
Abstract
The aim of this work was to investigate the mechanisms underlying the inactivation of Listeria monocytogenes in raw milk under high hydrostatic pressure (HHP). The results showed that a 450-MPa treatment for 5 min could completely eliminate 108 colony-forming units/mL of viable bacteria, as indicted by increased uptake of propidium iodide. Morphological damage to the cell wall, cell membrane, and cytoplasmic components by HHP treatments was observed on scanning and transmission electron micrographs. The protein bands on sodium dodecyl sulfate–polyacrylamide gel electrophoresis gels differed between HHP-treated and untreated L. monocytogenes, in that HHP decreased the protein content and caused partial protein degradation. Therefore, our results indicate that HHP inactivates L. monocytogenes by causing morphological changes in the internal and external cellular structures, as well as through membrane damage, cell wall rupture, and membrane protein degradation.
Introduction
L
High hydrostatic pressure (HHP) is a cold pasteurization technique that is recognized in the food industry as a means to preserve organoleptic flavor. HHP has been approved by the U.S. Food and Drug Administration for use in food-processing operations (Demazeau and Rivalain, 2011). HHP can render harmful microorganisms inactive without detrimentally affecting the natural characteristics of the food, thereby leading to improvements in the overall food quality. In recent years, high-pressure technology has been widely used in the processing of meat products, dairy products, aquatic products, and fruit and vegetable products (Follonier et al., 2012). Recent studies have been reported on the behavior of L. monocytogenes in various food products treated by HHP, such as cheese, smoked salmon, ham, milk, and beef loin (Rastogi et al., 2007). As an in-package cold pasteurization technique, HHP is particularly interesting as a listericidal postprocessing treatment for ready-to-eat meat products.
High pressure brings about a number of changes in the morphology, cell membrane, cell wall, biochemical reactions, and genetic mechanisms of microorganisms (Huang et al., 2014). Each of these changes may contribute to microbial inactivation resulting from HHP treatment. Cell death occurs when multiple parts of a cell are damaged and when the accumulated damage exceeds the cell's capacity for repair. Under some circumstances, the damaged cell can recover if the post-treatment conditions are favorable (Follonier et al., 2012). However, the particular effects of HHP on the cell membrane integrity and ultrastructure of L. monocytogenes have not yet been thoroughly characterized. The objectives of this study were to evaluate morphological damage and cell death of HHP-treated L. monocytogenes cells in raw milk. We analyzed the bacterial viable cell counts, morphological features, and membrane protein changes after treatment with different levels of hydrostatic pressure.
Materials and Methods
Bacterial culture preparation
Listeria monocytogenes BCRC 15354 was obtained from the Bioresource Collection and Research Center in Hsinchu, Taiwan. The strain was cultured in brain heart infusion (BHI) broth (Difco) at 37°C for 24 h, until it reached the stationary phase. The cells at stationary phase were harvested by centrifugation (10 min at 3000×g) and washed once in phosphate-buffered saline (0.01 M phosphate, 0.137 M NaCl, pH 7.3). The washed cells were resuspended in whole milk at a concentration of approximately 1×108 colony-forming units (CFU)/mL. Inoculated milk samples were aseptically transferred to sterile polyethylene bags in 8-mL aliquots and were vacuum packaged immediately.
HHP treatment
HHP treatments were performed using a laboratory high-pressure system with a 300-mL pressure vessel (Foodlab Plunger Press model S-FL-850-9 W; Stansted Fluid Power Ltd., Stansted, UK). The pressure transmission fluid used in this study was water, and the bags containing milk samples were subjected to pressure treatment at 250, 300, 350, 400, and 450 MPa, with a holding time of 5 or 10 min at room temperature (25°C). After processing, the samples were placed in an ice bath and immediately processed for future analysis.
Enumeration of surviving cells
After the pressure treatment, both treated and untreated milk samples were 10-fold serially diluted in 0.1% sterile peptone solution (Becton Dickinson, Cockeysville, MD). Subsequently, 1-mL aliquots of these dilutions were used for viable cell counting by the pour plate method with BHI agar and incubation at 37°C for 24 h. The results were expressed as CFU/mL.
