
Abstract
β-Lactamases and extended-spectrum β-lactamases (ESBLs) producing pathogenic bacteria were widely studied previously in China, but were seldom focused on foodborne Salmonella. In this study, an investigation concerning β-lactamases and ESBLs producing Salmonella recovered from retail raw chickens was performed. Sixty of 699 foodborne Salmonella isolates were detected as β-lactamases and ESBLs-producing ones that covered 12 Salmonella serotypes and exhibited different pulsed-field gel electrophoresis genotypes. Forty-four of 60 β-lactamases and ESBLs-producing strains were simultaneously resistant to ampicillin, amoxicillin/clavulanic acid, ceftiofur, ceftriaxone, and cefoxitin. The most commonly detected β-lactamases and ESBLs-encoding gene was bla TEM-1 (n=44), followed by bla OXA-1 (n=38), bla CMY-2 (n=29), bla PSE-1-like (n=1), bla CTX-M-3 (n=16), and bla CTX-M-15 (n=1), respectively. Fourteen, 24, 21, and 1 isolates were detected simultaneously positive for 1, 2, 3, and 4 of the detected β-lactamases and ESBLs-encoding genes, respectively. A Salmonella strain simultaneously co-carrying bla TEM-1, bla OXA-1, bla CMY-2, and bla CTX-M-3 was first reported in the present study. Amino acid substitution of Trp244Cys/His247Leu was detected in PSE-1, Val218Asp in CMY-2, and Asp242Gly in CTX-M-15 enzymes, respectively. A difference was found among the amino acid sequences of the detected OXA-1, CMY-2, CTX-M, PSE-1, and TEM-1. The results demonstrated that β-lactamases and ESBLs were emerging and prevalent in foodborne Salmonella.
Introduction
S
Materials and Methods
Bacterial isolates
Six hundred ninety-nine Salmonella isolates were recovered from 1152 retail raw chicken carcasses sampled from 192 supermarkets and 96 wet markets in 6 provinces (Shaanxi, Sichuan, Henan, Guangdong, Guangxi, and Fujian) and 2 National cities (Beijing and Shanghai) of China from March to December in 2010. Among 699 Salmonella isolates, 86 were from Henan province, 65 from Shaanxi, 69 from Sichuan, 77 from Fujian, 100 from Guangxi, 117 from Guangdong, 116 from Beijing, and 69 from Shanghai city, respectively. During sampling, three cities including a capital city were selected in each province and three counties were selected in each National city. In each city (county), four large supermarkets, four small supermarkets, and four wet markets (which include butcher shops for live poultry and fish stalls as well as other stands for fruits and vegetables) were visited for sample collection. In each marketplace, four samples were collected randomly. Salmonella was isolated and identified as previously described in the United States Department of Agriculture Food Safety Inspection Service protocol for isolation and identification of Salmonella from Meat, Poultry, and Egg Products (USDA FSIS, 2014). The isolates were serotyped using O and H hyperimmune seras (Statens Serum Institut, Artilerivej, Denmark) and slide agglutination method according to the manufacturer's instructions. Salmonella isolates used in this study were different from those that were recovered in 2007–2008 in our previous studies (Yang et al., 2014).
Antimicrobial susceptibility test
The minimum inhibitory concentrations (MICs) of the antimicrobial agents to the isolates were tested using the agar dilution method, and the susceptibility was determined according to the guidelines of the Clinical and Laboratory Standards Institute (CLSI, 2010) except streptomycin (US FDA, 2014). The following antimicrobial agents were tested: amikacin (8–64 μg/mL), gentamicin (2–16 μg/mL), kanamycin (4–64 μg/mL), streptomycin (32–64 μg/mL), ampicillin (4–32 μg/mL), amoxicillin/clavulanic acid (4/2–32/16 μg/mL), ceftiofur (4–64 μg/mL), ceftriaxone (2–64 μg/mL), cefoxitin (4–32 μg/mL), sulfisoxazole (64–512 μg/mL), trimethoprim-sulfamethoxazole (0.5/9.5–4/76 μg/mL), chloramphenicol (4–32 μg/mL), ciprofloxacin (1–8 μg/mL), nalidixic acid (4–32 μg/mL), levofloxacin (1–8 μg/mL), gatifloxacin (1–8 μg/mL), and tetracycline (4–16 μg/mL). E. coli ATCC 25922 and ATCC35218, and Enterococcus faecalis ATCC 29212 were used as quality-control organisms in MICs determinations.
