
Abstract
Foodborne pathogenic bacteria can live in the intestinal tract of food animals and can be transmitted to humans via food or indirectly through animal or fecal contact. Organic acid blend products have been used as nonantibiotic modifiers of the gastrointestinal fermentation of food animals to improve growth performance efficiency. However, the impact of these organic acid products on the microbial population, including foodborne pathogens, remains unknown. Therefore, this study was designed to examine the effects of a commercial organic acid and botanical blend product (OABP) on populations of the foodborne pathogenic bacteria, Escherichia coli O157:H7 and Salmonella Typhimurium. Pure cultures (2×106 colony-forming units [CFU]/mL) of each pathogen were added to tubes that contained water-solubilized OABP added at concentrations of 0, 0.1, 0.5, 1, 2, 5, and 10% (vol/vol; n=3). Water-solubilized OABP reduced (p<0.05) the growth rate and final populations of E. coli O157:H7 and Salmonella Typhimurium in pure culture at concentrations >2%. E. coli O157:H7 and Salmonella Typhimurium were added (2×105 and 3×106 CFU/mL, respectively) to in vitro mixed ruminal microorganism fermentations that contained water-solubilized OABP at concentrations of 0, 1, 2, 5, and 10% (vol/vol; n=3) that were incubated for 24 h. OABP addition reduced (p<0.05) final populations of E. coli O157:H7 and Salmonella Typhimurium in the ruminal fluid at concentrations ≥5%. The acetate–to-propionate ratios from the in vitro fermentations were reduced (p<0.05) by OABP treatment ≥5%. Treatments to reduce foodborne pathogens must be economically feasible to implement, and results indicate that organic acid products, such as OABP, can enhance animal growth efficiency and can be used to reduce populations of pathogenic bacteria.
Introduction
F
Typically, the commensal microbiome exists in an equilibrium that is beneficial to the host animal and excludes pathogenic bacteria (Duncan et al., 1999; Leatham et al., 2009) through a variety of mechanisms, including the antimicrobial effects of short-chain fatty acids (Wolin, 1969; Van Immerseel et al., 2006; Siragusa and Ricke, 2012). Inclusion of organic acids have been shown to improve feed efficiency and animal growth, and the present microencapsulated commercial organic acid and botanical blend product (OABP) has been used safely in animal feeds for >5 years in the European Union (Grilli et al., 2007; Piva et al., 2007b; EFSA, 2012). Short- and medium- chain fatty acids are directly active against Salmonella Typhimurium and Escherichia coli (Russell and Diez-Gonzalez, 1998), and have been shown to reduce pathogen colonization and shedding in swine (Boyen et al., 2008) and poultry (Van Immerseel et al., 2004b; Van Immerseel et al., 2006; Grilli et al., 2013). In some cases, the greatest antipathogenic effect has been observed in microencapsulated preparations that allow acids to reach targeted portions of the gastrointestinal tract and enhance growth most effectively (Piva et al., 2007a, b; Grilli et al., 2011, 2013). Therefore, the aim of this study was to determine whether an OABP could affect the growth of foodborne pathogens in pure culture and in in vitro mixed ruminal microorganism fermentations.
Materials and Methods
Bacterial strains and culture conditions
Escherichia coli O157:H7 strain 933 (ATCC 43895) and Salmonella Typhimurium used in this study were obtained from the Food and Feed Safety Research Unit (USDA/ARS, College Station, TX) culture collection. Both E. coli O157:H7 strain 933 and Salmonella Typhimurium were naturally resistant to 25 μg/mL novobiocin (NO) and were made resistant to 20 μg/mL of nalidixic acid (NA).
Pure culture studies
E. coli O157:H7 strain 933 or Salmonella Typhimurium cultures were anaerobically (90% N2, 5% H2, 5% CO2 atmosphere) incubated at 39°C in anoxic Tryptic Soy Broth (Difco Laboratories, Detroit, MI) for 24 h. OABP, a proprietary blend of citric acid, sorbic acid, thymol, and vanillin, was solubilized in sterile distilled water and was added to each tube to reach final concentrations of 0, 0.1, 0.5, 1, 2, 5, or 10% vol/vol. Growth rates of each tube (n=3) were estimated via measurement of absorbance changes using a Spectronic 20D spectrophotometer (600 nm) (Thermo Spectronic Inc., Madison, WI); maximum specific growth rate was calculated via the formula: (ln OD2 – ln OD1)/ΔT. Final optical densities (OD) after 24 h of incubation were measured by a Gilford 2600 spectrophotometer (600 nm, 1-cm cuvette). Cultures with OD >0.7 OD units were appropriately diluted in 0.9% NaCl.
