
Abstract
During a nationwide surveillance in Korea, 13 methicillin-resistant Staphylococcus aureus (MRSA) strains were isolated from imported and domestic meat between 2009 and 2011. The predominant MRSA genotype was SCCmec type V, and only two agr types (I and II) were found. Unexpectedly, sequence type ST72 comprised more than 50% of the isolates; this is the first instance of type ST72 in food from Canada. Two Spanish pork isolates were ST398, which caused human disease in Europe, and they carried leukotoxin genes, lukS, lukF, and lukE-lukD. Furthermore, P71 and P6 harbored all of the known leukocidin genes, lukS-lukF-lukE-lukD-lukM. Our collected MRSA strains were multidrug resistant with various antimicrobial and heavy-metal resistance genes. Toxin genes that are commonly found in clinical MRSA also were detected in our meat strains. One MRSA strain exhibited an uncommon type of enterotoxin, sec-see-seg-sei-sel-sem-sen-seo-sep. Plasmids (1.5–15.0 kb) were found in 12 of the 13 MRSA isolates. Repetitive sequence-based polymerase chain reaction of the genomic DNA showed 3 clusters with 95% similarity. The presence of multidrug-resistant and toxigenic MRSA in meat products suggests that comprehensive surveillance should be continued for imported meats in Korea.
Introduction
S
In Korea, MRSA has been harbored by meats and animals, including dogs, chickens, cattle, and pigs (Lim et al., 2012). MRSA sequence type ST398 has been described as a possible threat to human health and has been isolated from domestic animals, humans who are close contact with infected animals, and retail meats (Khanna et al., 2008; Lim et al., 2012). Therefore, there is a growing concern that MRSA strains of animal origin might be transmitted into the food chain.
Antimicrobials and metal-containing compounds are employed as a food supplement for livestock (Injac et al., 2008). MRSA strains are frequently resistant to antibiotics, including the aminoglycosides, macrolides, chloramphenicol, tetracycline, and fluoroquinolones (Barrett, 2004; Schito, 2006). MRSA strains that are resistant to zinc and cadmium have been documented, and the czrC gene has been detected in those isolates (Rosdahl and Rosendal, 1983; Cavaco et al., 2010). MRSA is able to produce various virulence factors, such as adhesins, enterotoxins, exfoliative toxins, hemolysins, leukocidins, and other toxins, which cause severe illness in humans (Jarraud et al., 2002; Campbell et al., 2008).
During a nationwide surveillance, we isolated 13 MRSA strains from imported and domestic meat between 2009 and 2011 in South Korea. The objective of this investigation was to characterize their genotypes, antimicrobial resistance, and virulence genes.
Materials and Methods
Sample collection and isolation of S. aureus strains
Between 2009 and 2011, 1391 imported meats—beef (n=378), pork (n=552), and chicken (n=461)—and 1419 domestic meats—beef (n=505), pork (n=500), and chicken (n=414)—were randomly collected by the National Monitoring Program in South Korea. Meat samples (25 g) were minced, mixed with 225 mL of buffered peptone water (Becton Dickinson [BD], Sparks, MD), and vigorously shaken for 5 min. One milliliter of rinse solution was added to 9 mL of Mueller Hinton broth (BD) supplemented with 6.5% sodium chloride, and the mixture was incubated at 36°C overnight. One milliliter of this pre-enrichment broth was transferred into 9 mL of mannitol salt broth (BD) and incubated for 48 h at 36°C. Then a portion of this broth was plated onto mannitol salt agar (BD) for S. aureus and BBL CHROMagar MRSA agar plate (BD) for MRSA selection, and incubated 24–48 h at 36°C. Presumptive S. aureus yellow-colored colonies and MRSA mauve-colored colonies were isolated. Presumptive S. aureus colonies were confirmed using a tube coagulase test with rabbit plasma (BD), a catalase test, and a latex slide agglutination test (BD). The polymerase chain reaction (PCR) amplification of nucA to confirm S. aureus strains was conducted as described previously (Louie et al., 2002). Presumptive MRSA colonies were confirmed as MRSA by performing mecA PCR (Strommenger et al., 2006) and penicillin binding protein 2' latex agglutination test (Oxoid Ltd., Hampshire, England).
