
Abstract
In this study, 15 Gram-negative isolates from Minas Frescal cheese sold in commercial establishments in Rio de Janeiro, Brazil, were able to produce antimicrobial substances (AMSs). Seven, four, two, one, and one isolates identified as Yersinia, Acinetobacter, Enterobacter, Escherichia, and Hafnia genera, respectively, were considered potentially pathogenic. All 15 AMS+ isolates were resistant to at least 1 antibiotic; however, 7 strains presented resistance to at least 3 antibiotics from different classes, exhibiting multiresistance profiles. The strains were also subjected to plasmid profile analysis. All isolates presented different plasmid forms with most ranging in size from 1 to 10 kb. Activity against various pathogens associated with food was tested and all 15 AMS+ showed the same activity spectrum, inhibiting all Escherichia coli and Salmonella strains that were tested. Although restricted, the action spectrum of AMS-producing strains is extremely relevant to the food industry because Gram-negative bacteria such as E. coli and Salmonella spp. are most often associated with foodborne illnesses. The findings of this study reveal that even AMS produced by pathogens can have potential applications against other foodborne pathogens.
Introduction
T
Brazil is the third largest cheese producer in the world. Along with mozzarella and Prato cheese, Minas Frescal, a fresh white cheese, is one of the most consumed in Brazil (USDA, 2012; Gomes et al., 2013). During its production, however, inadequate hygienic conditions can promote pathogenic contamination and spoilage (Gomes et al., 2011; Cusato et al., 2013; Freitas et al., 2013).
Another important point involved in the microbiological quality of cheeses is related to the reduction of sodium. In recent decades, health risks associated with high sodium intake is leading the manufacturers to develop low-sodium alternatives; however, sodium chloride has shown efficiency against pathogenic and spoilage microorganisms in a variety of food systems, including Minas Frescal cheese (Gomes et al., 2011; Hystead et al., 2013).
According to Verraes et al. (2013), food may also be contaminated with antimicrobial-resistant bacteria from soil, water, or even animal and human fecal materials. The consequences of antimicrobial-resistant pathogenic bacterial ingestion involve an increase in the number of hospitalizations, and consequently, an increased risk of invasive infection and mortality (Mølbak, 2004; Verraes et al., 2013).
Current approaches to improve food security have relied on the development of more efficient chemical preservatives or application of more drastic treatments, such as high temperatures. However, these solutions have many drawbacks, such as alterations in the organoleptic and nutritional properties of foods and increased toxicity (Ananou et al., 2007).
In this current scenario, antimicrobial substances (AMSs) produced by bacteria, particularly bacteriocins, can be perceived as alternatives to improve food preservation. The use of these substances plays an important role in biopreservation, an organic method of food preservation that is defined as the extension of shelf life and food safety through natural or controlled microbiota and/or antimicrobial compound use (Ananou et al., 2007; Gálvez et al., 2010).
Studies investigating AMS application as biopreservatives in food matrices have found that such applications appear to be good alternative methods for the prevention and control of food spoilage and growth of pathogenic bacteria (Balciunas et al., 2013). However, bacteriocins produced by Gram-positive bacteria, the most studied AMS today, almost entirely inhibit the growth of other Gram-positive bacteria. Gram-negative pathogens, such as Escherichia, Salmonella, and Yersinia, are hardly inhibited by bacteriocins of Gram-positive bacteria, therefore possibly producing antibiotic-resistant genes and making treatment of cases, when necessary, more difficult (Fleming et al., 2010). These problems highlight the importance of further studies on AMSs produced by Gram-negative bacteria that are active against these microorganisms.
We describe here the investigation of AMS production by Gram-negative bacteria isolated from Minas Frescal cheese to identify AMS-producing strains for potential use against foodborne pathogens.
Materials and Methods
Sample collection and bacterium identification
Minas Frescal cheese samples available for sale in arbitrarily chosen commercial establishments in the city of Rio de Janeiro, Brazil were collected in the packing supplied by the commercial establishments and were immediately taken for analysis. The samples were diluted and bacteria were isolated by inoculation on eosin–methylene blue agar (Himedia, São Paulo, Brazil). Every fifth isolate was selected and subjected to further procedures to identify AMS-producing isolates using a commercial Gram-negative bacteria identification kit (Bactray®, Laborclin, São Paulo, Brazil).
