
Abstract
Based on recent risk assessments, up to 83% of listeriosis cases from deli meat in the United States are predicted to be from ready-to-eat deli meats contaminated during processing at retail grocery stores. Listeria monocytogenes is known to use sanitizer tolerance and biofilm formation to survive, but interplay of these mechanisms along with virulence potential and persistence mechanisms specific to deli environments had yet to be elucidated. In this study, 442 isolates from food and nonfood contact surfaces in 30 retail delis over 9 months were tested for inlA premature stop codons (PMSCs); inlA encodes InlA, which is necessary to cause listeriosis. A total of 96 isolates, composed of 23 persistent and 73 transient strains, were tested for adhesion and biofilm-forming ability and sanitizer tolerance. Only 10/442 isolates had inlA PMSCs (p<0.001). Strains with PMSCs were not persistent, even in delis with other persistent strains. Most (7/10) PMSC-containing isolates were collected from food contact surfaces (p<0.001); 6/10 PMSC-containing isolates were found in moderate prevalence delis (p<0.05). Persistent strains had enhanced adhesion on day 1 of a 5-day adhesion-biofilm formation assay. However, there was no significant difference in sanitizer tolerance between persistent and transient strains. Results suggest that foods contaminated with persistent L. monocytogenes strains from the retail environment are (1) likely to have wild-type virulence potential and (2) may persist due to increased adhesion and biofilm formation capacity rather than sanitizer tolerance, thus posing a significant public health risk.
Introduction
L
An estimated 83% of listeriosis cases from consumption of ready-to-eat (RTE) deli meats (the leading cause of listeriosis) are from deli meats sliced at retail (Anonymous, 2010; Endrikat et al., 2010), which underscores the need to characterize L. monocytogenes from retail deli environments. L. monocytogenes is frequently introduced into the retail deli environment, which can support its growth and persistence (Gandhi and Chikindas, 2007; Lianou and Sofos, 2007). In a cross-sectional study of 121 retail establishments, L. monocytogenes was found at least once in 60% of stores. In a recent study by our group, we found that L. monocytogenes was present on 14.2% of nonfood contact surfaces (NFCS) and 4.5% of food contact surfaces (FCS) (Simmons et al., 2014). Twelve delis in our study showed evidence of L. monocytogenes persistence based on repeated isolation of isolates with the same pulsed-field gel electrophoresis (PFGE) type on at least three separate sampling dates (Simmons et al., 2014).
L. monocytogenes uses many strategies to survive and persist, including formation of biofilms and increased sanitizer tolerance (Poulsen 1999; Djordjevic et al., 2002; Truelstrup Hansen and Vogel, 2011) in food and food-handling environments. For example, microbial populations in biofilms can be difficult to destroy because they often have greater sanitizer resistance than planktonic organisms (Frank and Koffi, 1990; Lundén et al., 2003; Pan et al., 2006). Sanitizer tolerance (an adaptation allowing growth under sublethal concentrations of a sanitizer) is another important survival strategy (Aase et al., 2000; Soumet et al., 2005; Lourenço et al., 2009). The most commonly used sanitizers in retail delis are quaternary ammonium compounds (QACs) (Tezel et al., 2006; Fraise et al., 2012). However, their effectiveness in eliminating L. monocytogenes may be reduced by the presence of organic residue (inadequate cleaning) or by biofilm formation (Pan et al., 2006).
In this study, we characterized a large set of L. monocytogenes isolates from retail delis to determine whether persistent and transient strains possess different abilities to (1) tolerate QACs and (2) adhere to surfaces and form biofilms, and (3) to determine the virulence potential of isolates from FCS and NFCS to better understand the potential public health risks.
Materials and Methods
L. monocytogenes isolates
A total of 442 L. monocytogenes isolates were assessed in this study (Supplemental Table S1; Supplementary Data are available online at
Adhesion and biofilm formation assay
To assess adhesion and subsequent biofilm formation ability, we performed a 5-day adhesion-biofilm assay (Lemon et al., 2007; Chen, 2013). Day 1 of the assay served as an indicator of adhesion ability where days 3 and 5 were estimates of biofilm-forming capacity. Isolates were grown on BHI agar at 30°C for 24 h; isolated colonies were grown in BHI broth for 17 h at 20°C with shaking at 230 rpm. Cultures were diluted into Hsiang-Ning Tsai medium with 3.0% glucose to yield an OD600=0.05–0.10. An aliquot of 150 μL diluted culture was transferred to each polyvinyl chloride microtiter plate well (Corning Life Sciences, Corning, NY). Plates were incubated for 1, 3, and 5 d at 30°C to facilitate optimal motility and adhesion. On each specified day, the supernatants were removed, the wells were washed 3 times with sterile water and then stained with 0.1% (wt/vol) aqueous crystal violet for 30 min. Wells were washed four times with tap water and air-dried overnight. The stained biomass was suspended in 150 μL of 33% (vol/vol) acetic acid, transferred to a polystyrene plate and scanned at OD590. Isolates were evaluated in triplicate wells (technical replicates).
