
Abstract
Weak organic acids are widely used as preservatives and disinfectants in the food industry. Despite their widespread use, the antimicrobial mode of action of organic acids is still not fully understood. This study investigated the effect of acetic acid on the cell membranes and cellular energy generation of four Salmonella strains. Using a nucleic acid/protein assay, it was established that acetic acid did not cause leakage of intracellular components from the strains. A scanning electron microscopy study further confirmed that membrane disruption was not the antimicrobial mode of action of acetic acid. Some elongated Salmonella cells observed in the micrographs indicated a possibility that acetic acid may inhibit DNA synthesis in the bacterial cells. Using an ATP assay, it was found that at a neutral pH, acetic acid caused cellular energy depletion with an ADP/ATP ratio in the range between 0.48 and 2.63 (p<0.05) that was apparent for the four Salmonella strains. We suggest that this effect was probably due solely to the action of undissociated acid molecules. The antimicrobial effect of acetic acid was better under acidic conditions (ADP/ATP ratio of 5.56±1.27; p<0.05), where the role of both pH and undissociated acid molecules can act together. We concluded that the inhibitory effect of acetic acid is not solely attributable to acidic pH but also to undissociated acid molecules. This finding has implication for the use of acetic acid as an antimicrobial against Salmonella on food products, such as chicken meat, which can buffer its pH.
Introduction
O
Despite their widespread use, the antimicrobial mode of action of organic acids is still not yet fully understood (Mani-López et al., 2012). A number of explanations have been put forward for the antimicrobial activity of organic acids. Organic acids can exist either as charged or uncharged molecules, depending on the protonation state of their acidic group and as determined by the pKa of the acid and the pH of the environment (Brul and Coote, 1999). The antimicrobial effect of organic acids may be due to the lipid permeability of the undissociated molecules, which can freely diffuse into the cytoplasm. As these molecules diffuse across the membrane and encounter a neutral environment in the cytoplasm, dissociation of the acid into anions and free protons is favored (Eklund, 1983, 1985; Salmond et al., 1984). The accumulation of acid anions in the cytoplasm is a possible mode of antimicrobial action (Cherrington et al., 1991). In addition, due to the pH shifting from the internal pH range of optimal enzymatic activity, protein and nucleic acid synthesis are adversely affected by the presence of organic acids at an elevated level (Cherrington et al., 1990). Russell (1992) also reported that the accumulation of the acid anion hinders the proton motive force and inhibits the microbes' ability to re-alkalinize the cytoplasm. The accumulation of high levels of organic acids anions in the cytoplasm may interfere with essential metabolic reactions. In order to maintain the neutral pH and to restore homeostasis, bacterial cells induce an energetically expensive stress response, which may result in depletion of cellular energy (Davidson, 2001; Ricke, 2003). Roth and Keenan (1971) reported that lactic acid can cause sublethal injury to Escherichia coli, and indirect evidence indicated that the injury involved the disruption of the lipopolysaccharide (LPS) layer. A later study (Alakomi et al., 2000) reported that lactic acid permeabilized the membrane of Gram-negative cells and released the LPS from outer membrane. Citric and malic acids were found to chelate (or intercalate with) the outer membrane of Salmonella (Helander and Mattila-Sandholm, 2000).
Acetic acid is one of the most commonly used organic acids and has generally recognized as safe (GRAS) status for use in food and meat products (FDA, 2013). This includes its use in marinades for chicken in the form of vinegar and as a disinfectant on chicken carcasses (Liao et al., 2003; Over et al., 2009; Doležalová et al., 2010). A previous study (Tan et al., 2014) suggested that the inhibitory activity of acetic acid against Salmonella on chicken was due to other factors than its acidic pH. Here we report on an investigation into the mode of action of acetic acid against Salmonella with respect to its ability to disrupt cellular membranes and deplete cellular energy.