Gram-negative bacteria strains used as AMS testing indicators, including Escherichia coli ATCC25922, E. coli LMIFRJ, Hafnia alvei LMIFRJ, Klebsiella pneumoniae ATCC4352, Pseudomonas aeruginosa ATCC27853, Pseudomonas fluorescens ATCC13525, Proteus mirabilis LMIFRJ, Proteus vulgaris LMIFRJ, Salmonella enterica ATCC19214, Salmonella spp. I-LMIFRJ, Salmonella spp. VI-LMIFRJ, and Yersinia enterocolitica ATCC9610, were grown in Casoy broth (Himedia) at 37°C for 18 h. When necessary, the medium was supplemented with agar at 1.5% or 0.6% (wt/vol). All bacteria were stored in Casoy broth with 40% glycerol (wt/vol) at −20°C until required. LMIFRJ strains were isolated from foods in our previous studies.
AMS production and action spectrum
An agar-spot assay was performed as described by Giambiagi-deMarval et al. (1990) with minor modifications. AMS-producing cells were grown in 5 mL of Casoy broth (Himedia) for 24 h, and 5 μL of culture was spotted on Casoy agar plates. After incubation for 24 h at 37°C, the bacteria were killed by exposure to chloroform vapor and the plates were sprayed with the indicator strain culture (0.3 mL of a previously grown culture in 3 mL of Casoy soft agar). The plates were further incubated for 24 h at 37°C and clear zones around growing AMS-producing strains (inhibition zones) were measured. E. coli ATCC25922 was used as an indicator strain. To determine the action spectrum of the AMSs detected, Gram-negative bacteria from different genera were used as indicators.
Antibiotic resistance profile
Resistance patterns were determined by disc diffusion, in accordance with the procedures of Clinical and Laboratory Standards Institute (CLSI, 2013). Antibiotic resistance to the following antibiotics were tested: amikacin (30 μg), amoxicillin-clavulanic acid (20/10 μg), ampicillin (10 μg), cephalothin (30 mg), ceftazidime (30 μg), cefotaxime (30 mg), chloramphenicol (30 μg), ciprofloxacin (5 μg), gentamicin (10 μg), imipenem (10 μg), kanamycin (30 μg), nalidixic acid (30 μg), norfloxacin (10 μg), streptomycin (10 μg), tetracycline (30 μg), tobramycin (10 μg), and trimethoprim (5 μg). All these were obtained from Sensifar (São Paulo, Brazil). E. coli ATCC25922 was used as a control strain.
Plasmid profile
Plasmid DNA was isolated using a Qiaprep® Spin Miniprep Kit (Qiagen). Agarose gel electrophoresis was performed using 0.7% (wt/vol) agarose gels, as described by Sambrook et al. (1989).
Results and Discussion
Due to high moisture content, Minas Frescal cheese is a suitable medium for the growth of pathogens frequently associated with foodborne diseases. Gram-negative bacteria are considered an important hazard to the food safety of cheeses (Carvalho et al., 2007; Pintado et al., 2014).
Of the 50 Gram-negative isolates from the analyzed samples, 15 (30%) were able to produce AMSs against E. coli ATCC25922. Seven isolates were identified as Y. intermedia, four as Acinetobacter baumannii-calcoaceticus complex, one as Enterobacter sakazakii (Cronobacter sp.), one as E. cloacae, one as E. coli, and one as H. alvei (Table 1). To our knowledge, this is the first report on AMS production by Acinetobacter spp.
AMC, amikacin; AMP, ampicillin; CFL, cephalothin; IPM, imipenem; NOR, norfloxacin; STR, streptomycin; TET, tetracycline; TRI, trimethoprim.
Antibiogram test results showed that all isolates were resistant to at least one antibiotic. However, 7 isolates (Q5, Q29, Q37, Q38, Q44, Q45, and Q46) were resistant to at least 3 classes of antibiotics, expressing typical multiresistant profiles (Boucher et al., 2009; Heizmann et al., 2013).
A high incidence of resistance to antibiotics commonly administered to both human and animal populations, such as cephalothin (86.7%), ampicillin (80%), and the combination of amoxicillin–clavulanic acid (40%), was observed among the AMS+ isolates. Some studies have reported an association between the antibiotic use and antimicrobial resistance of foodborne pathogens, suggesting that controlling antibiotic administration to food animals is necessary to reduce the incidence of resistant organisms. However, current control measures are insufficient to limit the transmission of resistance through the food chain (Depoorter et al., 2012; van Boxstael et al., 2012; Verraes et al., 2013).