Sanitizer tolerance assay
Sanitizer tolerance was assessed using a multi-QAC (Clean Quick) (Procter and Gamble, Cincinnati, OH) in a standard broth microdilution assay in tryptic soy broth (TSB) (Becton Dickinson). QAC concentrations ranging from 3.2 ppm to 100 ppm in nutrient-rich and -limiting environments were tested as previously described (Chen, 2013). Isolated colonies were grown overnight on BHI plates at 37°C, then suspended in TSB broth to yield an OD600=0.124-0.140. The suspension was diluted 1:100 and transferred into 2X and 1/5X TSB with 50 μL QAC at a defined concentration to yield final concentrations of 1X and 1/10X TSB simulating nutrient-rich and -limiting conditions, respectively. Plates were scanned at 600 nm at 0 and 24 h to determine the minimum inhibitory concentration (MIC). Isolates that grew in ≥8 ppm QAC after 24 h at 37°C were considered tolerant to sanitizer. This threshold was selected as a conservative definition of tolerant isolates, which was previously defined as ≥4 ppm QAC by To et al. (2002). Sanitizer tolerance was evaluated in technical replicate wells on each plate.
InlA PMSC detection
A total of 442 isolates were characterized by inlA single nucleotide polymorphism (SNP) genotyping using the ABI SNaPshot (ABI, Warrington, UK) protocol as previously described (Van Stelten and Nightingale, 2008). InlA was amplified from lysates (Furrer et al., 1991); purified inlA PCR products served as the template for single base-pair extension reactions using primers targeting the 18 known inlA SNPs (Van Stelten and Nightingale, 2008). Products were analyzed by capillary electrophoresis (ABI 3730xl analyzer) and identified using GeneMarker v 2.4.0 (State College, PA).
Statistical analysis
For adhesion/biofilm formation and sanitizer tolerance assays, the isolates representing the same strain based on longitudinal PFGE data were averaged and categorized into two groups: persistent strains (1) and transient strains (0). Analysis of variance (ANOVA) with Tukey pairwise comparison was used to test the effects of “day,” “type,” and “day*type” interactions for adhesion/biofilm formation ability through use of a multivariate generalized linear model regression in SAS version 9.3 (SAS Inc., Cary, NC). Log10 transformation was applied where appropriate; significance was defined at p<0.05. A linear regression model was used to compare adhesion/biofilm ability on days 1, 3, and 5 postinoculation with nutrient-limiting and nutrient-rich sanitizer tolerance. An ANOVA analysis was also conducted to test the impact of “nutrient,” “type,” and “nutrient*type” on sanitizer tolerance. Each isolate response was categorized as sanitizer tolerant (≥8 ppm) or intolerant (<8 ppm), modeled using a logistic link function, then factor levels were compared using Tukey's adjusted pairwise comparison in SAS (version 9.3). For PMSC data analysis, differences in distribution of PMSCs among isolates from (1) delis with differences in overall prevalence (low [<1%], moderate [1–10%], or high [>10%] [Simmons et al., 2014)]) and (3) from different surfaces (FCS versus NFCS) were analyzed by chi-square analysis or Fisher's exact test (for small samples) using R (RStudio v0.98.943; Boston, MA), where significance was defined at p<0.05.
Results and Discussion
Persistent strains adhere to an abiotic surface significantly better than transient strains
We evaluated the ability of persistent and transient strains to adhere to multiwell polystyrene plates (day 1) and subsequently form biofilms (days 3 and 5). Overall, L. monocytogenes showed significantly greater cell density on day 3 than day 1 (p=0.044; Fig. 1), but similar cell density between day 1 and day 5, and day 3 and day 5 (p>0.05). Furthermore, overall persistent strains were more likely to adhere to polystyrene plates than transient strains were (p=0.0003; Fig. 1). Specifically, on day 1 persistent strains showed significantly greater adhesion ability compared to transient strains (p=0.0330), while no significant differences in biofilm formation capacity existed between persistent and transient strains on day 3 and day 5 (p>0.05). While we acknowledge the limitations of adhesion/biofilm assays as direct indicators of biofilm-forming capacity, persistent strains from food-processing environments in previous studies also showed stronger adhesion/biofilm-forming ability than transient strains (Lundén et al., 2000; Borucki et al., 2003). L. monocytogenes can survive stressful conditions under the protection of biofilms, which increases the potential for cross-contamination to foods in retail deli establishments, and is thus a public health risk (Sofos, 2009; Sofos and Geornaras, 2010; Truelstrup Hansen and Vogel, 2011).