Double disk synergy test
β-Lactamase and ESBLs-producing isolates were screened according to the double-disk synergy test method recommended by the CLSI combined with antimicrobial disks ([cefotaxime 30 μg, cefotaxime-clavulanate 30 μg/10 μg, ceftazidime 30 μg, and ceftazidime-clavulanate 30 μg/10 μg]) according to the manufacturer's instructions (Rosco, Taastrup, Demark) (Clinical and Laboratory Standards Institute, 2010). Phenotypic presence of β-lactamases of the isolates was determined by detecting diameter enhancement of inhibition zone of the clavulanate disk and corresponding β-lactam antimicrobial disk. If the enhancement value was >5 mm, the isolate was considered to be presumptive β-lactamases producers (CLSI, 2010).
DNA isolation, polymerase chain reaction (PCR), and gene analysis
DNA templates were prepared from putative β-lactamases-producing isolates by heating a bacterial suspension at 100°C for 10 min and centrifuging at 13200 rpm. β-lactamases and ESBLs-encoding genes including bla TEM, bla CMY, bla CTX-M, bla OXA, bla ACC, bla PSE, bla SHV, bla VEB, bla PER, and bla GES were amplified by PCR and sequenced for further confirmation. PCR conditions were conducted as described previously (Archambault et al., 2006). The primers for PCR amplification and sequencing are listed in Table 1. After amplification, DNAs were sequenced and aligned using GenBank online BLAST software (GenBank, 2014).
Pulsed-field gel electrophoresis (PFGE)
ESBLs-producing Salmonella isolates were subtyped by PFGE to determine their genetic relatedness according to the protocol recommended by the CDC (Ribot et al., 2006). Briefly, agarose (SeaKem® Gold Agarose, Lonza, Rockland, ME)-embedded isolates were lysed and DNA was digested with 50 U of XbaI (TaKaRa) for 1.5–2 h in a water bath at 37°C. The restricted DNA fragments were separated in 0.5×Tris-borate-EDTA (Xi'an Sanpu Chemical Co, Ltd., Xi'an, Shaanxi, China) buffer at 14°C for 19 h using a Chef-Mapper electrophoresis system (Bio-Rad, Hercules, CA) with pulse times of 2.16–63.8 s. Salmonella serotype Braenderup H9812 was used as standard control strain. The gels were stained with ethidium bromide at the concentration of 1 μg/mL, and DNA bands were visualized with UV transillumination (Bio-Rad). PFGE results were analyzed using the BioNumerics Software (Version 3.0; Applied-Maths, Kortrijk, Belgium) manually.
Homology of the detected β-lactamases and ESBLs
Homology of the detected β-lactamases and ESBLs were determined by the neighbor-joining method of Molecular Evolutionary Genetics Analysis software (MEGA5.1) based on amino acid sequences of CMY-2, CTX-M-3, CTX-M-15, TEM-1, PSE-1-like, and OXA-1.
Results
Sixty-four Salmonella serotypes were detected among 699 Salmonella isolates. Twelve serotypes were detected in 60 β-lactamases and ESBLs-producing isolates, among which, the most prevalent serotype was Salmonella Indiana (28/60, 46.7%), and the less prevalent ones were Salmonella Thompson (8/60, 13.3%), Salmonella Edinburg (6/60, 10.0%), Salmonella Typhimurium (4/60, 6.7%) and Salmonella Enteritidis (4/60, 6.7%) (Fig. 1). Based on 80% similarity, 5 clusters (assigned as A, B, C, D, E, respectively) and 4 single genotypic profiles were found among 60 β-lactamases and ESBLs producing isolates. Except isolates 768D and 926T, 674D and 974T, 185T and 68D, which were recovered from different province and/or marketplaces, and isolates 759T and 778T, 1073D and 993T, 819D and 879D, which were from same province and/or kind of market place, and showed 100% genotypic similarity, the majority of isolates exhibited different PFGE genotypes with different DNA profiles after their genomic DNA was digested by XbaI and separated by PFGE (Fig. 1).