Antimicrobial susceptibility
The Sensititre MIC susceptibility system (TREK Diagnostic Systems, Cleveland, OH) was used to test susceptibility to antibiotics. Minimum inhibitory concentrations (MIC) of antimicrobials were determined by broth microdilution following the guidelines of Clinical and Laboratory Standards Institute (CLSI) (CLSI, 2007). Etest (bioMérieux, Durham, NC) of oxacillin MIC was performed according to the manufacturer's instructions, using MH BBL II agar supplemented with 2% NaCl (BD) and incubation at 36°C for 18–24 h. The Etest of cefoxitin MIC was done using MH BBL II agar without NaCl. A methicillin-susceptible S. aureus strain (ATCC 25923), a MRSA strain (ATCC 33591), and an S. epidermidis strain (ATCC 12228) were used as control organisms for the antimicrobial susceptibility testing.
Chromosomal and plasmid DNA isolation
Chromosomal DNA was isolated by using a DNeasy blood and tissue kit (Qiagen, Valencia, CA) according to the manufacturer's instructions. Plasmid DNA was extracted and purified by using the QIAprep spin plasmid kit (Qiagen) with a modification consisting of 1 h incubation at 37°C in the presence of 10 μL of lysostaphin (1 mg/mL). DNA concentrations and purity were determined by measuring the absorbance at 260 nm and 280 nm in a Nanodrop 2000 UV spectrophotometer (Thermo Fisher Scientific, Wilmington, DE).
Multilocus sequence typing (MLST) and spa typing
MLST was performed as previously described by Enright et al. (2000). Amplicons of seven housekeeping genes for arcC, aroE, glpF, gmk, pta, tpi, and yqiL were obtained by PCR with the specified primers (Supplementary Table S1; Supplementary Data are available online at
Detection of genes for antimicrobial and heavy metal resistance and virulence
Antibiotic resistance genes of penicillin (blaZ, p480, p480-blaZ), tetracycline (tetK, tetL, tetM, tetS, tetW, tetO), macrolides–lincosamides–streptogramins B (ermA, ermB, ermC, mphC, sat4), macrolides (msrA, msrB), lincosamide (linA/linA), and aminoglycosides (ant(4')-la, aph(3')-llla, aac(6')-le-aph(2')-la) were examined by singlex or multiplex PCR. PCR primer and amplification conditions were previously reported (Supplementary Table S2) (Hauschild et al., 2008; Kumar et al., 2010). Genes (gyrA, gyrB, grlA, grlB, and norA) associated with ciprofloxacin resistance were amplified independently by PCR (Takahata et al., 1996; Takahashi et al., 1998). The amplicons were purified and sequenced with both forward and reverse primers. Mutations in gyrA, gyrB, grlA, grlB, and norA were identified using BLAST by comparison with the GenBank database (Schmitz et al., 1998; Noguchi et al., 2002). The presence of cadmium and zinc resistance genes in genomic DNA was analyzed by PCR with specific primers for the czrC, cadA, and cadD-cadX region (Supplementary Table S3) (Massidda et al., 2006; Cavaco et al., 2011). Virulence genes (bbp, clfA, clfB, cna, ebpS, fnbA, fnbB, map/eap, sdrC, sdrD, sdrE, sea, seb, sec, sed, see, seg, seh, sei, sej, sek, sel, sem, sen, seo, sep, seq, ser, seu, eta, etb, etd, hla, hlb, hld, hlg, hlg-2, lukS, lukF, lukE-lukD, lukM, bsaB, ear, splF, tst, edin, chp, efb, icaA, V8, arcA) were detected by singlex or multiplex PCR (Supplemental Table S4). Both positive and negative controls were included in each run of PCR. The sizes of the PCR products were determined by comparing them with the migration of appropriate DNA ladders (Life Technologies, Grand Island, NY). The amplified products were confirmed by sequencing and GenBank database homology comparison in a BLAST search.
Repetitive sequence-based PCR (rep-PCR) DNA fingerprinting
Rep-PCR was performed as previously described (Church et al., 2011). The band patterns were compared with a Pearson correlation analysis using the web-based DiversiLab software (version 2.1.66) to assess band position and intensity, and the distance matrices. To create dendrograms, the unweighted-pair group method with arithmetic mean was employed.