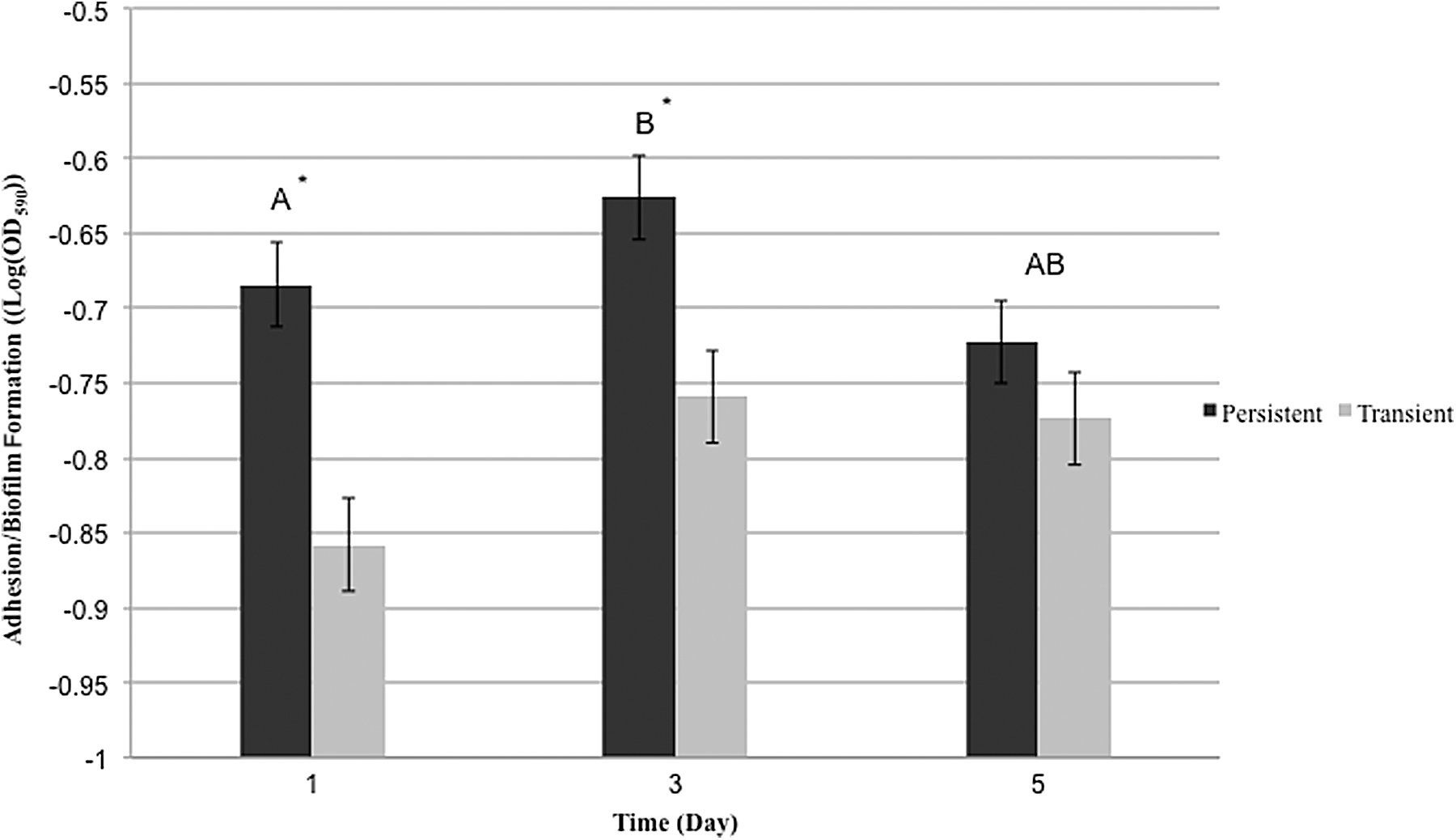
Adhesion and biofilm formation on polystyrene at 30°C as determined by a crystal violet–based staining assay at 1, 3, and 5 days postinoculation. Height of the bars indicates average log(OD590) for both the persistent and transient groups and the error bars represent 1 standard error of means (SEM). The dark bar represents persistent isolates, while the gray bar represents transient isolates. Comparisons were made using Tukey's adjusted comparisons in SAS. Letters represent the comparison results for the ability of isolates to adhere on day 1 and form biofilms on days 3 and 5. An asterisk indicates a significant difference between the persistent and transient isolates (p<0.05).
There is no significant difference between persistent and transient strains' ability to tolerate QAC
We assessed 23 persistent and 73 transient isolates' ability to tolerate QAC sanitizer. There was no significant difference between persistent and transient strains' ability to tolerate QAC (p=0.119; Fig. 2). Furthermore, persistent and transient strains displayed similar sanitizer tolerance under both nutrient-rich and nutrient-limiting conditions (p>0.05; Fig. 2). Not surprisingly, L. monocytogenes isolates collectively were significantly more tolerant to QACs under nutrient-rich conditions than under nutrient-limiting conditions (p=0.036; Fig. 2). In agreement with our study, others found no correlation between persistence and sanitizer tolerance (Holah et al., 2002; Heir et al., 2004). Lundén et al. observed that when L. monocytogenes was exposed to sublethal concentrations of QACs, all strains developed equivalent QAC tolerance over time, with resistance increasing over 28 days (Lundén et al., 2003). Tolerance to QACs is common in L. monocytogenes, with isolates from food-processing environments or foods showing higher MICs than isolates from animals or humans (Mereghetti et al., 2000). However, our understanding of the mechanisms that facilitate sanitizer tolerance is limited. While we appreciate the limitations of testing sanitizer tolerance in a broth medium system, these findings underscore the importance of using recommended sanitizers concentrations with confirmed removal of organic matter that could inactivate sanitizers (Holah et al., 2002; Lundén et al., 2003).