Materials and Methods
Preparation of Salmonella cell suspension and acid treatments
Salmonella enterica serovar Typhimurium (ATCC 33062 isolated from chicken tissue and ATCC 14028, a laboratory strain of human origin) and Salmonella enterica serovar Enteritidis (ATCC 13076 of human origin and ATCC 49216 of animal origin) were used in the study, and cell suspensions were prepared according to Tan et al. (2014). Briefly, each of the Salmonella strains was streaked on tryptic soy agar (Oxoid, UK) from pure glycerol stock cultures and incubated for 24 h at 37°C. A colony of each Salmonella strain was inoculated into 10 mL of tryptic soy broth (Oxoid, UK) and incubated for another 24 h at 37°C. The broth cultures were centrifuged at 5000×g (Hettich, USA) for 10 min at 4°C. The supernatant was discarded and the pellet was resuspended in 10 mL of 1× phosphate-buffered saline (PBS; 2.7 mM KCl, 10 mM Na2HPO4, 17 mM KH2PO4, 137 mM NaCl; 1st-Base, Singapore) and used immediately.
Three sets of acid treatments were prepared for experiments. For the first set of treatments, a 5-M stock solution of acetic acid (Merck, Germany) was diluted in 1× PBS (1st-Base) to a concentration of 0.3 M (pH 3.8) and 0.5 M (pH 3.25). These concentrations were used to simulate the use of acetic acid in the form of vinegar in marinades (≈pH 4) and were established to be bactericidal to Salmonella in a previous study (Tan et al., 2014). For the second set of treatments, solutions of 0.3 M and 0.5 M acetic acid were prepared as above and the pH was adjusted to pH 6.4 using a few drops of concentrated sodium hydroxide (NaOH; Merck, Germany) to maintain the concentrations of the solutions. This was done to simulate the pH environment that Salmonella attached to chicken meat surfaces are exposed to as reported in a previous study (Tan et al., 2014). A third acid treatment consisted of 1 M HCl adjusted to pH 3.8 and pH 3.25 using concentrated NaOH to allow comparison between the effect of organic and inorganic acid at the same pH. Ampicillin-treated cells (United States Biological, USA) were used as a positive control and untreated cells were used as a negative control. For experiments, a 5-mL aliquot of each acid treatment was added to a 5-mL Salmonella inoculum and incubated at 4°C for 24 h.
Determination of leakage of intracellular components
The integrity of the cellular membrane was determined initially and after 24-h incubation at 4°C treated with 0.3M (pH 3.8) acetic acid and 0.5 M (pH 3.25) acetic acid according to Dykes (1999) with modifications. Aliquots of the cells were centrifuged for 10 min at 10,000×g, 4°C. The absorbance of the cell-free supernatant fluid was read at 260 nm (which represented the leakage of nucleic acids) and 280 nm (which represented the leakage of proteins) with a UV-VIS Spectrophotometer (Shimadzu, Japan) using 1× PBS as a reference and ampicillin (United States Biological, US) as a positive control.
Scanning electron microscopy
Glass slides (Thermo Scientific, USA) were broken into approximately 5×5-mm sections and placed in a sterile petri dish (Jatikhas, Malaysia). Salmonella inoculum was prepared and treated with 0.5 M (pH 3.25) acetic acid. Specifically, 100 μL of untreated Salmonella inoculum and a 100-μL acetic acid–treated Salmonella inoculum were allowed to attach on the glass slides for 60 min at room temperature. All sample slides were rinsed in 1× PBS, air dried, and chemically fixed with 2.5% glutaraldehyde (R&M Chemicals, USA) for 40 min and gently washed twice again in PBS. Dehydration of the samples was conducted in a series of ethanol concentrations (20, 40, 60, 80, and 100% vol/vol ethanol in water; 20 min in each concentration) and air dried in the laminar flow. Samples were gold-sputtered using a sputter coater (Q150RS; Quorum, UK) and viewed under a scanning electron microscope (SEM) (S-3400N; Hitachi, Japan). The area, length, and perimeter of acetic acid–treated and controls cells under SEM were analyzed using ImageJ 1.22d (The National Institutes of Health, USA).