Quantitative bacterial enumeration
Aliquots were taken from all in vitro fermentations at 6 and 24 h to determine the effect of OABP on populations of E. coli O157:H7 and Salmonella Typhimurium. Samples were serially diluted (in 10-fold increments) in phosphate-buffered saline (pH 7.0), and subsequently plated on MacConkey's agar (supplemented with 25 μg/mL NO and 20 μg/mL NA) and incubated at 37°C overnight for direct counting of E. coli O157:H7 CFU/mL. To determine populations of Salmonella Typhimurium, samples were serially diluted as described above and plated on Brilliant Green Agar (supplemented with 25 μg/mL NO and 20 μg/mL NA) and incubated at 37°C overnight for direct counting.
In vitro mixed ruminal microorganism fermentations
Cattle were maintained in accordance with a protocol approved by the Southern Plains Agricultural Research Center Animal Care and Use Committee (ACUC No. 06002). Cattle were provided ad libitum access to water and minerals in all studies. Ruminal contents were collected by hand from the ventral sac of ruminally cannulated Holstein cows (n=3) fed on pasture. The ruminal contents were collected from all cattle at approximately the same time (between 8:00 a.m. and 9:00 a.m.). Immediately after removal from the rumen, the contents from each cow were strained via a fine mesh nylon strainer (Reaves and Co., Durham, NC) and pooled. Ruminal fluid was transported to the laboratory and incubated for 30 min at 39°C to allow gas production to buoy large particles to the top of the flasks. Microscopic examination of diluted ruminal fluid revealed the presence of few feed particles or protozoa, and an abundance of bacteria with different morphological shapes.
Ruminal fluid fermentations were performed by combining the ruminal fluid (1:3) with an anoxic basal medium containing (per liter): 292 mg K2HPO4, 202 mg KH2PO4, 436 mg NH4SO4, 480 mg NaCl, 100 mg MgSO4·7H2O, 64 mg CaCl2·H2O, 4000 mg Na2CO3, and 600 mg cysteine hydrochloride (Cotta and Russell, 1982). Approximately 104 to 105 CFU/mL E. coli O157:H7 strain 933 and 103 to 104 CFU/mL Salmonella Typhimurium were added to the buffered ruminal fluid. The resultant suspensions were transferred to 18×150 mm Balch tubes (10 mL per tube) containing 0.2 g of ground hay (ground to pass a 1-mm Wiley Mill screen). OABP was added to each tube to reach final concentrations of 0, 0.1, 0.5, 1, 2, 5, or 10% vol/vol in triplicate (n=3) tubes. Tubes were then sealed using butyl rubber stoppers with aluminum crimps and incubated for 24 h at 39°C under a N2, CO2, H2 (90:5:5 vol/vol) gas phase. Samples were removed after 24 h of incubation, serially diluted, and plated as described above. Gas samples (1 mL) were removed after 24 h of incubation via a gas-tight syringe and analyzed for hydrogen (H2) and methane concentrations using a Gow Mac thermal conductivity series 580 gas chromatograph (Gow Mac Instruments, Bridgewater, NJ) equipped with a Haysep Q column (60°C, 20 mL/min argon carrier gas). Final pH in each incubation was determined immediately upon opening of the tube, and volatile fatty acids (VFA) concentrations were determined by HPLC (Corrier et al., 1994). Ammonia concentrations were determined using a colorimetric method (Chaney and Marbach, 1962).
Statistical analysis
Experiments were performed in triplicate (n=3), and the values presented are means. Student t-test was used to determine significance of differences between means.
Chemicals
Unless specifically mentioned, all chemicals were obtained from Sigma Chemical Company (St. Louis, MO).