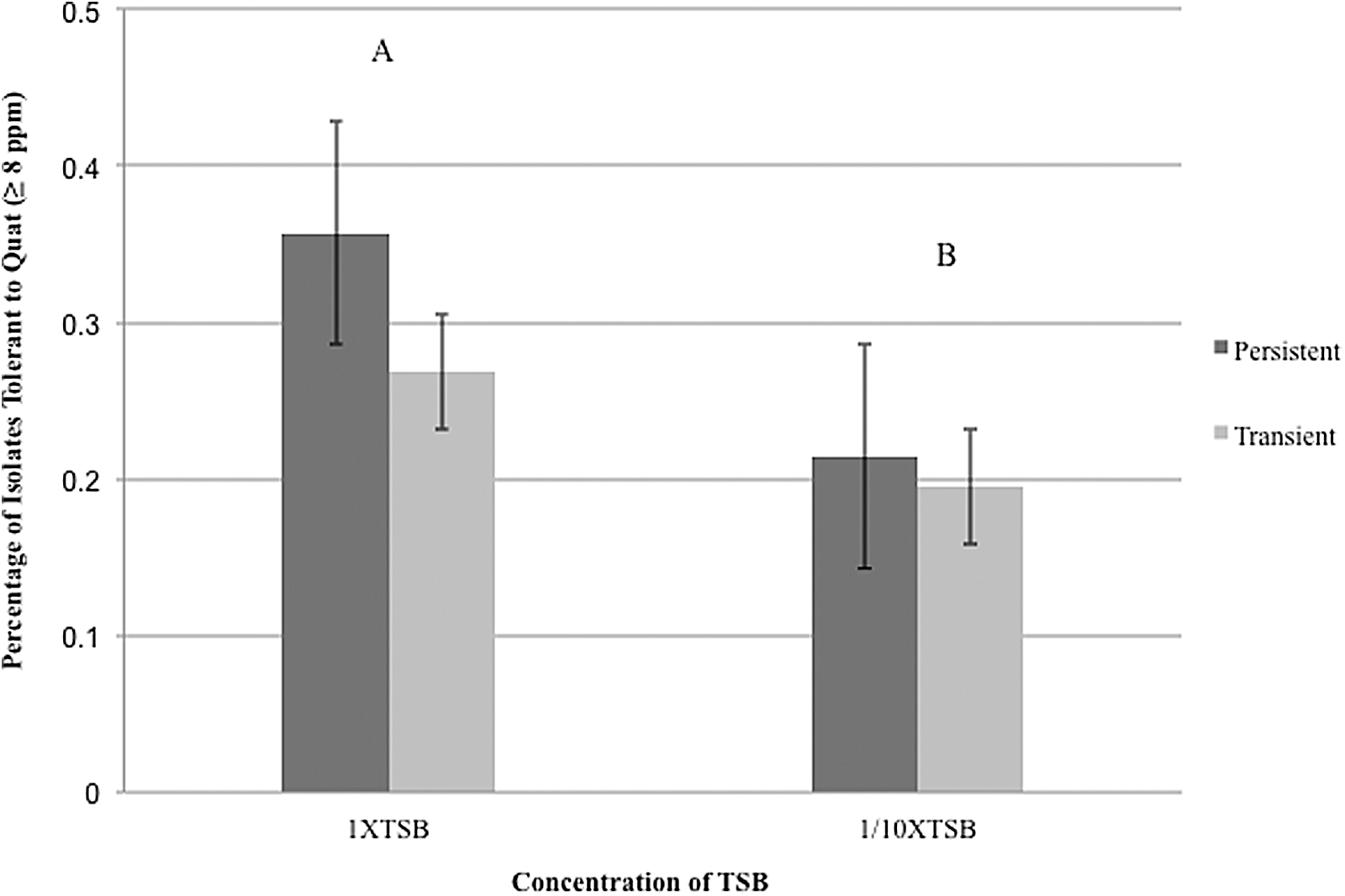
Sanitizer tolerance in 1/10X tryptic soy broth (TSB) and 1X TSB after incubation at 37°C for 24 h. Height of the bars indicate the percentage of persistent or transient isolates that have sanitizer tolerance ≥8 ppm. The dark bar represents persistent isolates, while the light gray bar represents transient isolates. The error bars represent 1 standard error of means (SEM) above or below the mean. Comparisons were made using output from the Tukey's test in SAS. Letter represents the comparison results for sanitizer tolerance between different concentrations of TSB.
