
Abstract
This study was performed in order to determine whether human isolated probiotic bacteria can be effective in reducing Campylobacter jejuni infection of chicken intestinal cells, in vitro, and in decreasing its colonization abilities within the chicken gut. Our results show that the probiotic strains Lactobacillus paracasei J.R, L. rhamnosus 15b, L. lactis Y, and L. lactis FOa had a significant effect on C. jejuni invasion of chicken primary cells, with the strongest inhibitory effect detected when a combination of four was administered. In regard to the in vivo effect, using all four strains in one combination prevented mucus colonization in the duodenum and cecum. Moreover, the pathogen load in the lumen of these two compartments was significantly reduced. When probiotics were introduced during the early growth period, the presence of the pathogen in feces was increased (p>0.05), but when they were given during the last week of growth, there was no significant effect. In conclusion, our data indicate that these four new probiotic strains are able to cause modifications in the chicken intestinal mucosa and can reduce the ability of C. jejuni to invade, in vitro, and to colonize, in vivo. These probiotics are now proven to be effective even when introduced in broiler's feed 7 days before slaughter, which makes them cost-effective for the producers.
Introduction
C
Poultry (ducks, turkeys, and chickens) can rapidly become colonized with Campylobacter, and in general, each broiler cycle starts with a negative flock (Wagenaar et al., 2006). Practical biosecurity measures in farms are proactively necessary prior to their arrival from hatchery, for once Campylobacter is introduced in the flock it spreads rapidly, and up to 100% of broilers at slaughter age can harbor the human pathogen (Jacobs-Reitsma, 1995). Other preventive measures aim to control Campylobacter colonization in poultry, and these include water treatment (Chaveerach et al., 2002), diet modification and feed additives (van de Giessen et al., 1998), vaccination (Heres et al., 2004), passive immunization (Buckley et al., 2010), and use of probiotics (Stern et al., 1990; Alemka et al., 2010) and prebiotics (Santini et al., 2010).
One very effective measure involves the inclusion of live microorganisms (probiotics) in poultry feed. When administered in adequate amounts, probiotic microorganisms can provide a beneficial effect to the host digestive system (Ventura et al., 2009). These beneficial effects have been described in some cases of viral-induced diarrhea (Szajewska, 2007) as well as preventive effects in Clostridium difficile–associated diarrhea (Parkes et al., 2009). The probiotics act in the intestine through their high affinity for the epithelial mucus layer by producing inhibitory compounds and adjusting the immune system (Patterson and Burkholder, 2003). In artificially contaminated broilers, it has recently been shown that probiotics can be successfully used to reduce pathogen load (Neal-McKinney et al., 2012).
Although the beneficial effect of probiotics is controversial, their role in improving the productive parameters and their antipathogenic effect is well characterized. The aim of this study is based on one of our recent observations that four new human isolated probiotic strains (Lactobacillus paracasei J.R, L. Rhamnosus 15b, L. lactis Y, and L. lactis FOa) have the ability to reduce C. jejuni invasion of chicken primary cells. Based on this initial outcome, we investigated the possibility of reducing the presence of C. jejuni in naturally contaminated farmed broilers by analyzing its presence in the feces and the intestinal contents.
Materials and Methods
Bacterial strains and culture conditions
The following strains were used in this study: C. jejuni 81-176, C. jejuni NCTC 11168, and C. jejuni NCTC 11168H. Bacteria were grown on Mueller-Hinton agar (CM0337 Oxoid) at 37°C under microaerophilic conditions generated by Campygen gas packs (CN0025 Oxoid). Human isolated probiotic bacteria (L. paracasei J.R, L. rhamnosus 15b, L. lactis Y, and L. lactis FOa) were grown in MRS broth media (CM 0359, Oxoid) at 37°C for 24 h and lyophilized. The lyophilized probiotics were incorporated in the feed and provided 1×109 colony-forming units (CFU)/kg feed (1%) using an automatic mixing unit.
Quantification of C. jejuni in animal samples
C. jejuni presence was analyzed in feces, cecal, and duodenal content and also in the cecal and duodenal mucus. In order to quantify the pathogen presence in the cecal and duodenal content, 100 μL of content was diluted in sterile phosphate-buffered saline (PBS) and serial dilutions were plated on Mueller-Hinton agar. For the quantification in the mucus, 100 μL of cecal and duodenal mucus was diluted in sterile PBS and plated on Mueller-Hinton agar plates. Plates were incubated at 37°C in microaerophilic conditions.