Measurement of the effect of acetic acid on the intracellular ATP/ADP ratio
All three sets of acid treatments as described above were used on Salmonella cells in this assay. ATP and ADP were measured using the ADP/ATP Ratio Bioluminescent Assay Kit (ab65313; Abcam, UK) according to the manufacturer's instruction. Briefly, 100 μL of reaction mix (10 μL of ATP Monitoring Enzyme and 90 μL of Nucleotide Releasing Buffer, Abcam, UK) was added to the appropriate wells of a 96-well white plate (Greiner Bio-one, Germany) and left for 2 h at room temperature to eliminate ATP contamination. Background luminescence (Data A) was then read using an Infinite® 200 PRO microplate reader (Tecan, Switzerland). The treated and untreated Salmonella cultures prepared as above were centrifuged at 10,000×g for 10 min and resuspended in 1× PBS to remove any interfering substances or acid from the cells. Ten microliters of cell suspension was transferred to a 96-well white plate and luminescence (Data B) was read. The level of ATP was calculated by subtracting the background luminescence from the ATP luminescence reading. To measure ADP levels, 1 μL of ADP Converting Enzyme (Abcam, UK) was added to the wells and luminescence (Data C) was read again after 2 min. The ADP/ATP ratio was calculated using the equation provided by the manufacturer:
Statistical analyses
All experiments were conducted in triplicates with independent cultures. Differences in the leakage of intracellular components of the respective controls and acetic acid treatments were determined using one-way analysis of variance (ANOVA). A post-hoc Tukey's test was used for pairwise comparison to determine the significant difference between samples at p<0.05.The differences of the ADP/ATP ratios between acid treatments and the four Salmonella strains were analyzed using a two-way ANOVA with a Tukey's multiple-comparison test. All tests were conducted using SPSS software version 16.0 (PASW Statistics 20; SPSS Inc.).
Results and Discussion
Determination of leakage of intracellular components
The results of the cellular leakage experiments for all four strains of Salmonella treated with 0.3 M and 0.5 M acetic acid were not significantly different and therefore the data for Salmonella Typhimurium ATCC 33062 are presented as an example in Figure 1. The cells treated with ampicillin (positive control) had substantial leakage of intracellular nucleic acids (260 nm) and proteins (280 nm). Ampicillin belongs to the β-lactams antibiotics family, which act upon the penicillin-binding protein, leading to the weakening of the cell wall and ultimately cell death (Anderson et al., 2012). The level of nucleic acid and protein after treatment of cells with 0.3 M and 0.5 M acetic acid was not different (p>0.05) from untreated (negative control) cells, indicating that these treatments did not result in leakage of intracellular components. By contrast, lactic acid is able to cause LPS release in Pseudomonas, E. coli, and Salmonella Typhimurium by disrupting the outer membrane of the cells (Alakomi et al., 2000). In addition, sorbic acid was found to cause membrane disruption in yeast as reported by Stratford and Anslow (1998) and Bracey et al. (1998). These studies hypothesized that the undissociated acid molecules of both lactic and sorbic acid are responsible for membrane disruption. The pKa values of lactic acid, sorbic acid, and acetic acid are 3.86, 4.75, and 4.74, respectively (Narendranath et al., 2001). At a given acidic pH, there is more undissociated acetic acid and sorbic acid present than in an equal concentration of lactic acid due to their higher pKa values. Our results showed that acetic acid at a high concentration of undissociated acids did not cause cellular membrane disruption. This indicates that different organic acids may inhibit bacteria by different mechanisms. For example, Maiorella et al. (1983) showed that acetic acid can interfere with yeast metabolism, which increased in the ATP requirement for cell maintenance, whereas lactic acid can inhibit growth of yeast by disrupting the cell membrane and releasing LPS from cells (Alakomi et al., 2000; Narendranath et al., 2001). Further work needs to be done to verify the differences in the antimicrobial mode of action of different organic acids. We concluded that acetic acid is unlikely to disrupt the cell membrane of Salmonella, as no leakage of intracellular component was apparent.