Biofilm-formation ability is negatively correlated with sanitizer tolerance after long-term incubation in nutrient-rich conditions
We hypothesized that enhanced cell adhesion and biofilm formation may increase sanitizer tolerance, which could enhance survival of L. monocytogenes in food-handling environments. Interestingly, on day 5 of the adhesion/biofilm assay, L. monocytogenes isolates with less biofilm-forming capacity were more tolerant to QACs (p=0.032, r 2=0.0392; Fig. 3). Greater cell adhesion and biofilm formation was not significantly associated with sanitizer tolerance under nutrient-rich or nutrient-limiting conditions on day 1 and day 3, respectively (p>0.05). These results agree with a previous study on the resistance of L. monocytogenes biofilms to sanitizers, which found that during the first week, biofilms were susceptible to sanitizers and cell density decreased significantly, but the resistance of biofilms to QACs progressively increased with time (Pan et al., 2006). While our study design was limited to 5 days without long-term adaptation to sanitizer, it suggests that sanitizer tolerance and adherence and biofilm formation are distinct mechanisms for promoting L. monocytogenes survival and persistence in the retail deli environment.
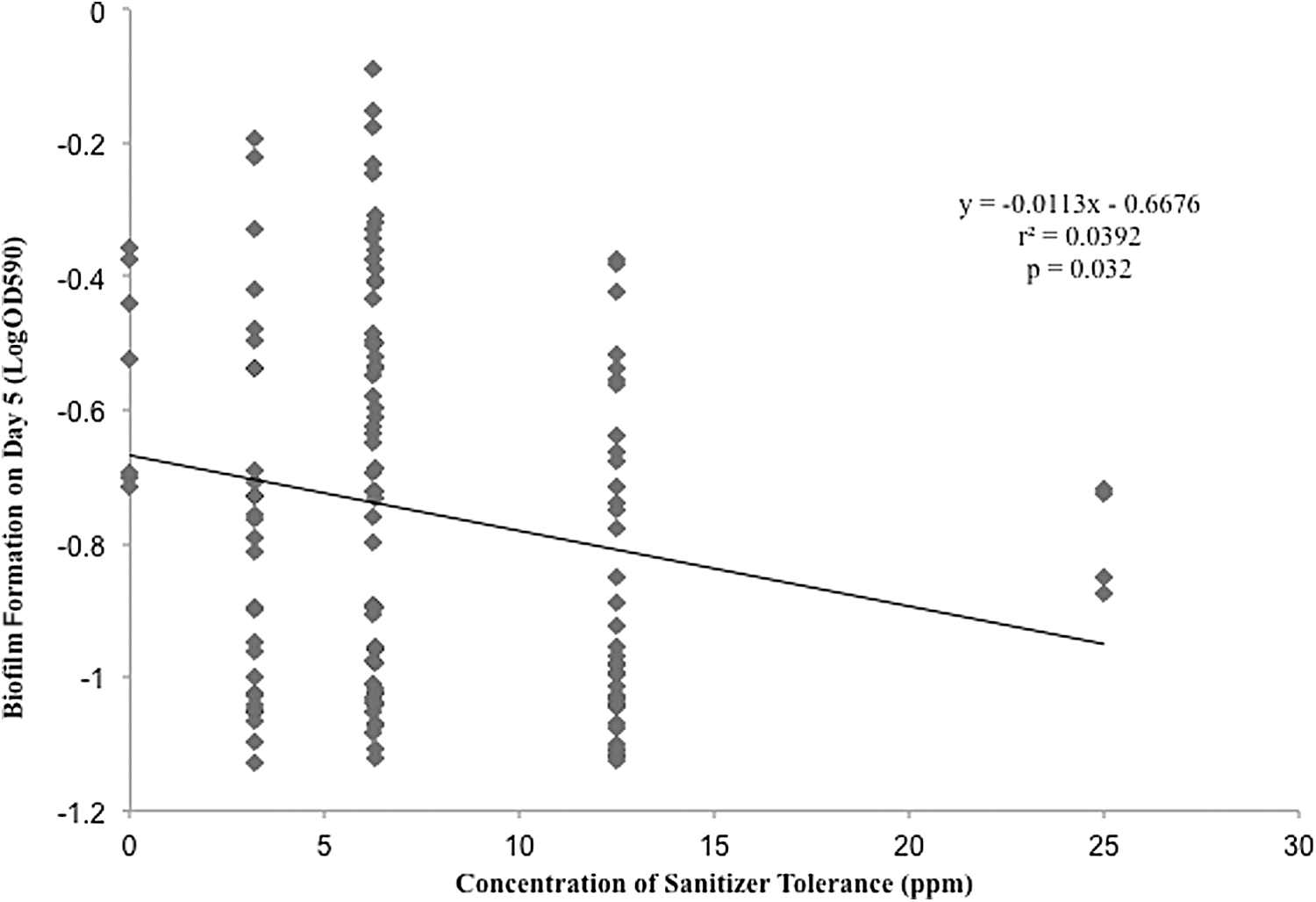
Relationship between biofilm formation (logOD590) on day 5 and sanitizer tolerance in nutrient rich conditions. Y-axis indicates biofilm formation level (logOD590). X-axis indicates the concentration of sanitizer tolerance. A linear regression line was drawn to show the negative correlation between isolates ability to form biofilms on day 5 and tolerance to sanitizer in nutrient-rich conditions.