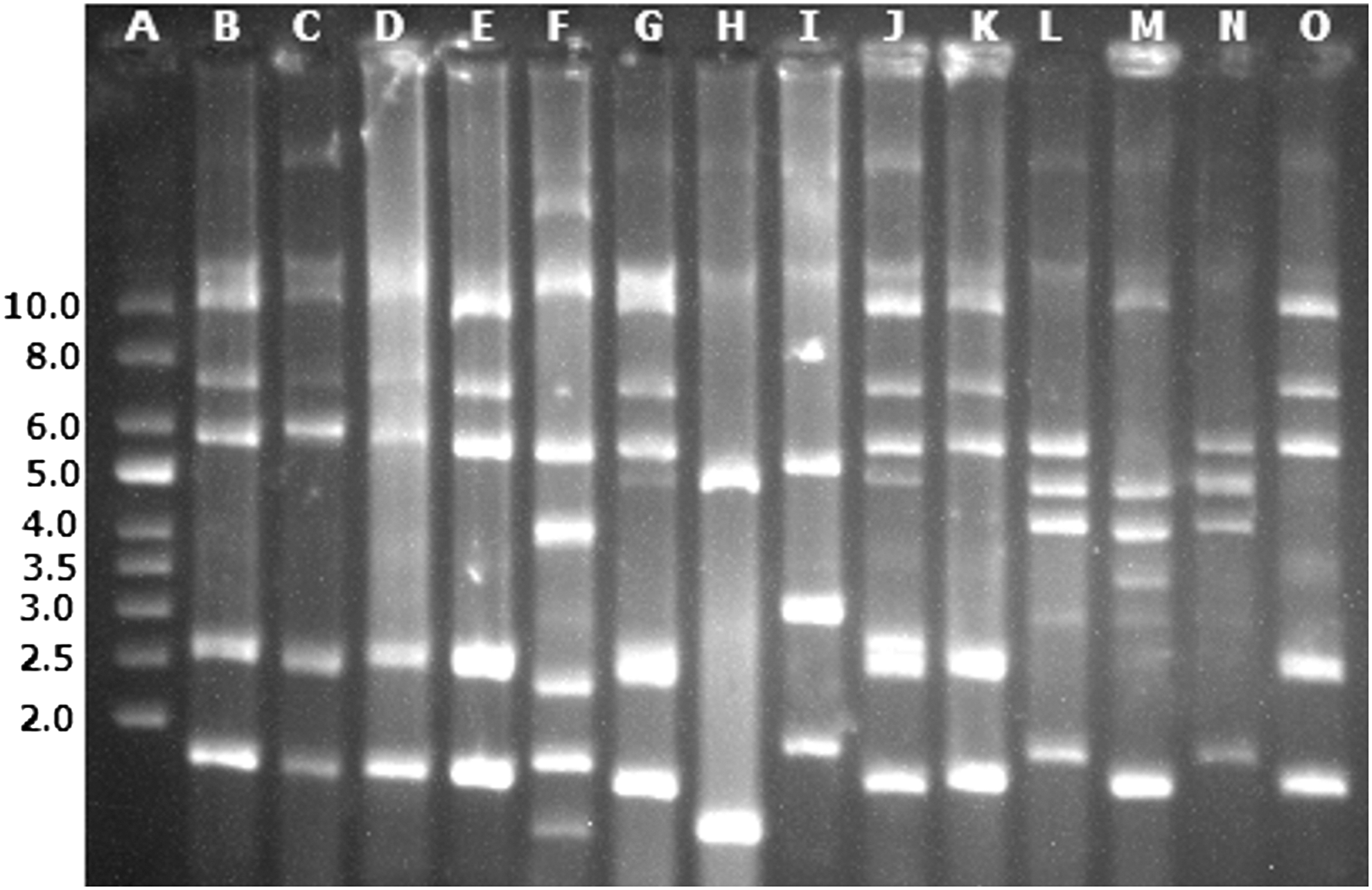
Besides the selective pressure created by antimicrobial use behind the emergence of resistance, in some cases, this genetically encoded resistance can be transferred horizontally, even to distantly related bacteria, by genetic transference, including transduction, transformation, and conjugation, often involving plasmids (Frye and Jackson, 2013). Therefore, plasmid profiles of the AMS+ isolates were evaluated using only small plasmids, appearing as bright bands on the gel mostly below the chromosomal DNA band. Plasmid DNA analysis revealed that all 15 strains harbored multiple plasmids with the majority ranging in size (approximately 1–10 kb; Figure 1). Seven plasmid profile groups were identified, with group I accounting for the largest number of strains (Table 1).
Regarding the action spectrum, all 15 AMS+ isolates, even those belonging to different genera and presenting different characteristics, showed the same activity spectrum, inhibiting only the E. coli and Salmonella strains that were tested. The inhibition zone diameters varied from 19 to 30 mm. Figure 2 illustrates E. coli ATCC25922 and S. enterica ATCC19214 inhibition by the producer strains.