Bird trial
A total of 280 chicken broilers (ROSS 308) were allocated randomly in 7 groups. Each group had five replicate floor pens with eight chicks per pen. The feeding program (Table 1) included the starter (0–10 days), grower (11–35 days), and finisher diets (36–42 days). Mixed-gender chicks were allowed ad libitum access to feed and water during the experiment (Table 2). Body weight, weight gain, and feed intake of each replicate were recorded at the 10, 35, and 42 days. At the end of the 42-day experiment, 3 birds per pen were randomly selected and slaughtered. Duodenal and cecal content as well as the mucus of these compartments was analyzed for identification of the Campylobacter jejuni. The experiments were performed according to the Romanian legislation in place (Law 471/2002 and government ordinance 37/2002) and under the supervision of National Sanitary Veterinary Agency. The ethics committee of Banat's University of Agricultural Sciences and Veterinary Medicine–King Michael I of Romania approved this work.
CF, crude fiber; CP, crude protein; ME, metabolizable energy.
L., Lactobacillus.
Isolation of chicken intestinal cells
Chicken intestinal primary cells were isolated as previously described (Clyne and Drumm, 1993; Byrne et al., 2007). Briefly after slaughter, sections of the intestine were excised and washed in cold saline (PBS). Biopsies were taken using endoscopy forceps and placed into primary cell culture medium at 4°C for transport. Primary chicken intestinal cells were grown in medium comprising Dulbecco Modified Eagle Medium-F12 and 10% fetal calf serum. Primary cells were washed twice in warm antibiotic-free primary medium to remove nonadherent cells and incubated in antibiotic-free medium for a minimum of 3 h prior to infection.
In vitro invasion studies with probiotics and C. jejuni
The invasion studies in the presence of probiotics were performed as previously described (Alemka et al., 2010). Briefly, probiotics were grown in Man Rogosa Sharpe broth for 6 h under microaerophilic conditions, and harvested by centrifugation at 5000 rpm for 5 min and resuspended in prepared tissue culture medium (RPMI 1640) at an OD600 of 0.4. A volume of 300 μL of probiotics in tissue culture medium was used to infect chicken primary cells for a period of 4 h. The medium on the cells, containing probiotics, was then replaced with 300 μL of C. jejuni culture (OD600 of ≈0.3). Infected cells were further incubated for 6 h. As a control, we have used probiotic nonpretreated cells. Following invasion, Mueller Hinton agar supplemented with Campylobacter selective supplement (Skirrows) was used to select for C. jejuni CFU.
Microscopy
The intestinal wall from cecum and duodenum was investigated for villus height, the height of intestinal epithelium, microvilli height (large and small diameter), and also the perimeter of goblet cells. Samples were fixed in 10% neutral formalin and included in paraffin blocks. The paraffin block sectioning was carried out using the Leica microtome, at a thickness of 4 μm. The histological sections were stained with Mallory's trichrome and Alcian blue method and were examined under Olympus CX41 research microscope fitted with digital camera and analyzed using the Quick PHOTO Micro2.2 software for the histomorphometry.
Statistical analysis
Experiments were conducted on at least three separate occasions in triplicates. Results are presented as the means±SD (error bars) of replicate experiments. Graphs were drawn using GraphPrism, and the unpaired Student t-test was used to estimate statistical significance. A p value of<0.05 was considered significant. The statistical analysis of the histomorphometry results was performed using the ANOVA and Tukey-SPSS IBM22 tests. The results are presented as the mean±SE. Differences were considered to be statistically significant at p<0.05, distinct significant at p<0.01 and very significant at p<0.001.
Results
Probiotic effects on chicken primary epithelia cells in vitro
C. jejuni is known for its capacity to colonize the chicken intestinal mucosa, and recent studies have shown that this human pathogen is able to invade primary avian intestinal cells (Byrne et al., 2007). Probiotic bacteria are known to reduce C. jejuni attachment to human intestinal epithelial cells in vitro (Alemka et al., 2010); however, this effect has not been previously shown with chicken epithelial cells. Combinations of two and four probiotic strains thereof were investigated for their effect on the pathogenicity of C. jejuni 81-176, C. jejuni 11168, and C. jejuni 11168H using chicken primary cells. Two sets of strains (L. paracasei J.R–L. rhamnosus 15b [Set A] and L. lactis Y–L. lactis FOa [Set B]) were used in combination to initially colonize primary chicken cell monolayers for 6 h. These two sets of isolates reduced the internalization of C. jejuni from ≈104 CFU/well to ≈103 CFU/well. We have next investigated a combination of four probiotic strains (Set A+Set B). The results show a dramatic decrease in invasion (≈102 CFU/well) compared to the control experiment (Table 3). The differences in invasion were significant (p<0.05) compared to an invasion assay performed in the absence of probiotics.