Assessment of cell membrane damage
The LIVE/DEAD® BacLight™ Bacterial Viability Kits were used to evaluate cell membrane damage of L. monocytogenes cells, according to the manufacturer's instructions. Fluorescence was measured at an excitation wavelength of 495 nm and an emission wavelength of 600 nm in a multimode microplate reader (Synergy H1, BioTek Instruments, Inc). The normalized data were plotted as percentages of PI uptake during and after pressure treatment at different pressures.
Electron microscopy
Bacterial cell morphology of L. monocytogenes was observed by scanning and transmission electron microscopy. The sample was fixed and processed following the recommended procedure for preparing specimens (Wang et al., 2013). The samples were subsequently observed under a scanning electron microscope (Hitachi S4700) and a transmission electron microscope (JEM-1400, Japan Electronics Co. Ltd).
Electrophoretic analysis of membrane proteins
Electrophoretic analysis was used to assess the extent of membrane protein damage caused by HHP treatments. Proteins were extracted from each bacterial cell using B-PER protein extraction reagents (Thermo Fisher, Scientific Inc., IL), according to the manufacturer's instructions. The protein extracts were analyzed by performing 12% sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE), and the protein bands on the gel were visualized by staining with Coomassie Brilliant Blue R-250 staining.
Results and Discussion
HHP treatment and bacterial survival
L. monocytogenes was completely inactivated by pressure treatment at 450 MPa for 5 min or 10 min, with the number of viable bacteria decreasing from 108 CFU/mL to zero (Fig. 1). Cell viability was not significantly affected by a 250-MPa treatment. However, the lethality significantly increases with pressure. Treatment with 350 MPa for 5 min reduced the bacterial population by 4.4±0.2 log cycles, whereas the same pressure treatment for 10 min reduced the population by 5.8±0.3 log cycles. Milk was a useful model for investigating the protective effect of foods against pressure, because its chemical composition is well known and many recent studies have added to our knowledge of the effects of HHP treatment on the constituents