
Abstract
The 63 pulsed-field gel electrophoresis (PFGE) types identified among 427 clinical isolates of Listeria monocytogenes that were characterized in a previous study by serotyping and PFGE (AscI) could be further divided into 17 PFGE groups. While the 63 PFGE types, all part of lineage I, were established based on the number and distribution of all bands in each DNA profile, the 17 PFGE groups were based on the configuration of small bands with sizes <145.5 kb. The 30 PFGE types of L. monocytogenes serovar 4b isolates (n=334) were divided into 8 PFGE groups; the 32 PFGE types of serovar 1/2b isolates (n=90) and the serovar 3b isolates (n=3, 1 PFGE type) were divided into 9 PFGE groups. An association was observed between PFGE groups and serovars. L. monocytogenes isolates belonging to PFGE groups I, J, Q, R, X, Z, Ö-4, and Ö-5 all shared serovar 4b, whereas isolates from PFGE groups D, G, O, P, T, U, Ö-1, Ö-2, and Ö-3 shared serovar 1/2b. Small fragments <33.3 kb were nonvisible in all L. monocytogenes isolates. From the results of the present study, a procedure for accelerating the identification of PFGE types when analyzing new PFGE profiles can be suggested. Therefore, we propose a stepwise procedure to PFGE profiling by first identifying the PFGE group using the smaller band patterns <145.5 kb, and then determining PFGE types based on the band patterns >145.5 kb.
Introduction
T
During the 1970s, 1980s, and 1990s, the global incidence of human listeriosis was mainly caused by L. monocytogenes serovar 4b (Guevara et al., 1979; Kathariou, 2002; Lopez-Valladares et al., 2014), although several 1/2a outbreaks have been reported since 2000 (Tham and Danielsson-Tham, 2014). However, serotyping is a poorly discriminating method for the epidemiological characterization of bacterial isolates associated with foodborne outbreaks.
Pulsed-field gel electrophoresis (PFGE) using restriction enzyme AscI is well established as an effective molecular subtyping method for characterizing L. monocytogenes (Brosch et al., 1996; Kerouanton et al., 1998; Lopez-Valladares et al., 2014) with utility in the identification of human listeriosis outbreaks (Buchrieser et al., 1993; Ericsson et al., 1997; Goulet et al., 1998; Danielsson-Tham et al., 2004; Knabel et al., 2012). L. monocytogenes isolates from human patients suffering from invasive listeriosis in Sweden between 1958 and 2010 have been effectively characterized by serotyping and PFGE (Lopez-Valladares et al., 2014). However, examination and assignment of PFGE types to specific profiles is a delicate and tedious process that would benefit from simplification.
Therefore, the main aim of this study was to accelerate the identification of PFGE profiles first by grouping the band patterns <145.5 kb and then by subtyping those >145.5 kb using PFGE band patterns of 427 L. monocytogenes isolates belonging to lineage I (serovars 4b, 1/2b, and 3b) as the data source. Possible association between PFGE groups and serovars was also attempted.
Materials and Methods
L. monocytogenes isolates
Lineage I L. monocytogenes (n=427) isolated during 1958–2010 from human invasive listeriosis in Sweden was selected for the present study. All of the isolates were previously characterized by serotyping and PFGE with restriction enzyme AscI according to Graves and Swaminathan (2001) with some modifications (Lopez-Valladares et al., 2014). PFGE types were established based on the number and distribution of all bands in each DNA profile. The profiles were considered distinguishable if there was a difference of one band or more. Serovar 4b isolates (n=334) were distributed over 30 PFGE types; 90 serovar 1/2b isolates gave 32 PFGE types and 3 serovar 3b isolates gave 1 PFGE type (Table 1). For storage, isolates were maintained in brain-heart infusion broth (Merck, Darmstadt, Germany) with 20% glycerol at −70°C.
Shared the same PFGE group (G) as some serovar 1/2b isolates.
Grouping of L. monocytogenes PFGE types
In a previous study, the 63 PFGE types of L. monocytogenes were established based on the number and distribution of all fragments in a DNA restriction profile (Lopez-Valladares et al., 2014). The AscI restriction fragments of each PFGE type were analyzed visually and sized against a Lambda ladder PFGE marker and the universal marker Salmonella Braenderup H9812. In the present study, PFGE groups were established solely on the number and distribution of small bands with sizes <145.5 kb. Thus, each PFGE group included several different PFGE types with identical configuration of bands with sizes <145.5 kb. The PFGE groups were designated by capital letters. Each isolate of L. monocytogenes was then assigned a multipart code. For example, the designation D:1/2b:75A indicates that the isolate belonged to PFGE group D, serovar 1/2b, and represented the 75th AscI profile within the serogroup 1/2 portion of the collection, respectively. The last character (i.e., ‘‘A’’) refers to variation among the AscI profiles. AscI-profile no. 75 comprises closely related variants (A, B, C) (Tenover et al., 1995). The AscI profiles of L. monocytogenes isolates representing the different PFGE groups are presented in Figure 1.