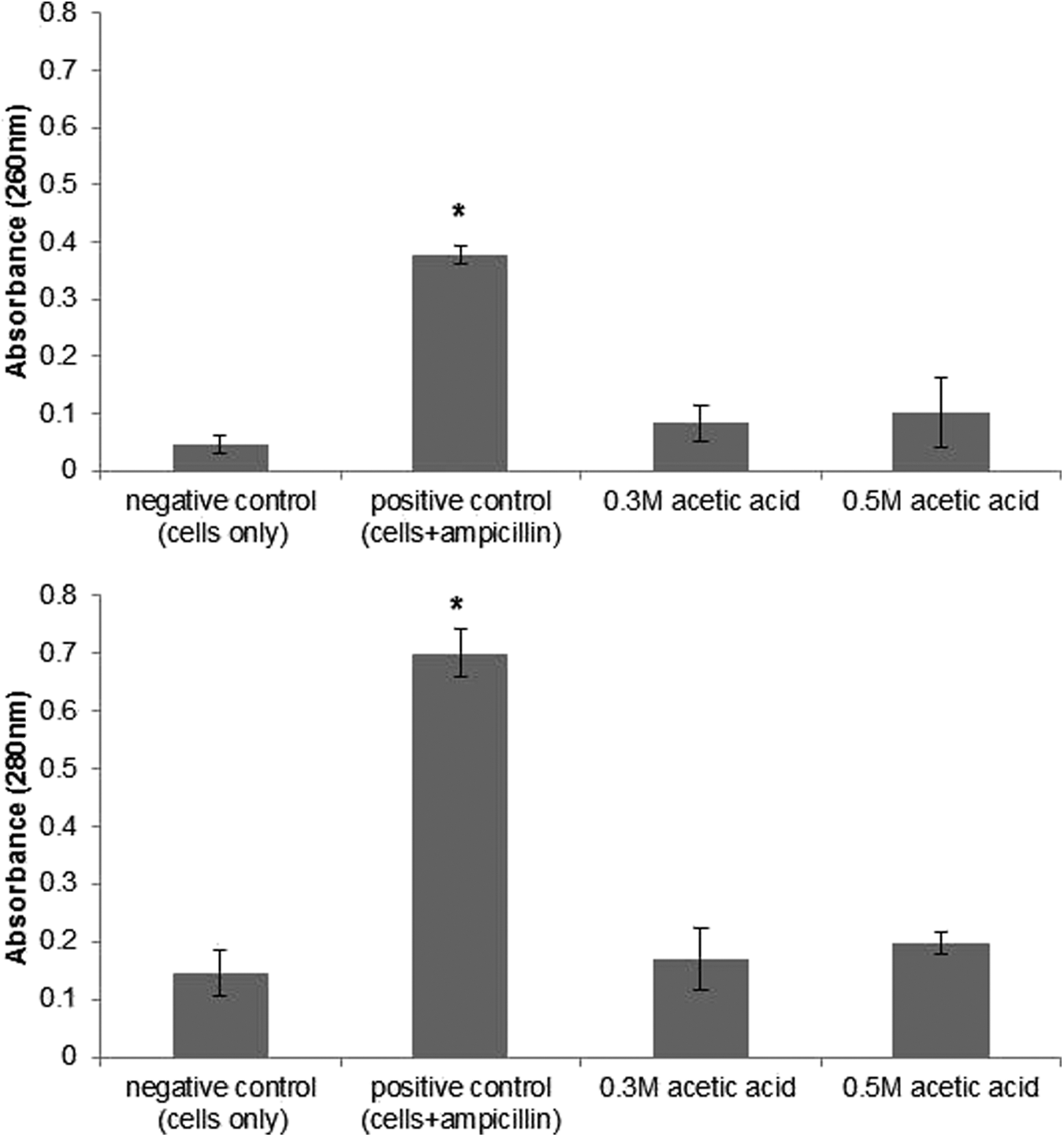
The leakage of intracellular components of Salmonella Typhimurium ATCC 33062 at 260 nm and 280 nm treated with 0.3 M and 0.5 M acetic acid. Results are presented as mean±SD where n=3. Asterisk indicates a significant difference between positive control and acetic acid treatments. Error bars represent standard deviation.