of milk (Huppertz et al., 2002). Dogan and Erkmen (2004) also reported that D values for L. monocytogenes at 600 MPa were about 2.43 min, 1.52 min, and 0.87 min in whole milk, peach juice, and orange juice, respectively. A higher D value in milk indicates that this food exerts a strong protective effect, compared with buffer systems, when L. monocytogenes is subjected to HHP (Black et al., 2007). Using response surface methodology, Gao et al. (2006) found that the optimal HHP process parameters for a 6-log reduction of L. monocytogenes in whole milk were 448 MPa and 41°C for 11 min. Jofré et al. (2009) reported that HHP treatment at 600 MPa was a highly effective postharvest strategy for decreasing levels of L. monocytogenes contamination and extending the safe shelf-life of refrigerated meat products.
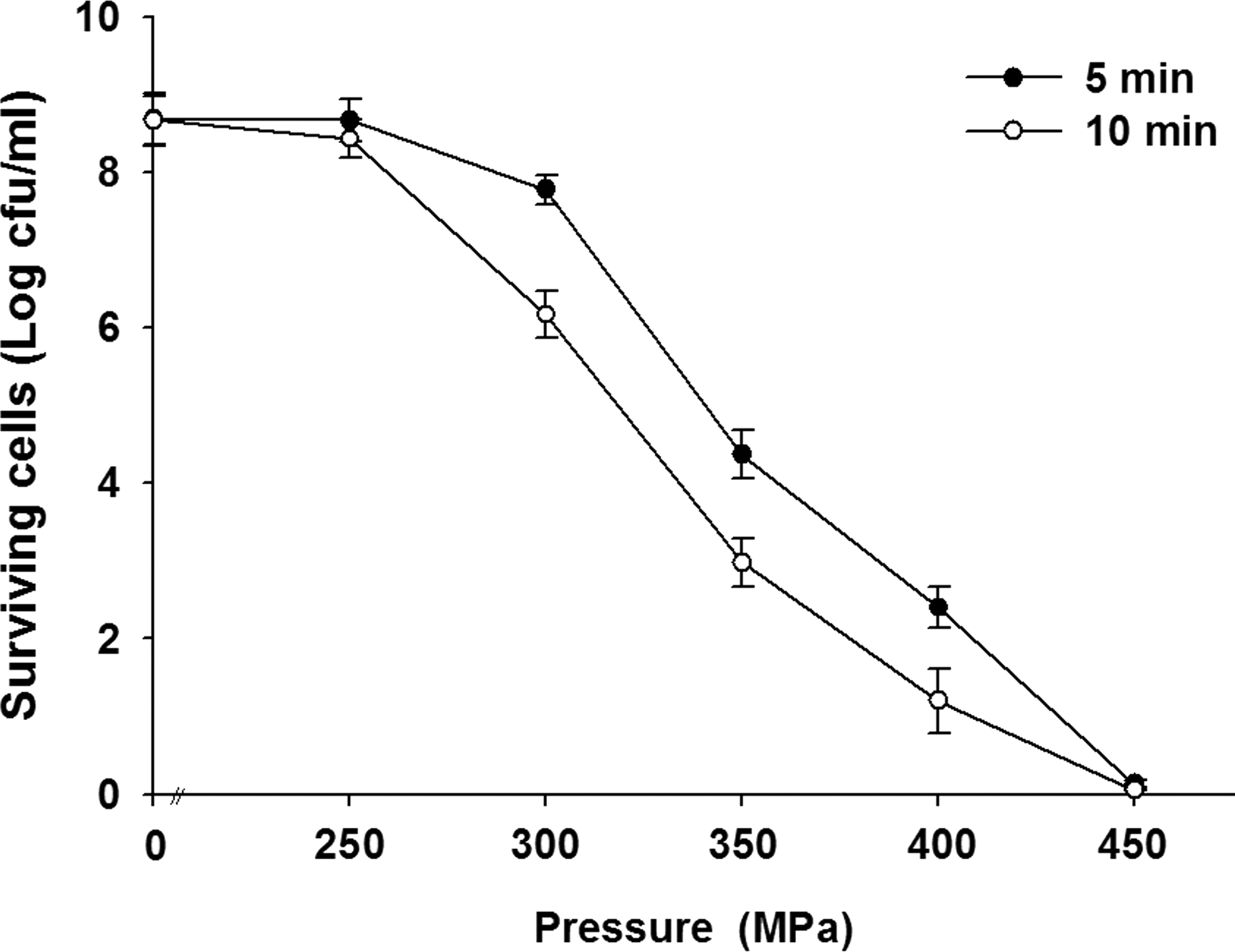
Effects of high hydrostatic pressure (100–400 MPa) on survival of Listeria monocytogenes cells. Data are mean±SD (n=3). cfu, colony-forming units.
Uptake of fluorescent dyes after exposure to HHP
The results show the difference in the amount of propidium iodide (PI) taken up before and after pressure treatment (Fig. 2); uptake of PI indicates a permanent loss of membrane integrity. As the levels of pressure increased, the number of red cells also increased, indicating that higher pressure caused more cell injury or death. Figure 3 shows the levels of PI uptake by cells at different pressures. High-pressure treatment of L. monocytogenes increased uptake of PI by 2.9 times and 4.8 times at 400 and 450 MPa for 5 min, respectively, compared with uptake by untreated cells. PI is a DNA-binding fluorescent dye that does not penetrate intact cytoplasmic membranes. PI fluorescence was previously used to assess pressure-mediated damage to cytoplasmic membranes and revealed irreversible membrane damage after pressure holding times inactivating the majority of the cell population (Gänzle and Vogel, 2001). Increased uptake of PI by Escherichia coli cells under various pressures was reported by Klotz et al. (2010). In our study, the loss of physical integrity of outer and inner membranes of L. monocytogenes cells was demonstrated by the increased uptake of PI (Moussa et al., 2007).