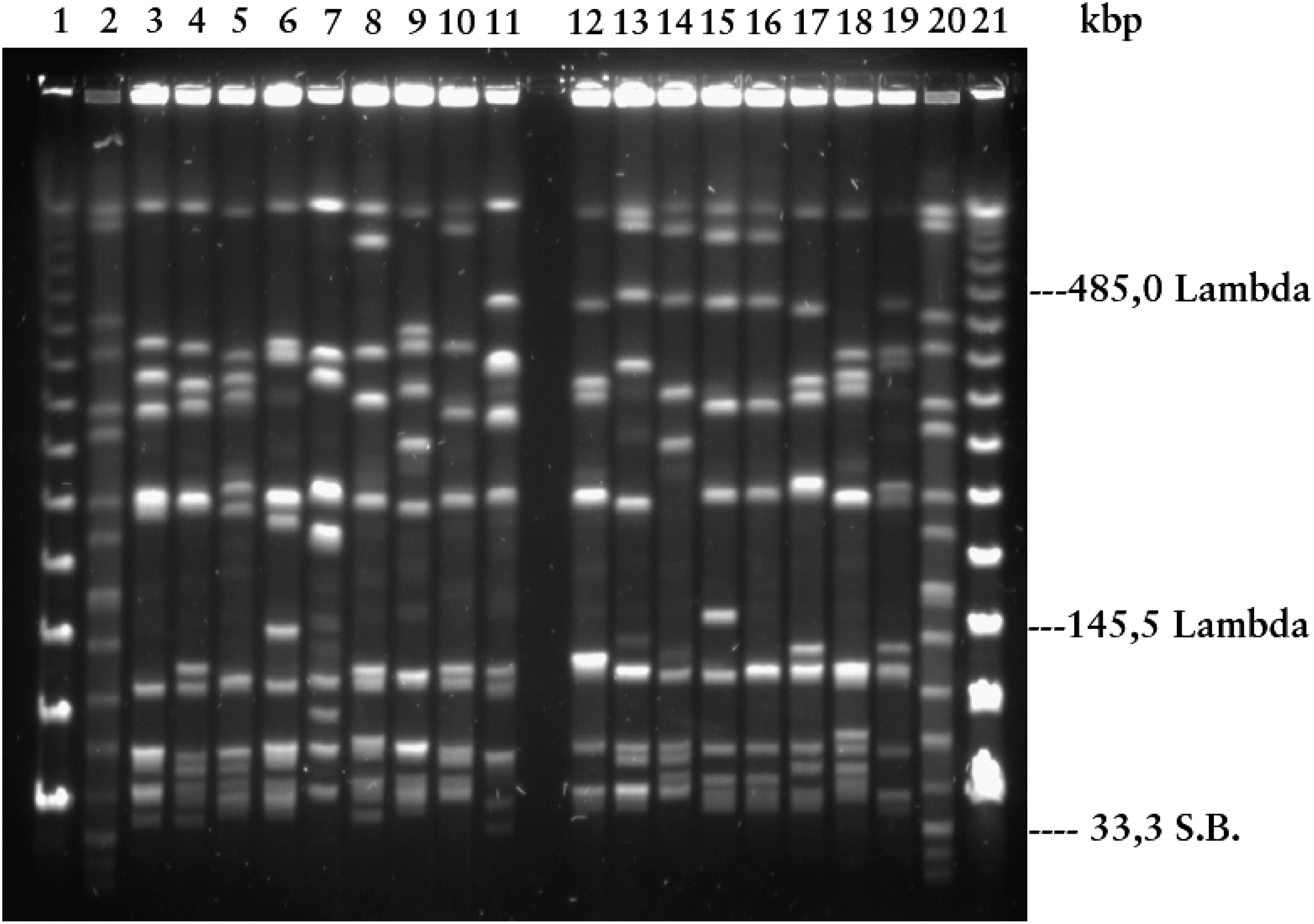
AscI profiles of Listeria monocytogenes lineage I, human strains. Electrophoretic conditions: 21 h, pulse times 4–40 s, 120° angle, and 14°C. Lane 1 (Lambda ladder PFGE marker No. 340 S); Lane 2 (Salmonella Braenderup H9812); Lane 3 (group G); Lane 4 (group D); Lane 5 (group O); Lane 6 (group P); Lane 7 (group T); Lane 8 (group U); Lane 9 (group Ö-1); Lane 10 (group Ö-2); Lane 11 (group Ö-3); Lane 12 (group I); Lane 13 (group J); Lane 14 (group Q); Lane 15 (group R); Lane 16 (group X); Lane 17 (group Z); Lane 18 (group Ö-4); Lane 19 (group Ö-5); Lane 20 (Salmonella Braenderup H9812); and Lane 21 (Lambda ladder PFGE Marker No. 340 S).