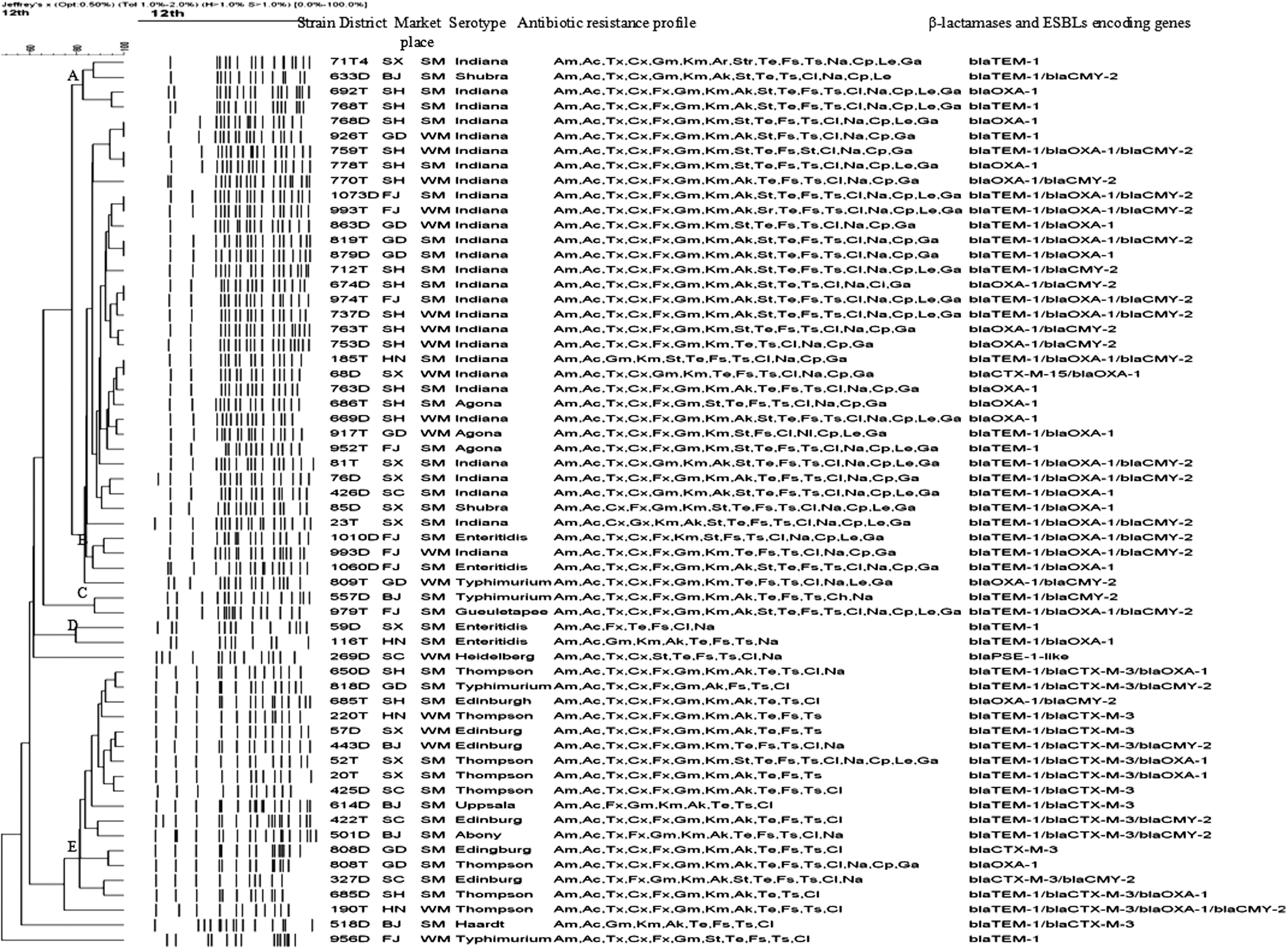
Pulsed-field gel electrophoresis dendrogram, showing district, marketplace, serotype, antibiotic resistance profile, β-lactamases, and extended-spectrum β-lactamases (ESBLs) encoding genes of the 60 β-lactamases and ESBLs-producing Salmonella isolates. SX, Shaanxi Province; BJ, Beijing City; SH, Shanghai City; GD, Guangdong Province; FJ, Fujian Province; HN, Henan Province; SC, Sichuan Province. SM, Supermarket; WM, Wet market. Am, ampicillin; Ac, amoxicillin/clavulanic acid; Tx, ceftriaxone; Cx, ceftiofur; Fx, cefoxitin; Gm, gentamicin; Km, kanamycin; Ak, amikacin; St, streptomycin; Te, tetracycline; Fs, sulfisoxazole; Ts, trimethoprim-sulfamethoxazole; Cl, chloramphenicol; Na, nalidixic acid; Cp, ciprofloxacin; Le, levofloxacin; Ga, gatifloxacin.