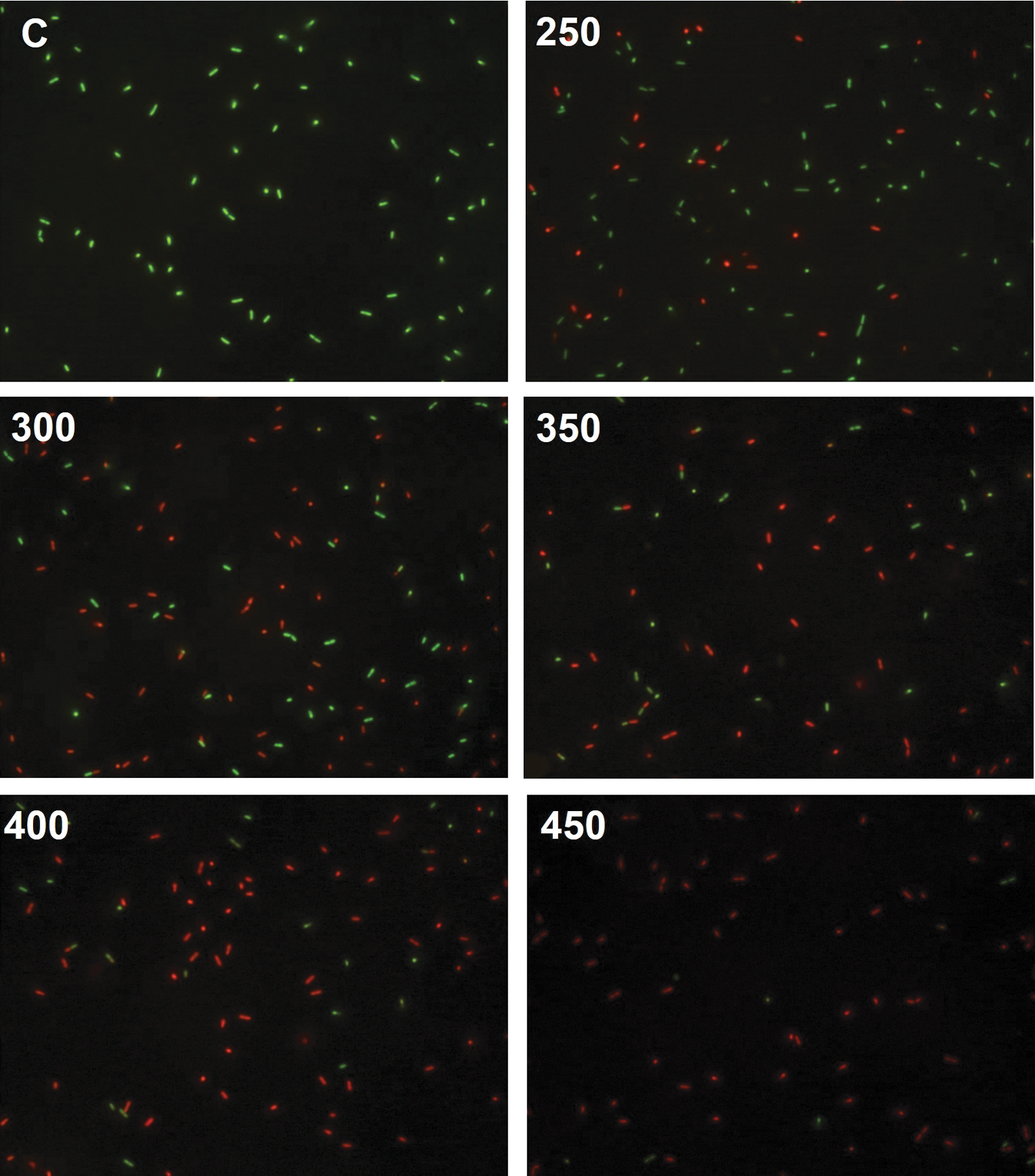
Fluorescence micrographs of untreated Listeria monocytogenes cells (C) and cells treated at pressures of 250, 300, 350, 400, and 450 MPa for 5 min. Cells were stained with SYTO 9 green fluorescent nucleic acid stain and propidium iodide (red fluorescent nucleic acid stain).