Results
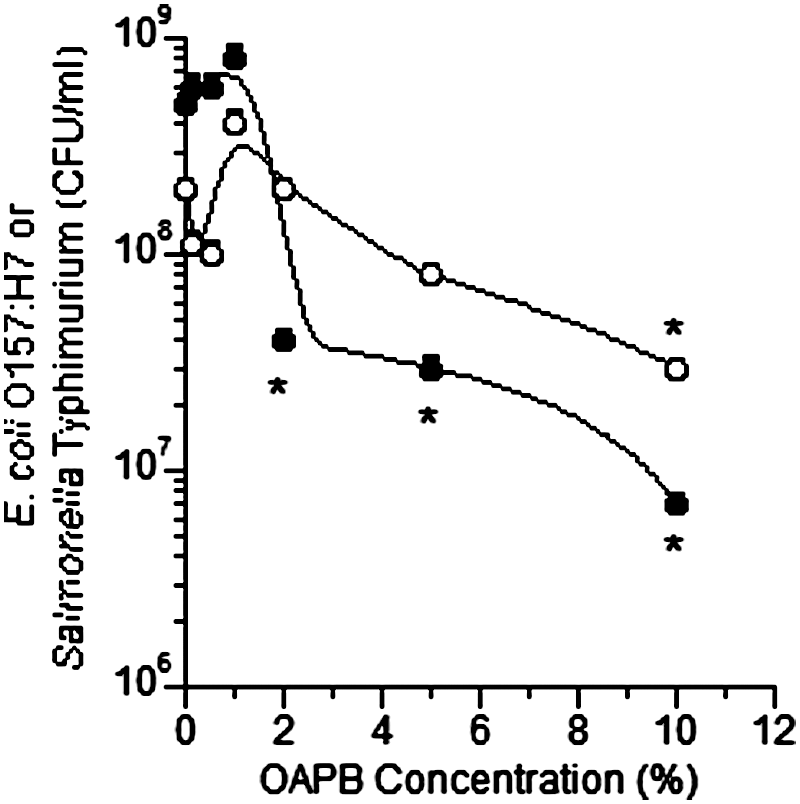
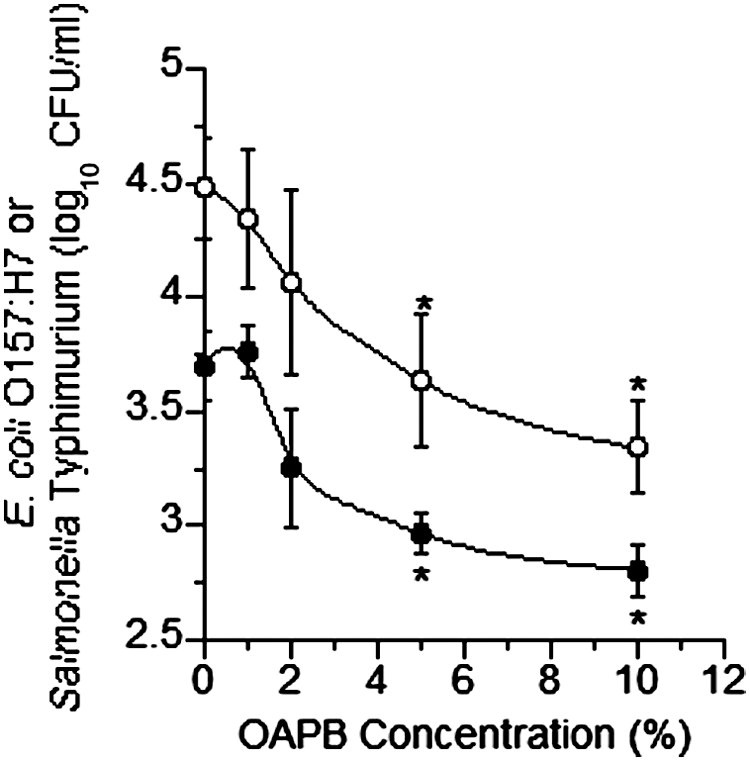
OABP addition to pure cultures at concentrations ≥2% reduced (p<0.05) the maximum specific growth rate of E. coli O157, while concentrations ≥1% reduced (p<0.05) the growth rate in Salmonella Typhimurium (Fig. 1). Populations (CFU/mL) after 24 h of growth of both E. coli O157:H7 and Salmonella Typhimurium were both reduced (p<0.05) by OABP concentrations of 10% and ≥2%, respectively (Fig. 2). Population differences at 6 h were similar to that at 24 h (data not shown). In mixed ruminal fluid microorganism fermentations containing inoculated populations of E. coli O157:H7 and Salmonella Typhimurium, OABP concentrations ≥5% reduced (p<0.05) pathogen populations (Fig. 3). The final pH was higher, but not statistically different in all concentrations of OABP; however, the acetate-to-propionate ratio (A:P) was reduced (p<0.05) in tubes containing OABP concentrations ≥5% (Fig. 4). Total VFA concentrations were not altered by OABP treatment, methane production and ammonia concentrations were lower in OABP-treated fermentations, but this difference was not significant (p>0.05).

Effect of increasing concentrations (vol/vol) of organic acid and botanical blend product (OABP) on the maximum specific growth rate (h−1) of Escherichia coli O157:H7 strain 933 (○) and Salmonella Typhimurium (●) grown in pure culture in tryptic soy broth at 39°C. Error bars indicate standard deviations; all cultures were grown in triplicate. Asterisks indicate differences from controls (p<0.05).