Among 60 β-lactamases and ESBLs-producing Salmonella isolates, 9 (18.3%) were simultaneously resistant to 17 antimicrobial agents with the same antibiotic resistance profile of AmAcTxCxFxGmKmArStrTeFsTsClNaCpLeGa; 9 isolates were co-resistant to 16 antibiotics with the dominant antibiotic resistance profile of AmAcTxCxFxGmKmStTeFsTsClNaCpLeGa (n=4); and 11 were resistant to 15 antibiotics with the dominant profile of AmAcTxCxGmKmAkStrTeFsTsNaCpLeGa (n=7). Multidrug resistance (MDR) could also be found in 6 isolates that were co-resistant to 14 antibiotics, 4 to 13 antibiotics, 9 to 12 antibiotics, and 6 to 11 antibiotics; however, no predominant antibiotic resistance profiles could be detected (Fig. 1). All β-lactamases and ESBLs-producing isolates were resistant to ampicillin and amoxicillin/clavulanic acid. Fifty-three (88.3%), 52 (86.7%), and 50 (83.3%) of the β-lactamases and ESBLs-producing strains could resist ceftiofur, ceftriaxone, and cefoxitin, respectively. Forty-four (73.3%) isolates were detected simultaneously as resistant to 5 tested β-lactam antibiotics and 10 (16.7%) were resistant to 4 of the 5 β-lactam antibiotics (Fig. 1).
Six β-lactamases and ESBLs (CMY-2, TEM-1, OXA-1, PSE-1, CTX-M-3, and CTX-M-15) encoding genes were detected in 60 β-lactamases and ESBLs-producing Salmonella isolates after PCR and DNA sequencing confirmation. Forty-four of 60 β-lactamases and ESBLs-producing isolates were detected carrying bla TEM, which was the most prevalent gene, followed by bla OXA (n=38), bla CMY-2 (n=29), bla CTX-M (n=17) and bla PSE (n=1), respectively (Table 2).
Forty-four bla TEM genes were all assigned as bla TEM-1 after their DNA sequences were aligned using GenBank online BLAST software (GenBank), and bla OXA was assigned as bla OXA-1, bla CMY as bla CMY-2, bla PSE as bla PSE-1 variant (designated as bla PSE-1-like), bla CTX-M as bla CTX-M-3 and bla CTX-M-15, respectively. Fourteen strains were detected positive for one β-lactamases and/or ESBLs-encoding genes including bla TEM-1 (n=6), bla OXA-1 (n=6), bla CTX-M-3 (n=1), and bla PSE-1like (n=1). Twenty-four strains were detected simultaneously carrying two β-lactamases and/or ESBLs-encoding genes (Table 2). One strain was simultaneously carrying four β-lactamases and/or ESBLs-encoding genes of bla TEM-1 /bla CTX-M-3 /bla OXA-1 /bla CMY-2 (Table 2).
