for the first time the existence of members of E. coli O25:H4-B2-ST131 clone among avian E. coli isolates in Brazil.
Footnotes
Acknowledgments
This work was partially supported by the CNPq (MAPA 578364/2008-5), Brazil. N.L.B. was the recipient of a CNPq PhD student (process number 202479/2011-0), and T.M.T. was the recipient of a CNPq studentship (RHAE 573367/2008-6).
Disclosure Statement
No competing financial interests exist.