CFU, colony-forming units; C., Campylobacter; L., Lactobacillus.
C. jejuni load in broilers feces and intestines is decreased by probiotics
Our experiments aimed to compare the inhibitory effect of probiotics on pathogen colonization and retention in the intestine. When probiotics were introduced in the diet at the beginning of the growth period (Fig. 1, Day 0), there was a marked increase in the presence of C. jejuni in the feces (Fig. 1A). The presence of the pathogen in the feces was higher in L2 (L. paracasei J.R and L. rhamnosus 15b, L. lactis Y, and L. lactis FOa) and L5 (L. paracasei J.R and L. rhamnosus 15b, L. lactis Y, and L. lactis FOa+25% threonine and methionine) compared to L1 (L. paracasei J.R and L. rhamnosus 15b). In the experimental group L5, there was an increase of C. jejuni in the feces; however, the differences were not significant compared to the control (p>0.05). We have next investigated the effect of the probiotic isolates on the presence of C. jejuni in the duodenal (Fig. 1B) and cecal (Fig. 1C) content. Our results indicated a massive decrease in C. jejuni presence in these two intestinal compartments. Based on these results, we hypothesized that the decrease in the pathogen presence in the duodenal and cecal content should also have an impact on the pathogen colonization of the intestinal mucus layer. Indeed, as shown in Figure 1D and E, the presence of the pathogen in the mucus layer was scarcely detectable. The inclusion of synthetic amino acids in the diet had no significant effect on the pathogen load.
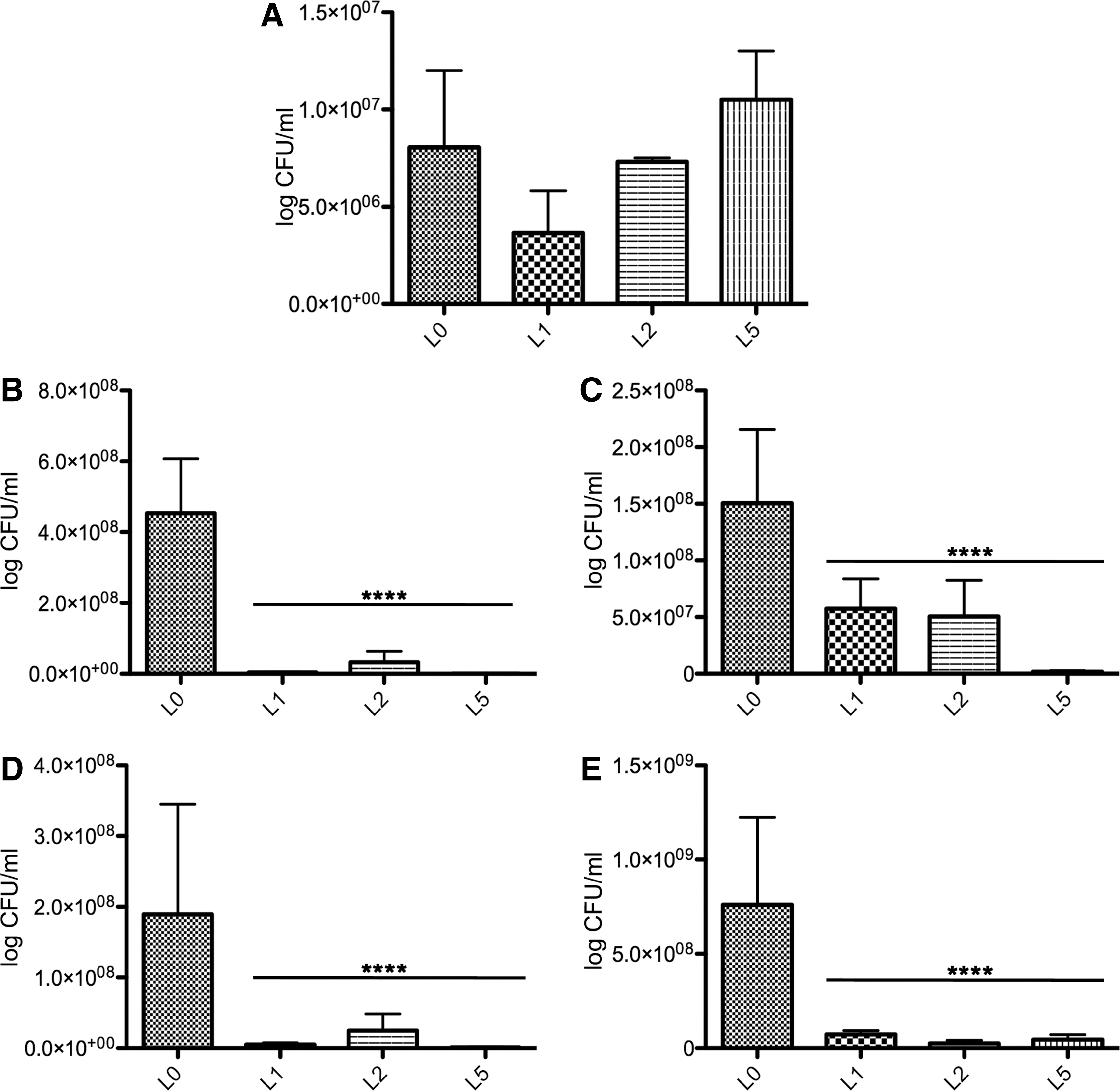
The effect of probiotics, included in broilers diet at day 0, and its effect on of Campylobacter jejuni load in the feces
The inclusion of probiotics in the diets of the experimental groups L3, L4, and L6 in the last week of growth had a significant negative effect on the pathogen load in the feces (Fig. 2A). The effects on the pathogen presence in the cecal and duodenal content or in the mucosa were similar to when they were introduced in their diets at the beginning of the growth period (Fig. 