Effect of increasing concentrations (vol/vol) of organic acid and botanical blend product (OABP) on populations (colony-forming units [CFU]/mL) of Escherichia coli O157:H7 strain 933 (○) and Salmonella Typhimurium (●) grown in pure culture in tryptic soy broth for 24 h at 39°C. Error bars indicate standard deviations; all cultures were grown in triplicate. Asterisks indicate differences from controls (p<0.05).
Effect of increasing concentrations (vol/vol) of organic acid and botanical blend product (OABP) on populations (colony-forming units [CFU]/mL) of Escherichia coli O157:H7 strain 933 (○) and Salmonella Typhimurium (●) inoculated in mixed ruminal microorganism fermentations from cows fed a pasture diet. Error bars indicate standard deviations from triplicate fermentations and asterisks indicate differences from controls (p<0.05).
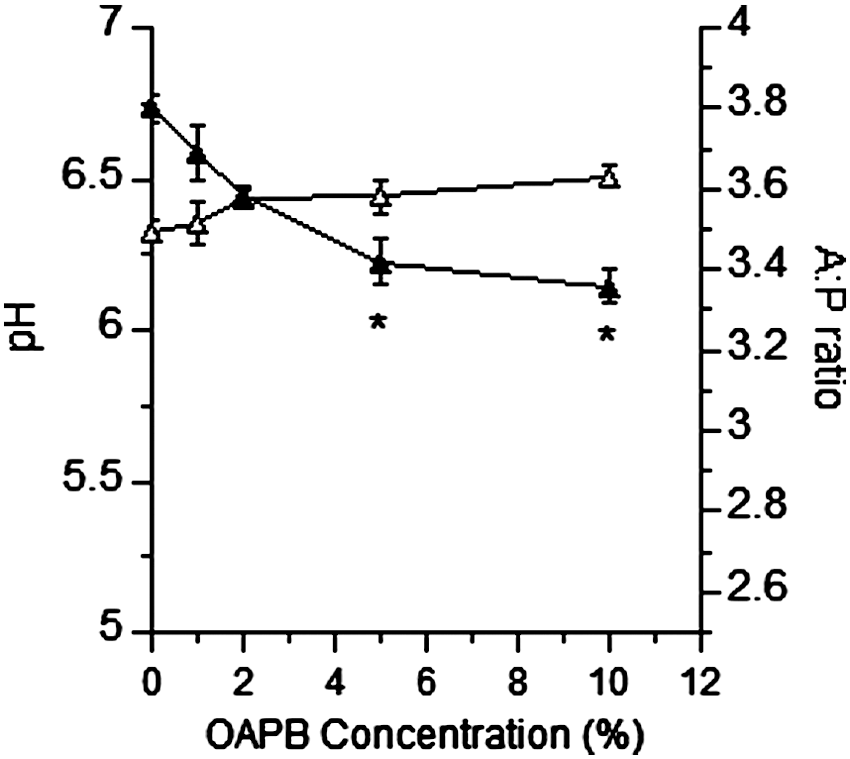
Effect of increasing concentrations (vol/vol) of organic acid and botanical blend product (OABP) on pH (△) and acetate:propionate ratio (A:P; ▲) in in vitro mixed ruminal microorganism fermentations from cows fed a pasture diet. Error bars indicate standard deviations from triplicate fermentations and asterisks indicate differences from controls (p<0.05).
Discussion
Foodborne pathogenic bacteria are a significant threat to human consumers and represent a significant drain on national gross domestic product. Salmonella is the second most common cause of human foodborne bacterial illness, causing at least 1.2 million human illnesses at an estimated cost of more than $14 billion a year in the United States (Scharff, 2010; Scallan et al., 2011). Shiga toxin–producing E. coli (STEC; such as E. coli O157:H7)–caused illnesses cost the American economy >$1 billion each year in direct and indirect costs from more than 175,000 human illnesses (Scharff, 2010; Scallan et al., 2011). Unfortunately, these pathogenic bacteria are often found in food animals, where their asymptomatic colonization makes detection difficult. Salmonella serotypes have been isolated frequently from cattle, swine, and poultry under many production conditions and environments (Bolton et al., 2012). Ruminant animals, in particular cattle, are the main reservoir for STEC; >$2 billion dollars have been spent by the U.S. cattle industry to combat STEC in processing plants because this pathogen has been declared an adulterant (Kay, 2003).