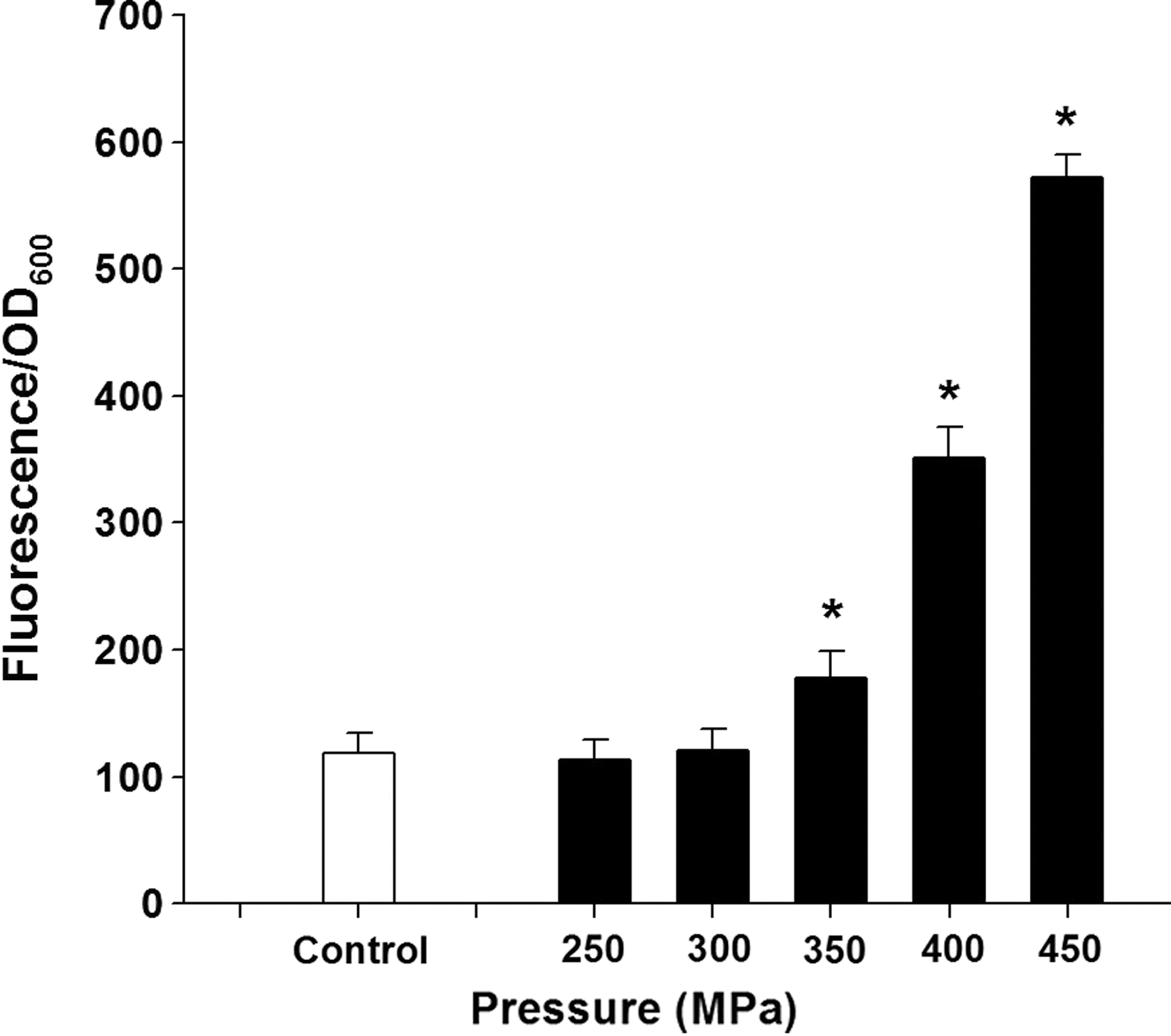
Uptake of propidium iodide by Listeria monocytogenes cells before (control) and after high-pressure treatment for 10 min at different pressures. Data are mean±SD (n=3). *p<0.05 indicates a significant difference from the control group. OD, optical density.