2B–E). The addition of threonine and methionine for groups L5 and L6 had no visible effect on colonization. These amino acids are important in the biosynthesis of mucin glycoproteins. Our results suggest that these probiotic isolates can be used in broiler's diets only in the last week before slaughter with a significant financial effect for the producers.
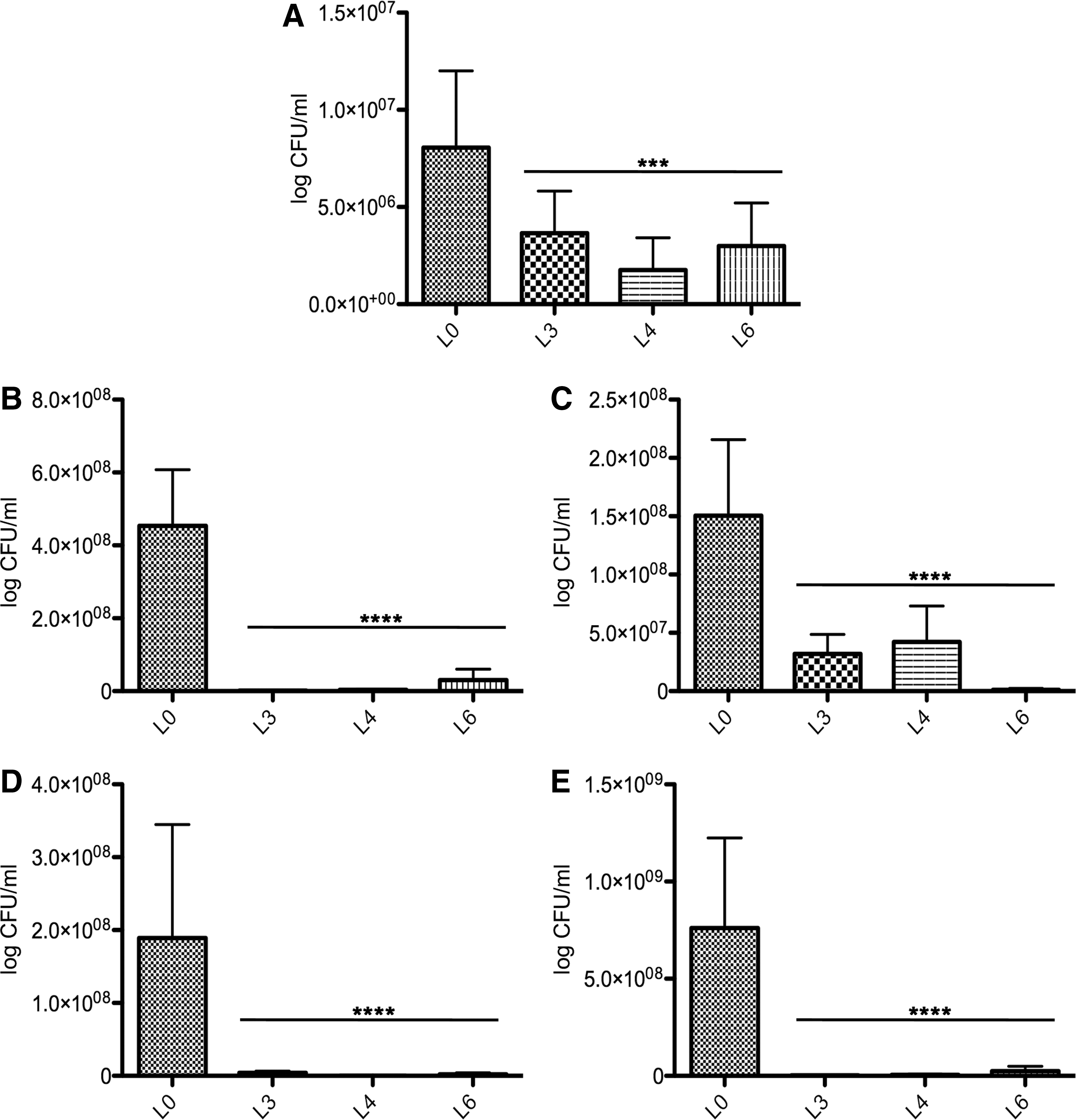
The effect of probiotics, included in broilers' diet in the last week of growth, and its effect on Campylobacter jejuni load in the feces
Effect of probiotics on poultry intestinal structures
The control group (L0) showed large goblet cells with granular cytoplasm (Fig. 3). In the cecum, the average thickness of the mucosa was 157.50±32.93 μm. This was significantly lower compared to the mucosa of all the other experimental groups (Fig. 4) (data not shown).
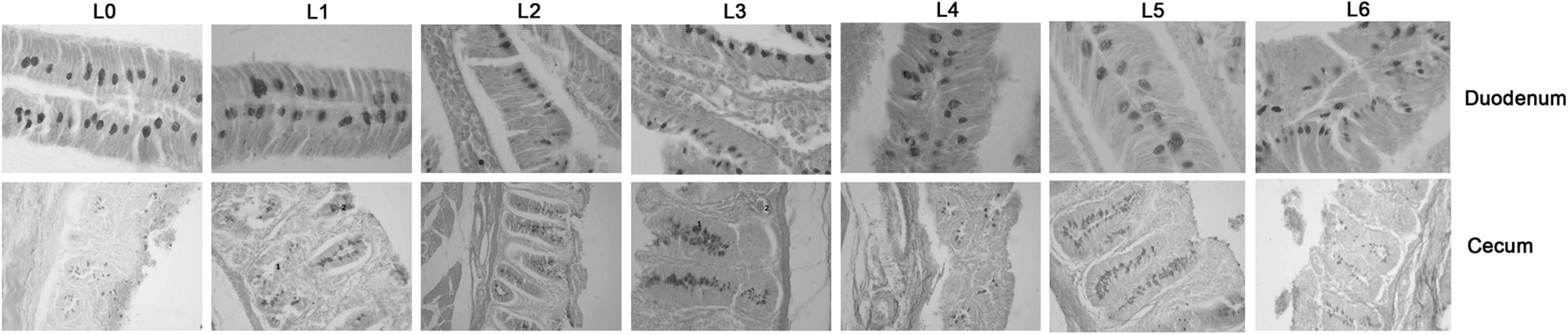
Goblet cells appearance following investigation of the duodenal and cecal sections. L0 is the control group, and L1–L6 are the experimental groups.