SEM of acetic acid–treated Salmonella cells
The SEM micrographs in Figure 2 show the cellular morphology of acetic acid–treated and untreated Salmonella cells. Some cell shrinkage was observed due to the alcohol dehydration process during the SEM preparation techniques (Sutton et al., 1993). Figure 2B shows no acid-induced disruption of outer cell membrane or cell wall after being treated with 0.5 M acetic acid as compared to an untreated cell (Fig. 2A). This further confirms that acetic acid does not cause cell wall or cell membrane disruption in Salmonella. Our study is in agreement with that of Thompson and Hinton (1996), who reported that there was no formic and propionic acid–induced damage on cytoplasmic membrane of Salmonella Enteritidis in a transmission electron microscopy study. The mean area, length, and perimeter of untreated Salmonella cells and acetic acid–treated cells are presented in Table 1. All these measures increased significantly (p<0.05) for cells treated with acetic acid. Individual cells with a length of up to 7.35 μm were apparent in acetic acid–treated but not in untreated samples (Fig. 2C, D, E, and F). The increase in the length of acid-treated cells may be because of the inhibition of DNA synthesis, while other macromolecules, such as RNA and proteins, continue to synthesize, resulting in increased cell length without cell division (Thompson and Hinton, 1996). Other foodborne pathogens such as Listeria monocytogenes, E. coli, and Bacillus cereus have been reported to produce filaments after prolonged exposure to pH and other stresses (Bereksi et al., 2002; Jones et al., 2003; Jydegaard-Axelsen et al., 2005; den Bensten et al., 2009). This feature is of concern because filamentous cells can rapidly divide into multiple daughter cells if nonstressful conditions return. In addition, filament formation can result in an underestimation of viable bacterial count because filamentous cells form a single colony on plated solid agar medium (Giotis et al., 2007; Jones et al., 2013). This may lead to a food safety risk for consumers.

Scanning electron microscope images of Salmonella Typhimurium ATCC 33062 on glass slides.
Data are presented as mean±SD where n=8. Different lowercase letters indicate significant differences between treated cells and untreated cells (within columns) at a p<0.05 level.
Measurement of the effect of acetic acid on the intracellular ATP/ADP ratio
As shown in Table 2, of all the treatments applied, 0.5 M acetic acid resulted in the highest ADP/ATP ratio (p<0.05) followed by 0.3 M acetic acid (p<0.05), pH 6.4 adjusted 0.5 M acetic acid (p<0.05), pH 6.4 adjusted 0.3 M acetic acid (p<0.05), and lastly HCl treatments at pH 3.8 and pH 3.25 (p<0.05). There was no difference in this ratio for HCl treatment cells at pH 3.8 and pH 3.25 and the negative control. This indicates that the HCl treatments at pH 3.8 and pH 3.25 were not causing energy exhaustion in Salmonella cells. Salmonella can survive from pH 3 to pH 9 if the pH value is purely due to the presence of HCl and NaOH (D'Aoust and Maurer, 2007; Tan et al., 2014). On the other hand, energy exhaustion was observed in cells treated with 0.3 M acetic acid and 0.5 M acetic acid at the same pH. This indicates that the energy exhaustion of cells is not merely caused by the acidic pH but also by the undissociated acetic acid molecules that easily diffuse into the cell cytoplasm and dissociate into anions. This in turn leads to an accumulation of high levels of toxic anions and interference with essential metabolic reactions (Ricke, 2003).