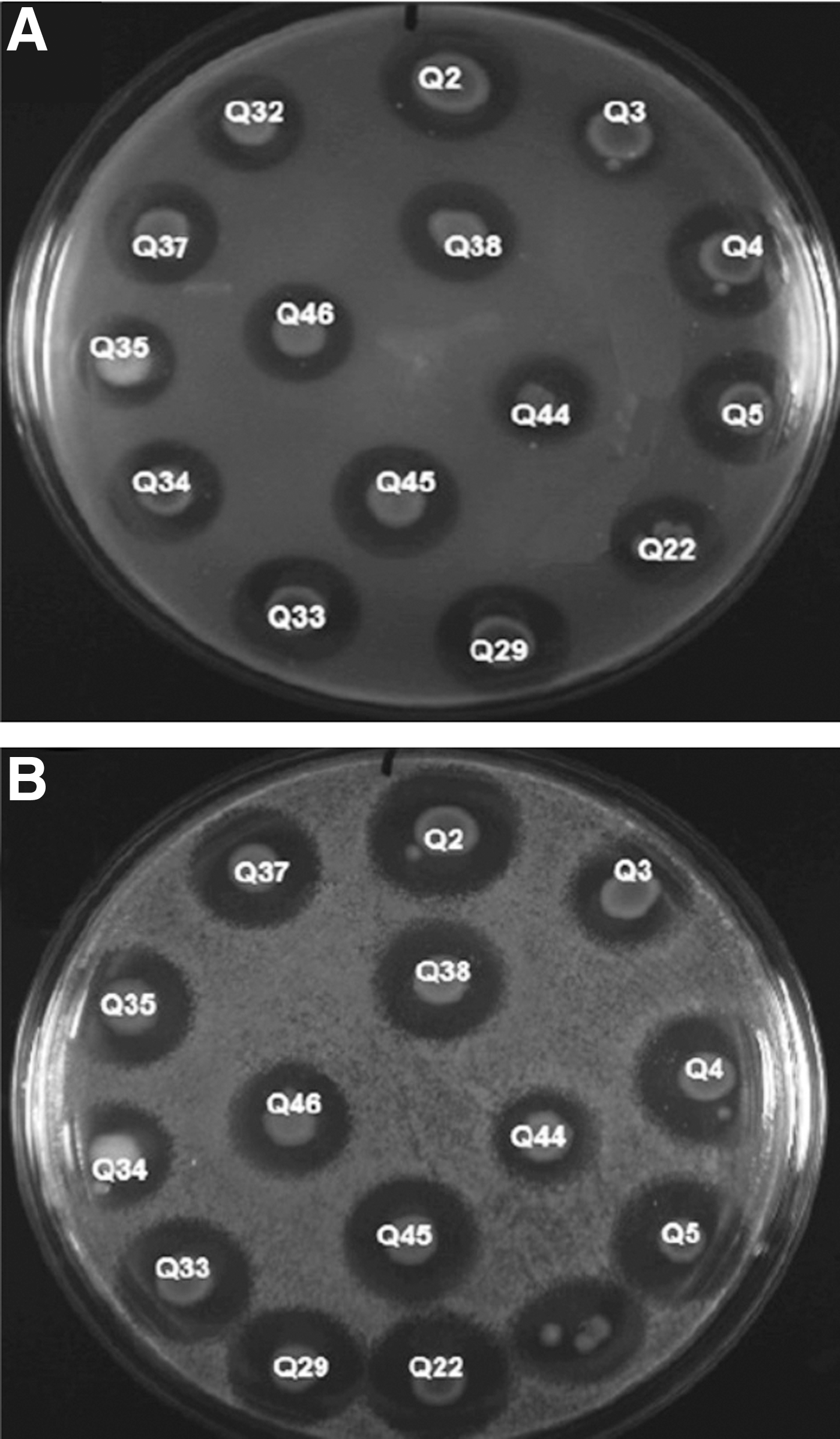
Inhibition of Salmonella enterica ATCC19214 (
Similar results were reported in a previous study that identified two coliform strains isolated from salad, cheese, and meat products as Klebsiella ozaenae and Raoultella terrigena, exhibiting inhibitory activity against E. coli and S. enterica indicator strains (Fleming et al., 2010).
Although restricted, the action spectrum of the AMS-producing strains is extremely relevant to the food industry because E. coli and Salmonella spp. represent Gram-negative bacteria that are most associated with foodborne illnesses (Sayah et al., 2005; Lamo-Castellví et al., 2007).
Conclusions
Gram-negative populations can reach high levels in cheeses and present wide species diversity. Although pathogenic species are potentially dangerous, their purified antimicrobial substances can positively contribute to the inhibition of others, extending the shelf life of cheeses by delaying or preventing growth of pathogens such as Salmonella and Escherichia coli. Because the 15 AMS+ strains identified in this study could have potential application against foodborne pathogenic strains, further studies to develop these AMSs are warranted.
Footnotes
Acknowledgments
This research was supported by grants from IFRJ and FAPERJ.
Disclosure Statement
No competing financial interests exist.