The amino acid sequence of the 44 detected TEM-1 was all identical to that (accession No. JN003853) in the GenBank as well as the 38 detected OXA-1 to that (accession No. EU162133.1) in the GenBank. Twenty-eight of the 29 detected CMY-2 were identical to that (accession No. EU752483.1) in the GenBank, except one CMY-2 was detected with amino acid substitution of Val218Asp. Salmonella isolate 269D was detected harboring PSE-1-like β-lactamase, which differed from PSE-1 (accession No. JX491633.1) in the GenBank with amino acid substitutions of Trp244Cys and His247Leu. Asp242Gly was detected between CTX-M-15 and CTX-M-3; it was this variation that changed CTX-M-15 to CTX-M-3.
Phylogenetic analysis indicated a difference could be found among the detected OXA-1, CMY-2, CTX-M, TEM-1, and PSE-1. PSE-1 and TEM-1 enzymes presented relatively high homology as well as CMY-2 and CTX-M (Fig. 2).
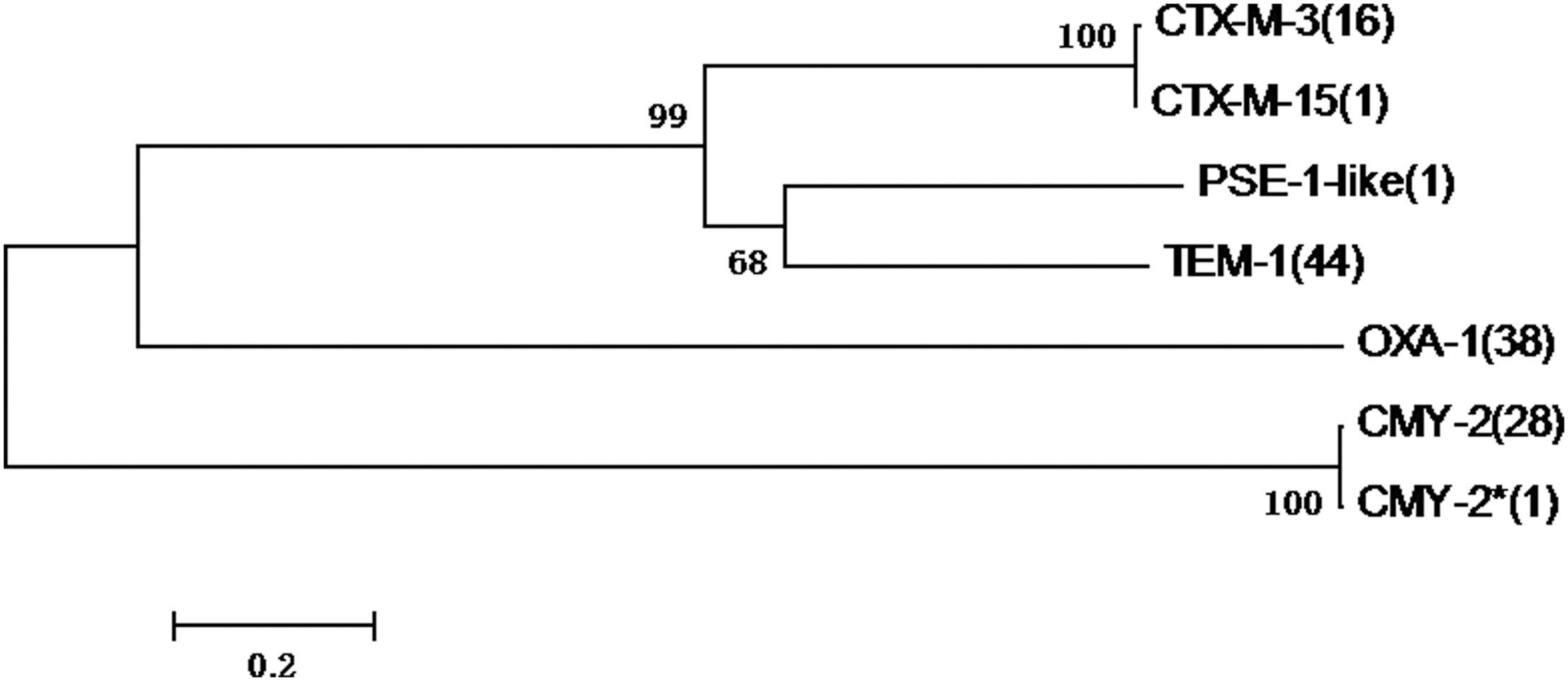
Homology of β-lactamases of TEM-1, CMY-2, OXA-1, PSE-1, and CTX-M-type. The number in the bracket indicates the number of isolates that carrying the corresponding β-lactamases and extended-spectrum β-lactamases.