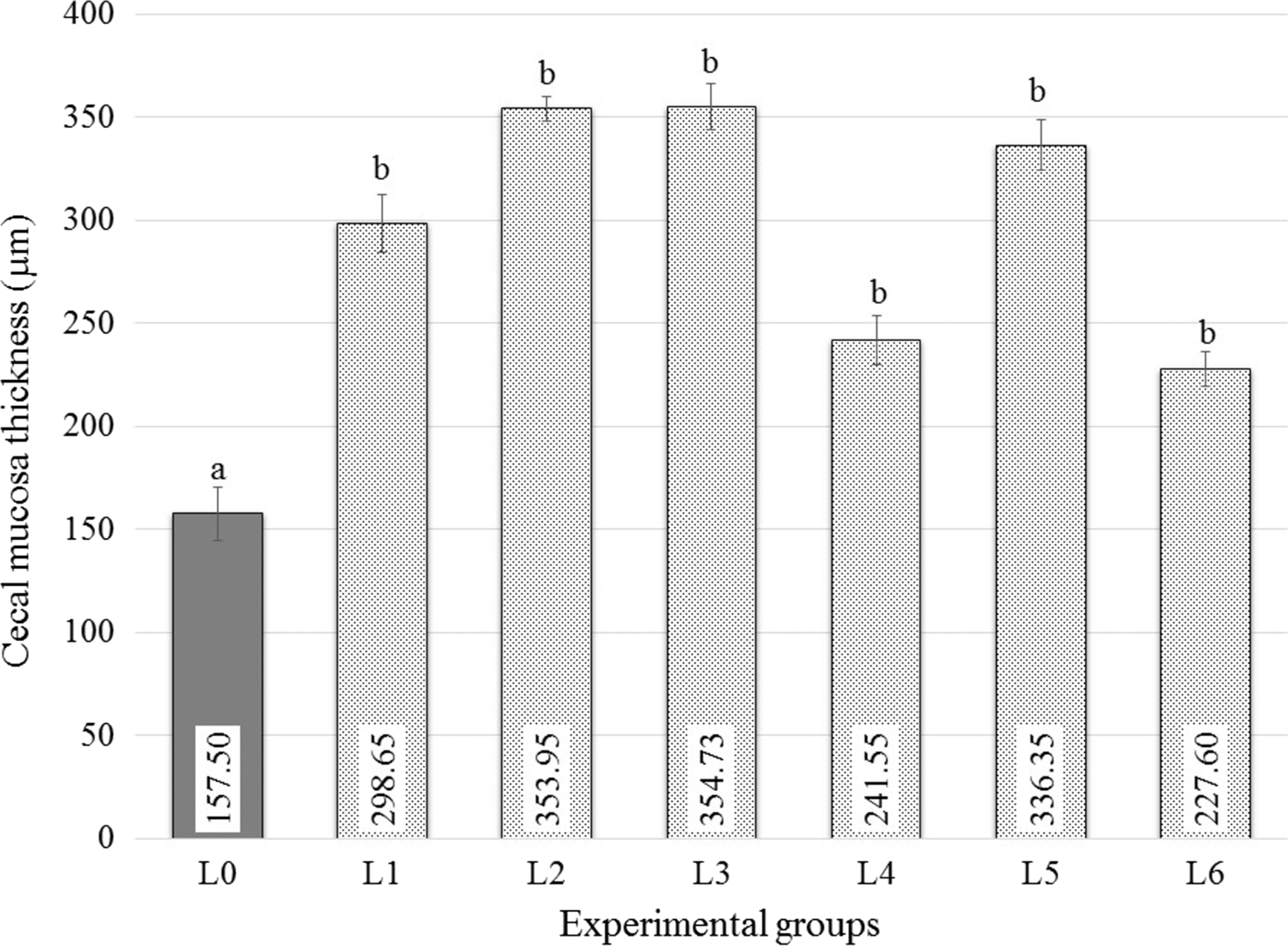
Thickness of cecal mucosa. The statistical analysis of the histomorphometry results was performed using the analysis of variance and Tukey-SPSS IBM22 tests. The numbers within the bars represent the thickness as measured and the letters represent the levels of significance.
In the experimental group L1, the goblet cells were large, with intense granular cytoplasm but with no significant differences between L0 and L1. In the villous epithelium, the goblet cells were large, with intense granular cytoplasm. The mucosa in the cecum was significantly thicker than L0 (p<0.05) and the goblet cells show granular cytoplasm (Figs. 4 and 5). In experimental group L2, the goblet cell area was smaller than L0 (p<0.05) with intense granular cytoplasm and leukocytes in the intraepithelial space. In the cecum, the mucosa had a hypertrophic appearance and the goblet cells displayed an intense granular cytoplasm (data not shown).