InlA PMSC mutations are extremely rare in isolates from the retail deli environment
A total of 10/442 (≈2.3%) of retail deli isolates contained PMSCs, indicating a clear underrepresentation in the population compared to wild-type (chi square; p<0.001; Table 1). In agreement, a study by Neves et al. found that all isolates from a cheese-processing environment had virulence potential or were fully virulent (Neves et al., 2008). A total of 7/30 delis represented in this study had isolates with PMSC (chi-square analysis comparing stores with and without PMSCs isolates; p<0.001); only 2 delis had more than 1 isolate with a PMSC. Nine of the 10 PMSCs identified were PMSC-4 and 1 was PMSC-3, the second and first most common PMSCs in previous studies, respectively (Van Stelten et al., 2010). Isolates containing PMSC-3 and PMSC-4 produce a truncated InlA protein and have <10% wild-type invasion efficiency (Van Stelten and Nightingale, 2008; Van Stelten et al., 2010). We hypothesized that cross-contamination from the retail deli environment may be the source of isolates with PMSCs, as up to 45% of isolates from RTE foods have PMSCs (Van Stelten et al., 2010). However, our data suggest that contamination of RTE foods with PMSC-containing isolates occurs prior to or after handing in the retail deli environment.
Store (deli) identifier among 30 delis sampled by Simmons et al. (2014).
Listeria monocytogenes prevalence for delis containing PMSC+ isolates from Simmons et al. (2014) where “high” ≥10%, “moderate” >1% and <10%, and “low” ≤1% overall prevalence for the deli.
Total isolates typed within each deli irrespective of sample location.
Total isolates with inlA PMSC within each deli.
Total food contact surface isolates found within each deli.
inlA PMSC+ isolates found on food contact surfaces within each deli.
Total nonfood contact surface isolates within each deli.
inlA PMSC+ isolates on nonfood contact surfaces within each deli.
PFGE type (Asc1-Apa1) assigned by Simmons et al. (2014) for isolates containing an inlA PMSC.
Sample source within deli for isolates containing PMSCs from Simmons et al. (2014).
LM, Listeria monocytogenes; FCS, food contact surfaces; NFCS, nonfood contact surfaces.
Isolates containing inlA PMSCs are transient strains and are more common on FCS
No isolates with PMSCs met the persistence criteria, and isolates containing PMSCs had rare PFGE types (Simmons et al., 2014). Of the 442 isolates tested, 73 (16.5%) were from FCS, while 369 were from NFCS. Less than 1% of NFCS isolates (3/369) contained PMSCs compared to approximately 9.6% of FCS isolates (7/73) (Fisher's exact test; p<0.001). Previous research has indicated that the majority of FCS isolates are from lineage II, while the majority of NFCS isolates are from lineage I (Hoelzer et al., 2011). In the United States, lineage II isolates are often isolated from food, and tend to have a high proportion of PMSCs, whereas lineage I isolates are frequently associated with human disease and rarely contain PMSCs (Gray et al., 2004; Jacquet et al., 2004). Consequently, our results underscore the risk associated with cross-contamination of FCS from NFCS, which has been observed in other studies (Hoelzer et al., 2011; Simmons et al., 2014).
Isolates with PMSCs were more common in moderate-prevalence delis (overall prevalence 1–10%) than in high-prevalence delis (≥10%) (Fisher's exact test p<0.05). Specifically 4.0% (4/99) of isolates in high-prevalence delis had PMSCs compared to 15.8% (6/38) of isolates collected from moderate-prevalence stores (Table 1). However, the likelihood of a store containing an isolate with a PMSC was not significantly different among categories of environmental L. monocytogenes prevalence as defined in Simmons et al. (Simmons et al., 2014). Prevalence was significantly associated with the proportion of FCS isolates in delis (Fisher's exact test, p<0.0001). Specifically, moderate-prevalence stores had a higher proportion of FCS isolates with PMSCs; 15.6% (5/32) of FCS isolates from moderate-prevalence delis had PMSCs compared to just 5.1% (2/39) in high-prevalence delis (p>0.05). Our data indicate that the probability of an isolate from a FCS to have a PMSC is similar regardless of L. monocytogenes prevalence within the deli, suggesting that these isolates do not originate within the deli, but may be brought in from outside the deli, perhaps from contaminated food products or trafficked into the deli by carts, employees, etc.
Isolates representing a single strain display significant variation in sanitizer tolerance and adhesion and biofilm formation ability, but not in inlA SNPs
Isolates of the same strain showed considerable variation in adhesion and biofilm formation ability (Fig. 4) and sanitizer tolerance (Fig. 5). Since sanitizer tolerance and adhesion and biofilm formation capacity can be an adaptive mechanism (Lundén et al., 2003), intra-strain variation is not surprising as previously described (Afzal Ghauri et al., 2007; Adrião et al., 2008). While no isolates with PMSCs were persistent, two isolates from a single store had PMSCs; both isolates were from the one-basin sink interior (a FCS) and had PMSC-4. Additionally, PFGE type CU-200-227 was found in three delis; it was associated with a FCS site and possessed PMSC-4 in each deli. This further supports that all three isolates of this pulsotype were the same strain, and may have originally been associated with a food product or other common source.