Discussion
Compared to our previous study (Yang et al., 2014), six β-lactamases and ESBLs (TEM-1, CMY-2, OXA-1, PSE-1, CTX-M-3, and CTX-M-15) encoding genes were detected from ESBLs-producing Salmonella isolates in this study, and the strains were identified simultaneously harboring one or more β-lactamases and ESBLs-encoding genes. Co-carriage of bla CTX-M-2 and bla SHV-5 has been identified in a few Klebsiella spp., and the isolate that carried both bla CTX-M-14 and bla SHV-12 was reported previously in Canada (Mulvey et al., 2004). In the Netherlands, one Salmonella Virchow strain harboring bla CTX-M-2 was also found to carry bla SHV-2 (Hasman et al., 2005). Although pathogenic isolates carrying more than one β-lactamases-encoding genes have been reported previously, they were not frequently being identified (Archambault et al., 2006). In the current study, 14 Salmonella isolates were positive for at least 1 of the 7 detected genes, 24 for 2 and 21 for 3 of the genes, and 1 isolate was detected carrying 4 genes simultaneously, which was not in agreement with the previous studies (Yang et al., 2014). The reasons are partially due to use of different antibiotics, bacteria transmission, and prevalence of β-lactamases-encoding genes in different districts, foods, and environment settings. As far as we know, pathogens carrying three β-lactamases-encoding genes simultaneously is rare, except one Salmonella containing bla CTX-M-15, bla SHV, and bla TEM-1H at the same time was reported previously (Archambault et al., 2006). Co-carriage of bla TEM-1, bla OXA-1, bla CMY-2, and bla CTX-M-3 in Salmonella isolate was first reported in this study. Furthermore, foodborne Salmonella that was positive for PSE type β-lactamase was rarely reported previously (Li et al., 2007; Liebana et al., 2013), and detection of the PSE-1 variant in chickenborne Salmonella in the study should be treated seriously.
The majority of studies concerning β-lactamases and/or ESBLs were conducted clinically; the samples were mainly collected from urine, wounds, abscesses, blood and respiratory tract of patients (Leverstein-van Hall et al., 2011). It is commonly considered that antibiotic-resistant isolates mainly emerged from the abuse of antibiotics in clinics (Li et al., 2013). However, in recent decades, some Gram-negative bacteria resistant to antibiotics have been widely detected in food-producing animals and retail meat (Doi et al., 2009; Mesa et al., 2006; Keelara et al., 2014a). It has been reported that the prevalence of ESBL-producing E. coli from the gastrointestinal tract of food-producing animal increased from 3% in 2003 to 15% in 2008, and in 2009 ESBLs-producing isolates were detected as 100% in all 26 broiler farms investigated (Leverstein-van Hall et al., 2011). Thus, the poultry industry has been considered another reservoir of β-lactamases-producing Gram-negative bacteria that might infect humans by consumption of contaminated retail meat (Pires, 2013). In one study, people working in the poultry industry seemed to have a higher risk of exposure to ESBLs-carrying intestinal bacteria (Huijbers et al., 2014). Leverstein–van Hall (2011) suggested the transmission of ESBL-producing E. coli from animals to humans was most likely through the food chain. Therefore, the large amount of Salmonella isolates carrying β-lactamases or ESBLs detected in retail chicken in this study suggests that β-lactamases-producing Salmonella perhaps can transmit from poultry to humans through touching or consumption of retail meat.