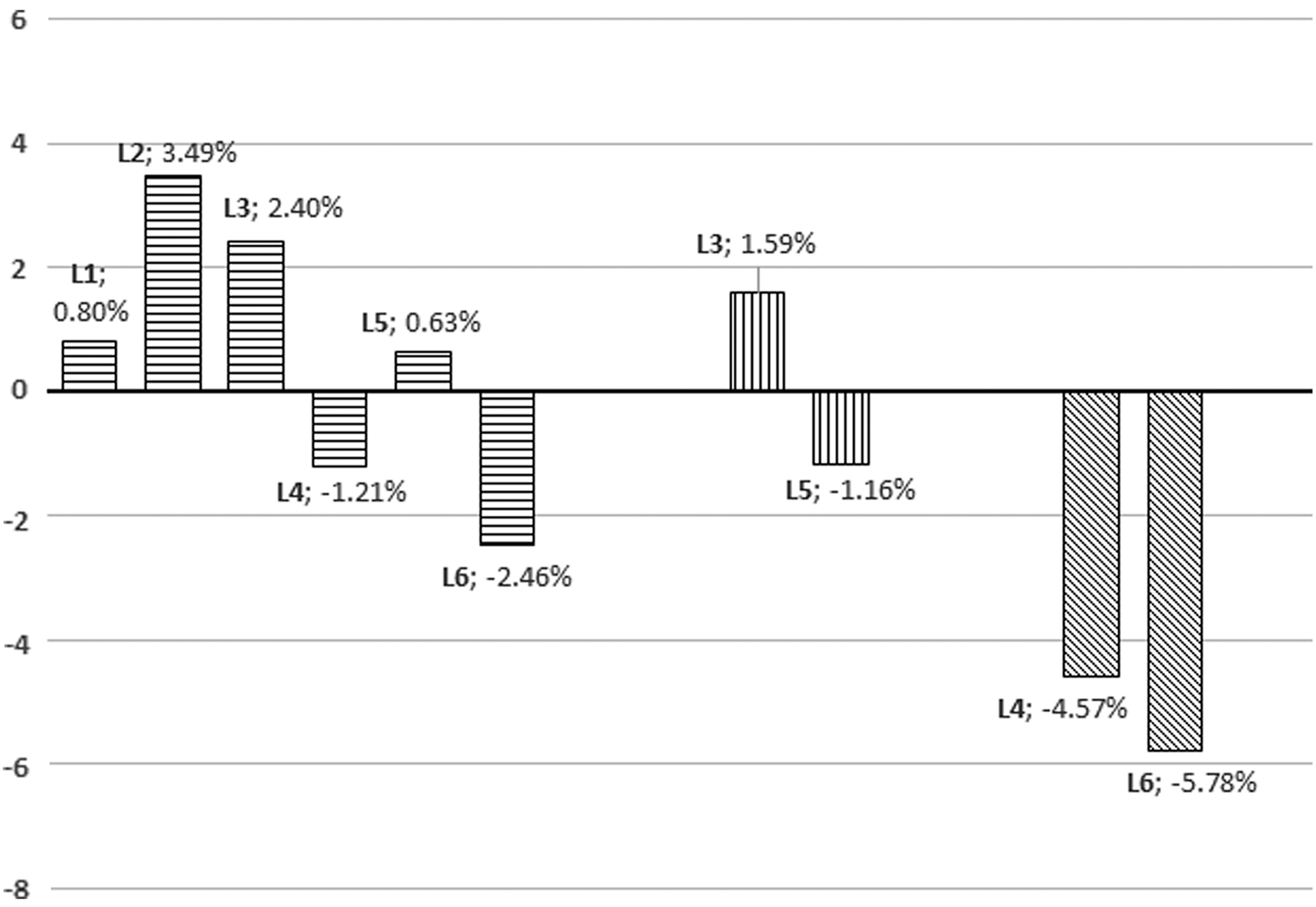
Percentage differences between the body mass mean values (control group L0 versus experimental groups L1–L6; experimental group L1 versus L3 and L5; and experimental group L2 versus experimental groups L4 and L6). The numbers represent significance as calculated in percentage.
In the experimental group L3, the goblet cells were smaller (p<0.05) with regular cytoplasmic appearance. The mucosa had a hypertrophic aspect and, in the chorionic mucosa, abundant leukocyte infiltrate was present. The cytoplasm of goblet cells had a granular aspect. In the L4 group, the goblet cells had a granular aspect leukocyte infiltration. In the experimental L5, the visible leukocyte infiltrate and capillary ectasia were present. The goblet cells were numerous in the epithelium, with many of them displaying granular cytoplasm. In the experimental group L6, the goblet cells were large with granular content. An abundant leukocyte infiltration was visible in the chorionic mucosa. The cecum mucosa is thicker compared to the control group (Fig. 4) with leukocyte infiltrate.
We can conclude that the presence of Campylobacter in the chicken intestine can be attenuated with probiotics. They can induce modifications in the thickness and structure of the intestinal mucosa (Fig. 4) with a proven effect on the number of campylobacters found in the intestinal compartments of the experimental broilers.