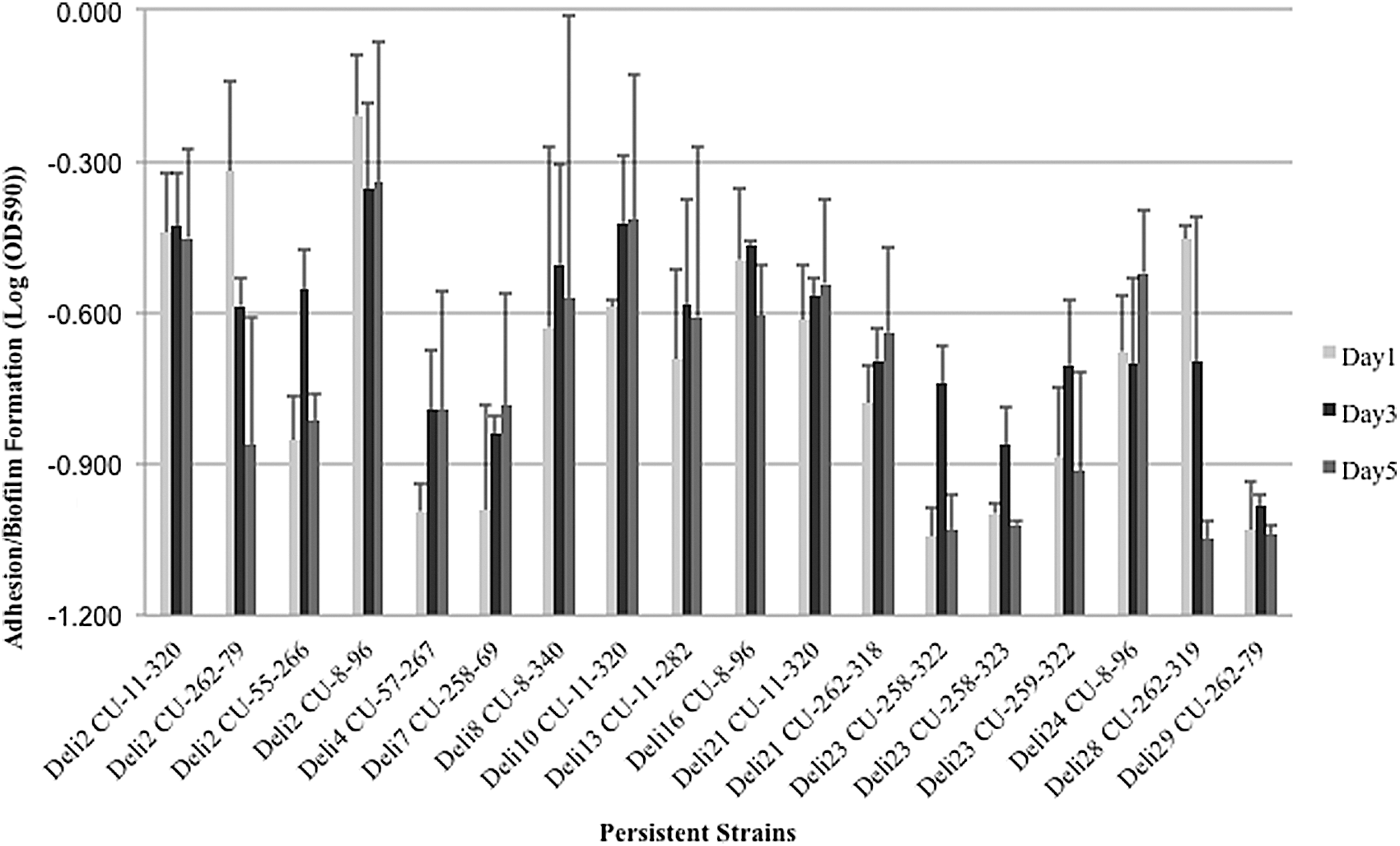
Variation in adhesion and biofilm formation capacity among isolates of the same persistent Listeria monocytogenes strain. Height of the bars indicates average log (OD590) for isolates within same persistent strains and the error bars represent 1 standard error of means above the means. The light bar represents cell adhesion on day 1 postinoculation; the dark bar represents biofilm formation on day 3 postinoculation; the gray bar represents biofilm formation on day 5 postinoculation.