Results and Discussion
The 63 PFGE types previously identified in 427 clinical L. monocytogenes isolates could be further divided into 17 PFGE groups based on the position of distinctly detectable bands <145.5 kb. Small fragments <33.3 kb were nonvisible in all L. monocytogenes isolates (Fig. 1). Five of the 17 PFGE groups were each represented by only 1 isolate, and were designated as capital letter Ö followed by a number (e.g., Ö-1). The 30 PFGE types of L. monocytogenes serovar 4b isolates (n=334) were divided into 8 PFGE groups (an average of 41.75 isolates per PFGE group); the 32 PFGE types of serovar 1/2b isolates (n=90) were divided into 9 PFGE groups (10 isolates per PFGE group). One of the nine serovar 1/2b PFGE groups also included the three serovar 3b isolates sharing one PFGE type (Table 1). The number of L. monocytogenes isolates, serovars, PFGE types, and PFGE groups are presented in Table 2. Numbers and PFGE groups of L. monocytogenes isolates, per year, from 1958 to 2010, are shown in Table 3. In Sweden, L. monocytogenes isolates belonging to the two most widespread PFGE groups over time (i.e., D and J) have been isolated from human cases over six decades. However, L. monocytogenes PFGE group I is currently the dominant PFGE group within lineage I in Sweden and has been responsible for 193 cases of listeriosis during 1958–2010. One of the most common PFGE types (4b:6) belonging to PFGE group I was identical to the epidemic cheeseborne L. monocytogenes strain in Switzerland, Denmark, and the United States during 1983–1990 (Lopez-Valladares et al., 2014) (Table 3).
An association was observed between PFGE groups and serovars. L. monocytogenes isolates belonging to PFGE groups I, J, Q, R, X, Z, Ö-4, and Ö-5 all shared serovar 4b. Isolates from PFGE groups D, G, O, P, T, U, Ö-1, Ö-2, and Ö-3 shared serovar 1/2b. With one exception, PFGE group G (which included PFGE type G:3b:2B) was associated with serovar 3b (Table 2). The occurrence of atypical combinations of serotypes and grouping based on different molecular typing methods has also been observed in other studies (Buchrieser et al., 1991; Nadon et al., 2001; Revazishvili et al., 2004). A portion of our L. monocytogenes collection has been characterized by multilocus variable-number tandem-repeat analysis with the resulting clusters generally agreeing with both the data generated by PFGE as proposed here and serotyping data (Lindstedt et al., 2008). A study of Staphylococcus aureus has shown that the configuration of small PFGE bands (10–85 kb) correlates better with the phylogenetic typing method multiprimer random amplified polymorphic DNA analyses than larger PFGE bands (>85–700 kb). The authors propose that this is due to a greater propensity for size instability in larger fragments since the frequency of random genetic events increases with the size of the DNA fragment (Blanc et al., 2002). In another staphylococci study, PFGE showed good concordance with multilocus sequence typing, with authors suggesting that PFGE is a reliable method for long-term, nationwide epidemiological surveillance studies (Hallin et al., 2007).
In a small study, Brosch et al. (1991) digested L. monocytogenes, serovar 4b isolates (n=42), with ApaI producing 16 PFGE types that could be further divided into 4 PFGE groups as defined by the close relatedness of 9–12 lower-molecular-weight fragments. Later, Brosch et al. (1994) compared low-molecular-weight AscI restriction fragments (23–60 kb) of L. monocytogenes isolates (n=176) and identified 2 genomic divisions that correlated with the flagellar (H) antigen type: division I (1/2a, 1/2c, 3a, 3c) and division II (1/2b, 3b, 4b, 4d, 4e). Their results, however, cannot be directly compared with the results from the present study, as different analytical approaches were used. In silico analyses (PFGE with restriction enzyme AscI) of different sequenced L. monocytogenes strains belonging to lineage I are presented in Table 4 (GenBank or EMBL-EBI). Although the strains are from different sources, time, and location, their AscI restriction fragments below the size of 145.5 kb are more conserved than larger fragments, supporting the utility of smaller fragments in grouping L. monocytogenes. Interestingly, a small fragment (1454 bp) was present in all L. monocytogenes strains.
AscI restriction fragment sizes were calculated from L. monocytogenes whole genome sequences obtained from either GenBank or EMBL-EBI.
Number of base pairs in restriction fragment.
From the results of the present study, a procedure for improving the identification of PFGE types when analyzing new PFGE profiles is proposed. First the smallest banding patterns below 145.5 kb are analyzed to determinate the PFGE group. Then, within the identified PFGE group, the banding patterns above 145.5 kb are analyzed to determine the PFGE type.
Conclusions
L. monocytogenes PFGE groups were established for each isolate based on the configuration of small AscI restriction fragments below the size of 145.5 kb. Within the identified PFGE group, the banding patterns above 145.5 kb were utilized to determine the PFGE type. This process simplifies the identification of PFGE types. Moreover, it is possible to establish an association between PFGE groups and serovar 4b, 1/2b, and 3b (i.e., lineage I).
Footnotes
Acknowledgment
This study was supported by the Stiftelsen Grythytte Akademi Stipendiefond, to whom we express gratitude.
Disclosure Statement
No competing financial interests exist.