Scanning electron microscopy (SEM) and transmission electron microscopy (TEM) analysis after exposure to HHP
Figure 4 shows apparent damage to the cellular twist along with extracellular damage in L. monocytogenes cells after treatment with 450 MPa pressure for 5 min. In contrast, no damage was observed in untreated cells. The HHP-treated cells showed a progressive increase in the number of pimple-like lesions and swellings; some cells were crushed and others disintegrated. In the previous study, electron micrographs showed that L. monocytogenes and Salmonella enterica serovar Thompson exhibited different levels of resistance to pressure, and some changes in the cell morphology and intracellular enzyme activity after pressure treatments were dependent upon the species (Mackey et al., 1994). Similar results have been obtained for Vibrio parahaemolyticus, E. coli, Shigella spp., and Staphylococcus aureus (Yang et al., 2012; Wang et al., 2013). TEM analyses indicated that high pressure causes cell envelope and intracellular damage, unlike that observed in control cells (Fig. 5). Results from another SEM study showed that pressure treatment caused bud scars on the surfaces of cells and that membrane integrity was lost in most of the cell population. These observations suggest that the cellular wall or membrane could be a target of high-pressure treatment but that intracellular damage should not be overlooked (Huang et al., 2014). The intracellular low-density regions observed in L. monocytogenes (Fig. 3) are similar to those observed by Garcia-Gonzalez et al. (2010) in pressure carbon dioxide–treated L. monocytogenes and E. coli. These authors suggested that the low-density regions are caused by transient membrane invaginations under pressure and are subsequently reversed upon pressure release, except for the low-density regions adjacent to the cell membrane.