The new isolated probiotic strains have no effect on broilers' growth performance
Recent studies have shown that in the case of chicken broilers artificially infected with C. jejuni, they display a decrease in weight gain which is most likely caused by a decrease in nutrient absorption at the intestinal level (Awad et al., 2014). The graphical representation of body mass differences (Fig. 5) shows that the experimental groups L2 and L3 have a higher average value of body mass (3.49% and 2.40%) when compared to the control group (L0). The body mass of chickens from groups L1 and L5 was very similar to the control group (L0), with L1 and L5 being only 0.80% and 0.63% higher, respectively. The experimental groups L4 and L5 recorded a lower body weight at the end of the experimental period, being 1.21% lower for L4 and 2.46% lower for L5; however, the differences were not statistically significant (p>0.05).
Discussion
C. jejuni, a Gram-negative bacterium, is often responsible for human gastroenteritis (Tauxe, 2002). Chickens are recognized as the main source for human infections because when they are contaminated with Campylobacter clinical signs are not visible (Wagenaar et al., 2013). Any attempt made toward reducing colonization in poultry can potentially reduce the incidence of campylobacteriosis in humans. The use of probiotics to competitively exclude C. jejuni has been proposed for poultry (Schoeni and Doyle, 1992); however, their effect on human colonization is poorly understood (Schoeni and Wong, 1994). We aimed to investigate the inhibitory effect of four new human isolated probiotic strains in regard to their ability to prevent in vitro infection of isolated chicken intestinal cells and also to test their potential in reducing the pathogen load in poultry (Baffoni et al., 2012).
Until now only a few studies have described in detail the possible role of probiotics in preventing C. jejuni colonization of poultry, with most of these probiotic strains originating from commercial probiotic products without a previous in vitro examination of their activity (Morishita et al., 1997; Chaveerach et al., 2004). It has been previously shown that C. jejuni is able to invade chicken intestinal primary cells in vitro (Byrne et al., 2007). In our experiments, the probiotic human isolates used, in combination, had a significant negative effect on the ability of C. jejuni to invade chicken intestinal epithelial cells in vitro. This finding is consistent with previous reported data showing that commercially available probiotics are able to reduce C. jejuni infection of human intestinal cells but to a much lower extent (Alemka et al., 2010).
The efficiency of our probiotics in reducing the invasion of chicken primary cells makes it plausible that they can be cost-effective in reducing the prevalence and colonization of Campylobacter in chicken broilers. Human isolated probiotics have been previously tested to prevent C. jejuni load in poultry, and were effective in preventing mucus colonization (Ganan et al., 2013). Moreover, avian-specific probiotics have also been shown to have a negative effect on colonization and fecal shedding of Campylobacter in broilers (Morishita et al., 1997). Our study went a step further and investigated the pathogen presence in duodenal and cecal mucus but also in the content of these two intestinal compartments. We are able to show that the probiotic human isolates used in this study can significantly reduce the pathogen load in both intestinal content and mucus colonization. Our approach also intended to identify the proper timing for their inclusion in the broilers feed, thus having an advantageous economic impact for the producer. The positive effect was evident either when the probiotics were introduced in the feed immediately after the chickens' arrival from the hatchery or introduced in the last week before slaughter. These results suggest that these probiotic strains can be efficiently used as prophylactics in broilers' feed even if they are introduced in the feed in the last week of the growth cycle. The mechanism involved in this competitive exclusion is largely unknown, but some studies are suggesting that probiotics can cause down-regulation of proteins involved in flagellar protein biosynthesis, with a great impact on their motility and ability to colonize or infect the host (Ding et al., 2005).
One of the possible explanations for their efficiency within the last week of growth could be that a mature gut is more reactive to changes in the microenvironment, being more adaptable to changes in the structure and function of the gut. Following the histomorphological examination of the tissue samples from duodenum and cecum, the addition of probiotics had a significant effect on the appearance and development of the goblet cells, which can explain the decrease in mucus colonization by affecting intestinal mucosa. Their effect on the mucosa has been previously shown in combination with prebiotics in broilers exposed to heat stress (Ashraf et al., 2013).
In conclusion, we have shown that human isolated probiotic bacteria (L. paracasei J.R and L. rhamnosus 15b, L. lactis Y, and L. lactis FOa) are able to efficiently inhibit C. jejuni invasion of chicken intestinal cells, in vitro, and can significantly reduce the pathogen presence within the chicken's intestine in vivo.
Footnotes
Acknowledgments
This work was supported by a grant of the Romanian National Authority for Scientific Research, CNCS–UEFISCDI, project number PN-II-RU-TE-2012-3-0092.
Disclosure Statement
No competing financial interests exist.