Data are presented as mean±SD where n=3. Different uppercase letters indicate significant differences between acid treatments (within columns) at a p<0.05 level. Different lowercase letters indicate significant difference between bacterial strains (within rows) at a p<0.05 level.
Table 2 also shows that 0.5 M acetic acid at pH 6.4 induces a higher ADP/ATP ratio (for example, a ratio of 1.85±0.42 for Salmonella Typhimurium ATCC 14028; p<0.05) as compared to 0.3 M acetic acid at pH 6.4 (0.75±0.19; p<0.05). This may be due to a higher concentration of undissociated acid molecules in the 0.5 M acetic acid at pH 6.4. A higher concentration of undissociated acid molecules results in higher accumulation of acid anions in the cell cytoplasm and causes more energy depletion than a low-concentration acid (Davidson, 2001). The 0.5 M acetic acid with an initial pH 3.25 induced a higher ADP/ATP ratio (Table 2, p<0.05) than the 0.5 M acetic acid at pH 6.4, and this indicates that at the same concentration the acetic acid with a lower pH has a better bactericidal activity as compared to that at a near-neutral pH. As reported by Davidson (2001), pH is considered a primary determinant of effectiveness of organic acids because it affects the concentration of undissociated acid formed. In addition, the uncharged, undissociated acid molecules, which are primarily responsible for the antimicrobial activity, are most prevalent at low pH and therefore organic acids have optimal inhibitory activity at a low pH (Aslim et al., 2005; Nazer et al., 2005). The overall ADP/ATP ratio of Salmonella Typhimurium ATCC 33062 was higher than for the other three strains (p<0.05; Table 2), whereas Salmonella Typhimurium ATCC 14028 had a lower overall ADP/ATP ratio (p<0.05) as compared to other strains. This indicates that Salmonella Typhimurium ATCC 33062 is more sensitive to and Salmonella Typhimurium ATCC 14028 is more resistant to acetic acid treatments than other strains. We do not know what caused the differences between strains and this needs to be further investigated. Although strain variations were apparent in the study, 0.3 M and 0.5 M acetic acid at their respective initial pH had a better inhibitory effect as compared to acetic acid at pH 6.4 and HCl treatments. We conclude that the undissociated acid molecules can cause energy depletion in Salmonella in the absence of a pH influence. However, the antimicrobial effect of acetic acid was better in the presence of both acidic pH and undissociated acid molecules than only one of them. This suggests that marinades or other treatments with acetic acid should be effective against Salmonella to some extent on chicken meat even though the buffering effect of meat neutralizes the acidic pH. It also implies that acetic acid could be applied as an antimicrobial against Salmonella on food products for which a neutral pH is an intrinsic property and necessary for its organoleptic properties.
Conclusions
The antimicrobial mode of action of acetic acid includes factors other than its acidic pH. Acetic acid did not cause leakage of intracellular components and the SEM study further confirmed that membrane disruption is not the antimicrobial mode of action of acetic acid. Some elongated Salmonella cells observed in the micrographs show a possibility that acetic acid may inhibit DNA synthesis in the bacterial cells. At a neutral pH, acetic acid caused energy depletion in cells, probably due to the undissociated acid molecules. The antimicrobial effect of acetic acid was better under acidic conditions where the role of low pH and undissociated acid molecules may act together. We concluded that the inhibitory effect of acetic acid is not solely attributable to the low pH but also to the undissociated acid molecules.
Footnotes
Acknowledgments
We acknowledge Ms. Amreeta Sarjit for her technical assistance.
Disclosure Statement
No competing financial interests exist.