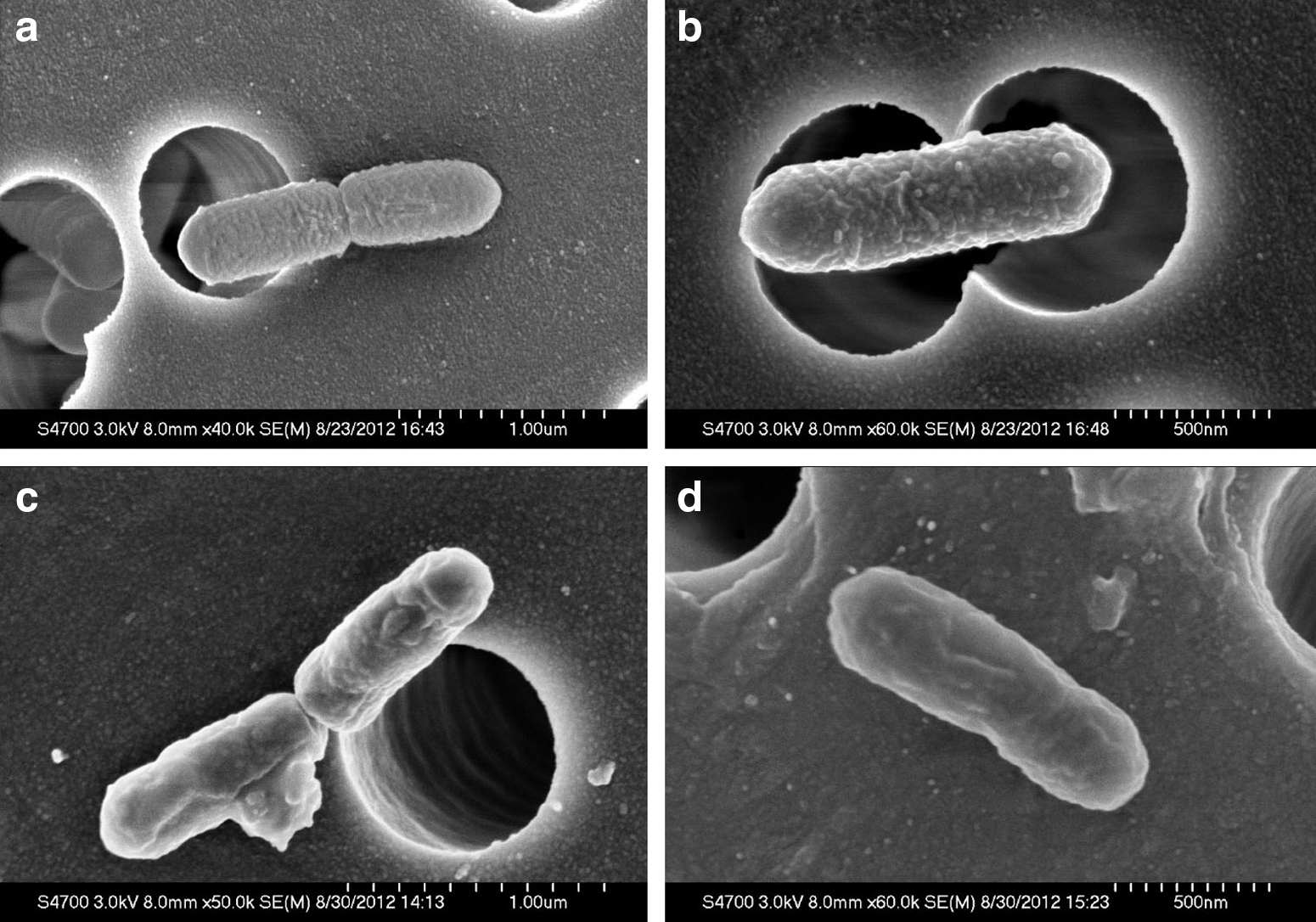
Scanning electron micrographs of Listeria monocytogenes cells:
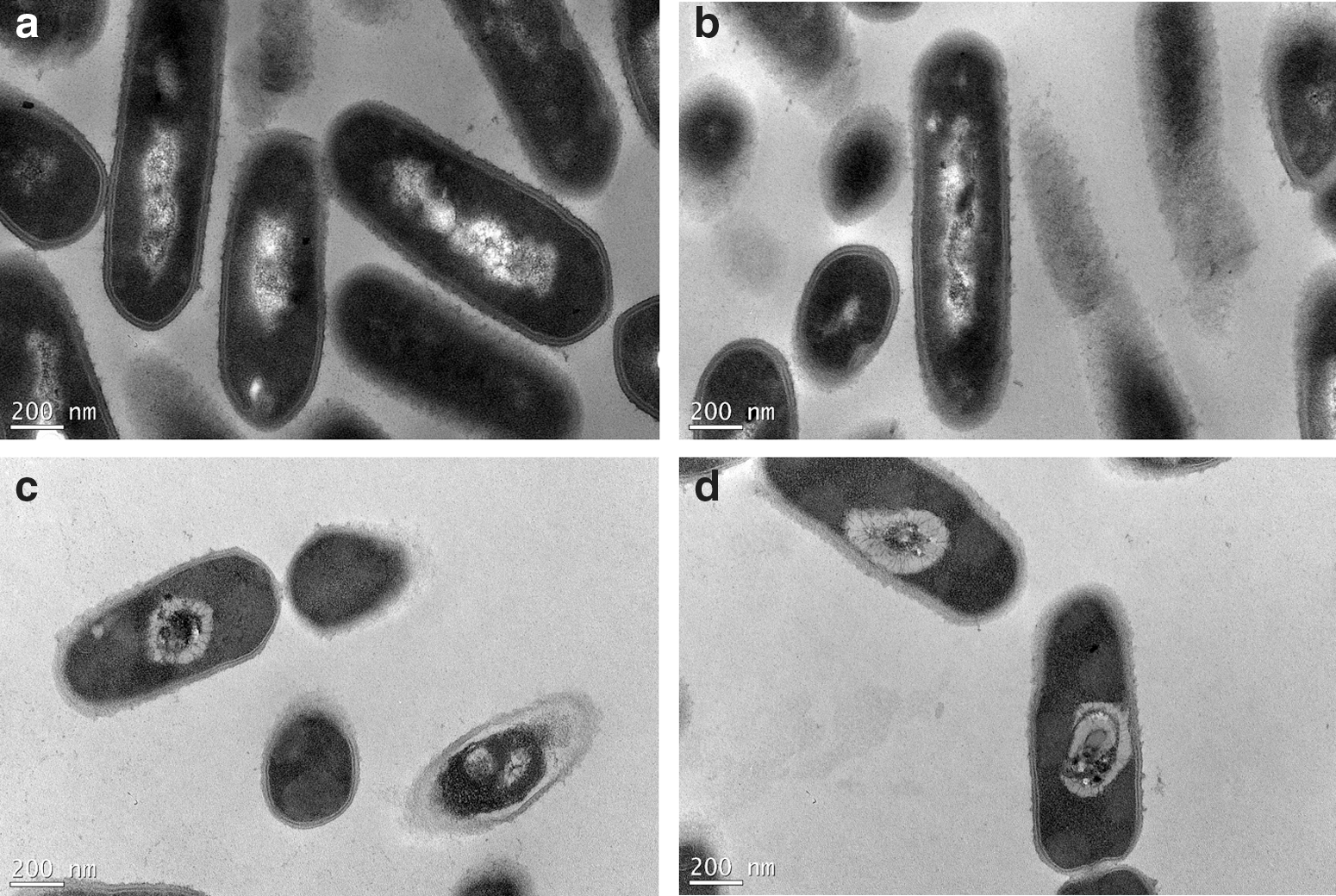
Transmission electron micrographs of Listeria monocytogenes cells:
Effect of HHP on membrane proteins
The effect of pressure on the whole-cell protein of L. monocytogenes cells is shown in Figure 6. After treatment at pressure >250 MPa, the bacterial cell population decreased, and the associated membrane protein content drastically changed. Membrane extracts from cells treated at 400 or 450 MPa showed a major protein band (45 kDa) appearing in a similar quantity unlike that in control cells, and 2 proteins (40 kDa and 25 kDa) that were observed in control cells seemed to disappear upon pressure treatment. Microorganisms are adversely affected by HHP treatment through damage to the cell wall, membranes, enzymes, and nucleic acids. In this study, our results show that hyperbaric conditions drastically alter cell morphology and membrane proteins, leading to the inactivation of L. monocytogenes cells. There seems to be a strong relation between the onset of membrane damage and loss of viability in this bacterial species. The viability of L. monocytogenes cells considerably decreased at 450 MPa because this treatment caused the cell rupture. This finding is in contrast to the results of Spilimbergo et al. (2009), who observed that the initial cellular envelope damage was not lethal to cells. Pressure application leads to the disruption of weak