Specific manipulation of the gastrointestinal microbial population to increase the fermentation efficiency and therefore enhance animal production and growth efficiency has long been a goal of nutritionists and microbiologists (Martin, 1998). Compounds used in this role have included antimicrobials, such as ionophores and antibiotics; but growing negative perceptions around antibiotic resistance limit the use of these type of approaches (Witte, 1999). Organic acids can alter the function and ecology of the intestinal microbial ecosystem (Ricke, 2003; Van Immerseel et al., 2006), often with the result of improving feed conversion and the growth of animals (Piva et al., 2002; Grilli et al., 2010). Because short-chain fatty acids can be utilized in the gastrointestinal tract, they must often be coated to ensure they reach appropriate sections of the gastrointestinal tract (Piva et al., 2007b), and protected acids are more effective than unprotected acids in preventing experimental pathogen colonization (Boyen et al., 2008). Furthermore, coated or microencapsulated acids can be added at a lower inclusion rate because they reach targeted intestinal sections, especially distal portions, and improve animal growth and performance in swine (Piva et al., 2007a, b; Grilli et al., 2010). In previous in vivo studies in weaned pigs, OABP treatment increased average daily gain and final weights, probably mediated by increased feed intake (Grilli et al., 2010).
Because organic acids have been used to control pathogens in foods, to manipulate the gastrointestinal microbial ecosystem, and to improve animal growth efficiency (Goodson and Rowbury, 1989; Harris et al., 2006), it was suggested that they could be used as a nonantibiotic method to reduce the growth of E. coli and Salmonella spp. in live food animals (Van Immerseel et al., 2004a, c; Harris et al., 2006; Boyen et al., 2008) Short-chain fatty acids, medium-chain fatty acids, and other organic acids have an important antimicrobial activity against the most common pathogens of the intestinal tract (Ricke, 2003; Van Immerseel et al., 2006), likely by altering bacterial membrane fluidity and dissipation of proton-motive force (Goodson and Rowbury, 1989; Russell and Diez-Gonzalez, 1998). Organic acids play important roles in Salmonella invasiveness (Durant et al., 2000; Fernández-Rubio et al., 2009), as well as intestinal colonization and shedding (Van Immerseel et al., 2004a, 2005). For example, butyrate supplementation reduced the expression of the pathogenicity island SPI-1 in Salmonella (Gantois et al., 2006), but did not impact the invasive genes hilA and invF; however, acetate addition induced expression of the invasive genes hilA and invF (Durant et al., 2000). Salmonella populations and shedding in broiler ceca were reduced by feeding a variety of organic acids (Van Immerseel et al., 2005). Strains of E. coli, including STEC, are also inhibited by short-chain and volatile fatty acids in many environments (Wolin, 1969).
In the present study, the microencapsulated OABP product was shown to reduce the growth rate and populations of both Salmonella Typhimurium and E. coli O157:H7 in vitro in pure cultures. OABP addition also decreased pathogen populations in mixed ruminal microorganism fermentations when fed at levels similar to what was effective when fed to swine and poultry in previous studies. This product has been shown to improve growth of swine (Grilli et al., 2010), and in the present in vitro study OABP reduced the A:P ratio, which would indicate an improvement in fermentation energetic efficiency in a ruminant animal. Other parameters of ruminal fermentation function were not altered in this in vitro study. Collectively, our data suggest that the organic acid product OABP can be used to improve ruminal or intestinal fermentation efficiency while simultaneously reducing populations of critical foodborne pathogenic bacteria in the mixed microbial population. While no treatment is going to eliminate all foodborne pathogens, organic acid products may have a place as part of an integrated multiple- hurdle pathogen-reduction scheme.
Conclusions
Foodborne pathogenic bacteria are found in live animals and are a significant threat to food safety and human health. Intervention strategies have been sought to reduce carriage of foodborne pathogenic bacteria in food animals, but there is currently not an economic incentive to producers in regard to reducing pathogens in their animals. Nonantibiotic growth-promoting feed additives, such as organic acid products like OABP, can improve animal growth efficiency and were shown in this study to reduce pathogen growth and survival in in vitro fermentations mimicking the ruminal microbial ecosystem. Results indicate that organic acid treatment, such as OABP, could provide an economic advantage to producers as it improves food safety, thus potentially allowing a food safety intervention to “pay for itself.” However, further research is needed to confirm this microbiota-altering effect in live animals of the various food animal species.
Footnotes
Acknowledgments
A grant was provided to support the research by the company Vetagro SpA.
Disclosure Statement
No competing financial interests exist.