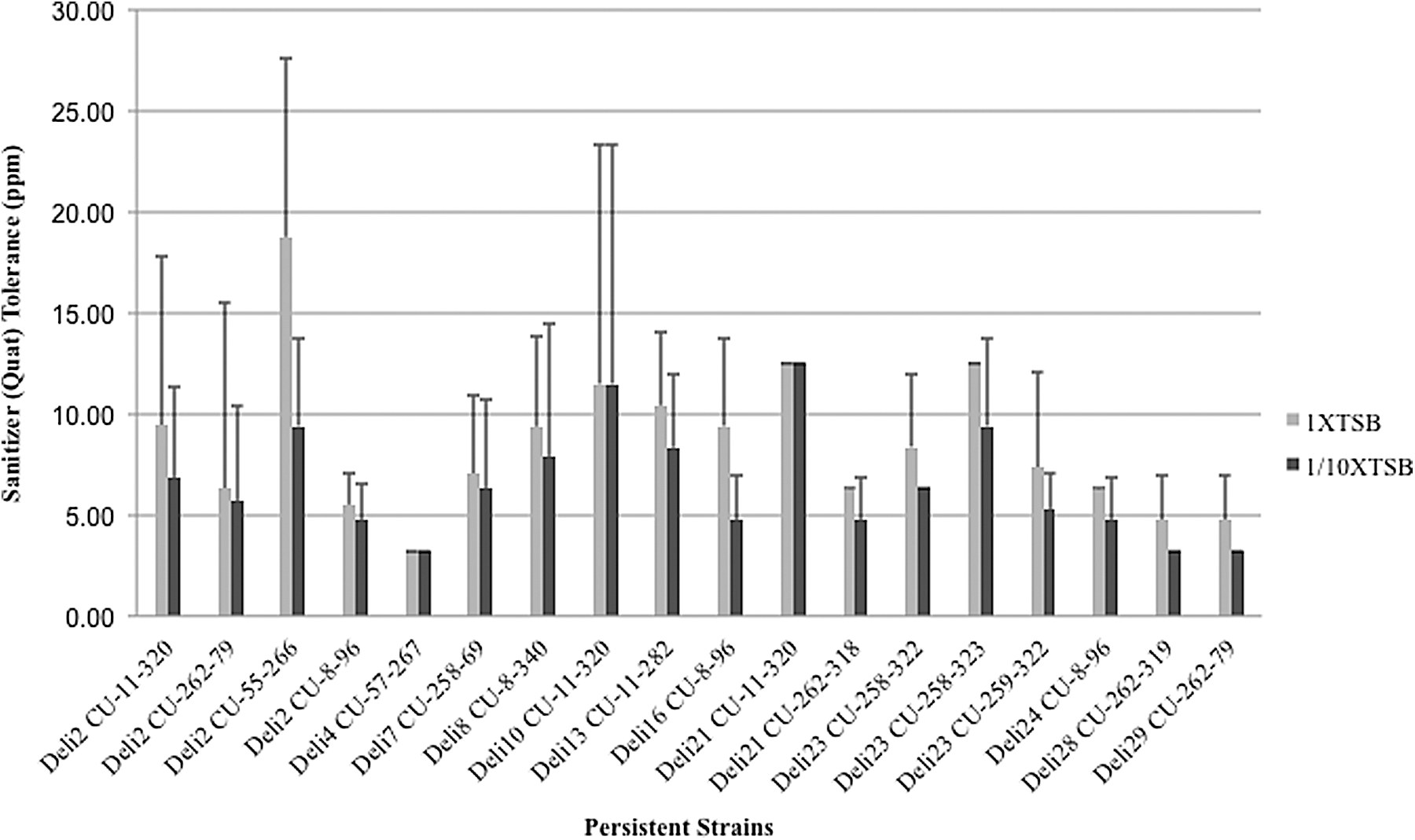
Variation among isolates within same persistent Listeria monocytogenes strains based on sanitizer tolerance assay. Height of the bars indicates average sanitizer tolerance for isolates within same persistent strains and the error bars represent 1 standard error of mean above the means. The light bar represents sanitizer tolerance in nutrient-rich conditions (1X tryptic soy broth [TSB]); the dark bar represents sanitizer tolerance in nutrient-limiting conditions (1/10XTSB).
Conclusions
In this study, we characterized persistent and transient strains from retail deli establishments. We found that persistent L. monocytogenes strains adhered to abiotic surfaces better than transient strains on day 1, and a negative correlation was found between enhanced cell biofilm formation on day 5 and sanitizer tolerance under nutrient-rich conditions. These observations add to the understanding of the ability of L. monocytogenes to persistent in the retail food environment. We also found that inlA PMSCs were very rare among retail deli environment isolates and were strongly associated with transient strains, isolates from food contact surfaces, and isolates from moderate-prevalence delis. These results indicate that the vast majority of isolates from the retail delis environment, including those from FCS, have virulence potential and that isolates with PMSCs are unlikely to persist in the retail deli. To ensure food safety in delis, we suggest future work be directed toward developing strategies to reduce L. monocytogenes persistence. Meanwhile, practices that keep L. monocytogenes at reduced levels including hygienic design, prevention of cross-contamination, effective cleaning and sanitation, and temperature control are critical to preventing listeriosis resulting from consumption of deli meats contaminated at retail.
Footnotes
Acknowledgments
We acknowledge Xiaosu Tong for statistical analyses consultation. This project was supported by the Food Marketing Institute (Purdue University grant #206298) and the U.S. Department of Agriculture–Agricultural Research Service (#1935-42000-049-60D).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.