bonds, thereby causing the unfolding of proteins. When pressure is released, proteins refold in different configurations (Ray et al., 2001). In this study, SDS-PAGE results showed a difference in the pattern of protein bands between untreated and HHP-treated L. monocytogenes cells. These results indicate that the bacterial proteins were damaged during pressure treatment. This could be one of the most important potential mechanisms of inactivation by HHP.

Sodium dodecyl sulfate–polyacrylamide gel electrophoresis analysis of membrane proteins of Listeria monocytogenes cells after pressurization (250–450 MPa) for 5 min. M, molecular markers; C, control.
Conclusions
This study made it possible to investigate various cellular targets for high-pressure treatment of L. monocytogenes cells in whole milk. Electron micrographs have shown that pressurization significantly elicited modifications of morphological characteristics of cells. Some physical damage was inflicted, as reflected by the development of buds on the cell surface, and membrane integrity was lost in most of the treated cells, leading to increase uptake of PI and loss of viability. The possible mechanisms for inactivation of L. monocytogenes include cell membrane damage, cell wall rupture, and membrane protein degradation. However, in addition to causing other biochemical changes that probably occur simultaneously, therefore, we believe that the results of our study provide valuable information for future studies.
Footnotes
Acknowledgments
This research work was supported by the Ministry of Economic Affairs, 103-EC-17-A-22-0719, Taiwan, Republic of China.
Disclosure Statement
No competing financial interests exist.