Previous studies indicated the following: (1) TEM-1 β-lactamases can inactivate penicillins and early cephalosporins; (2) CMY-2 is a plasmid-mediated ampC-like enzyme that can destroy β-lactam antibiotics and spread horizontally to different Salmonella serotypes (Lindsey et al., 2011; Keelara et al., 2014b); (3) PSE-1 can readily hydrolyze carbenicillin and oxacillin; (4) CTX-M are classified to ESBLs that mainly compromise extended-spectrum cephalosporins and aztreonam; (5) OXA-1 belongs to carbapenemases with the ability to hydrolyze carbapenem antibiotics; and (6) OXA-related enzyme-encoding genes may be carried in multidrug-resistant plasmids (Huovinen et al., 1988; Bush et al., 1995; Zaidi et al., 2007). In this study, 60 Salmonella isolates were detected carrying 1 or more β-lactamases and ESBLs-encoding genes, which means the potential of disseminating β-lactam and carbapenem resistance in the food chain would be a serious problem, although most studies on β-lactamases and ESBLs focused on clinical- and animal-derived bacteria (Karim et al., 2001; Archambault et al., 2006).
β-Lactamases mutated frequently since the first enzyme was identified. As a result, “new” β-lactamases appeared and could inactivate the latest antibiotics including extended-spectrum cephalosprins and carbapenems (Bush et al., 1995). Among β-lactamases, CTX-M-14 was identified as the predominant ESBLs in China (Yu et al., 2007; Liu et al., 2009; Rao et al., 2014); however, CTX-M-14 was not detected, but CTX-M-3 was detected in this and previous studies (Wu et al., 2006; Cheng et al., 2009; Du et al., 2012), which indicated that perhaps CTX-M-3 was also the prevalent ESBLs in Salmonella in China. Additionally, there were a great number of variations among CTX-M enzymes, which was consistent with the conclusion reported previously (Bush et al., 1995; Archambault et al., 2006).
Although vast TEM variants were identified in our study, few of them currently appeared as dominant ESBLs (Bush et al., 1995). Thus, it may be speculated that TEM enzymes would decrease gradually but CTX-M would be the prevalent ESBLs. A U.S. surveillance indicated that TEM-10, TEM-12, and TEM-26 enzymes were frequently prevalent at first but then almost disappeared, which further strengthened our speculation (Bush, 2008).
β-Lactamases and ESBLs-encoding genes may locate in chromosomes or plasmids, of which plasmids represent one crucial role of MDR emergence and dissemination (Carattoli et al., 2013). By means of PCR-based replicon typing technology, plasmids can be classified into different families including IncI1, IncF, IncHI1, IncHI2, and IncN, and so on. Most of them carry multiple resistance determinants, conferring many types of antibiotic resistance (Carattoli et al., 2011). Previous studies showed that carbapenemases including OXA- and CMY-type β-lactamase are mainly successfully disseminated by plasmids. For example, Poole et al. (2009) found that the bla OXA-48 gene was identified in the IncL/M-type plasmid, and Lindsey et al. (2011) suggested IncA/C plasmids were associated with the spread of the bla CMY-2 gene. In addition, bla TEM and bla SHV were reported typically locating in transferable plasmids, CTX-M-15 was mainly found on the IncI1 and IncF plasmids, and CTX-M-3 was highly related to the IncL/M plasmids (Schultsz et al., 2012; Rodriguez et al., 2014). Since ESBLs of CTX-M-3 and CTX-M-15 detected in this study are two of the increasing reported ones worldwide, and bla CTX-M was found to be located in plasmids more frequently than in chromosomes, and many plasmid families are MDR plasmids that contain multiple resistance determinants (Rodriguez et al., 2014), β-lactamases and ESBLs-encoding genes detected in this study may be potential elements for antimicrobial resistance transmission, especially these isolates containing two or more β-lactamases and ESBLs-encoding genes.
In conclusion, emergence of β-lactamases and ESBLs in foodborne Salmonella is of great public health concern. Management of the food processing and supply chain, combined with prudent use of antibiotics in livestock breeding, are crucial for decreasing β-lactamase-carrying isolates.
Footnotes
Acknowledgments
This research was supported in part by National Natural Science Foundation of China (No. 31171682), the Fundamental Research Funds for the Central Universities of Northwest A&F University, China, and the High Technology Research and Development Program of the Xinjiang Uygur Autonomous Region (201317106).
Disclosure Statement
No competing financial interests exist.