Results
Prevalence of S. aureus and MRSA in domestic and imported meats
Generally, higher numbers of S. aureus and MRSA were recovered from imported than from domestic meats (Table 1). In imported meats, 40.94% of the pork samples were contaminated with S. aureus, which is higher than other meats. In contrast, S. aureus was isolated from 36.23% of the domestic retail chicken meat, a prevalence that is 3–5 times greater than in the domestic beef and pork meats. Low numbers of MRSA strains were positive in imported and domestic meats. Furthermore, we investigated 63 domestic duck meats for S. aureus and MRSA, and 13 of them had S. aureus (data not shown). No MRSA was present in the duck meats.
Antimicrobial and metal resistance as well as their genetic profiles
All MRSA strains were resistant to oxacillin, cefoxitin, and penicillin (Table 2). Tetracycline resistance was the most common of these (6/13); then, clindamycin (5/13), erythromycin (4/13), and kanamycin resistance (3/13) followed. All isolates were susceptible to rifampin, chloramphenicol, vancomycin, trimethoprim, linezolid, mupirocin, and sulfamethoxazole.
Strains resistant to a specific antibiotic are highlighted in bold. MIC breakpoint criteria are not available for streptomycin and tiamulin (italics).
Among antibiotic resistance genes (blaZ, p480, p480-blaZ) associated with penicillin, isolate P71 was found to harbor both blaZ and p480 genes, and B80 to possess the blaZ gene only (Table 3). tetK and tetL genes were carried by all isolates resistant to tetracycline phenotypically. Erythromycin-resistant isolates were all positive to ermC and mphC, whereas ermA and ermB genes were all negative. Macrolide (msrA) and lincosamide (linA/linA) resistance genes were consistent with their phenotypic resistance patterns. For the aminoglycosides (kanamycin and gentamicin), the gene encoding aph(3')-llla was amplified from three isolates. Zinc resistance conferred by the czrC gene was predominant in most MRSA isolates (10/13). Among four different primer pairs between cadD and cadX, three regions were amplified from all isolates with the exception of Canadian pork isolate P219.
Genotyping of MRSA isolate
The nucA gene was found in all strains except a C71 chicken isolate from Brazil (data not shown). SCCmec type IV was detected in six isolates and, among them, type IVA was detected in all isolates except a Canadian pork isolate (IVB) (Table 4). Four MLST types (ST5, ST8, ST72, ST398) were found, and the majority of MRSA were ST72. Six spa types were observed, and the most common spa type was t148.
Genetic profiles of virulence genes
Profiles of virulence genes are summarized in Table 5. All MRSA isolates were positive for genes for adhesins (bbp, clfA, clfB, ebpS, sdrD, sdrE), enterotoxins (seg, sei, sen), hemolysins (hla, hlb, hld, hlg), leukocidin (lukE-lukD), chemotaxis-inhibiting protein (chp), fibrinogen-binding protein (efb), N-glycosyltransferase (icaA), and V8 protease (V8) genes. Adhesion (fnbB), enterotoxins (sea, seb, sed, seh, sej, sek, seq, ser, seu), exfoliative toxins (eta, etb, etd), island marker (splF), toxic shock syndrome toxin 1 (tst), and epidermal cell differentiation inhibitor gene (edin) were not found in any MRSA strains. All but one isolate carried adhesin (cna, C67), enterotoxin (sem, P35), and hemolysin genes (hlg-2, P68). All but one isolate lacked adhesin (map/eap, B80), enterotoxin (sec, see, sel, sep, B15), and island marker genes (bsaB, P6). Frequency of other virulence genes, such as fnbA (7/13), seo (9/13), lukF (9/13), ear (10/13), and arcA (3/13) genes, was variable.
Virulence genes that all MRSA isolates carry are highlighted in bold.
Virulence genes that are negative for all MRSA isolates: fnbB, sea, seb, sed, seh, sej, sek, seq, ser, seu, eta, etb, etd, splF, tst, edin.
Rep-PCR and plasmid profiles
Rep-PCR analysis identified 3 distinct clusters among the 13 MRSA strains (Fig. 1). Dendrogram analysis suggested that the rep-PCR profiles of chicken isolates had the highest similarity index among meat types regardless country of origin. All Korean and Spanish isolates grouped in the same cluster with >95% similarity, respectively. Plasmids (1.5–15.0 kb) were found in 12 of the 13 MRSA isolates. MRSA C67, C71, P219, P71, B5, and P6, obtained from Brazil, Canada, and Korea, had similar-sized plasmids (Fig. 2, lanes 1–4, 11–12) and other isolates carried megaplasmids measuring more than the largest size of supercoiled DNA ladder available (>15.0 kb).
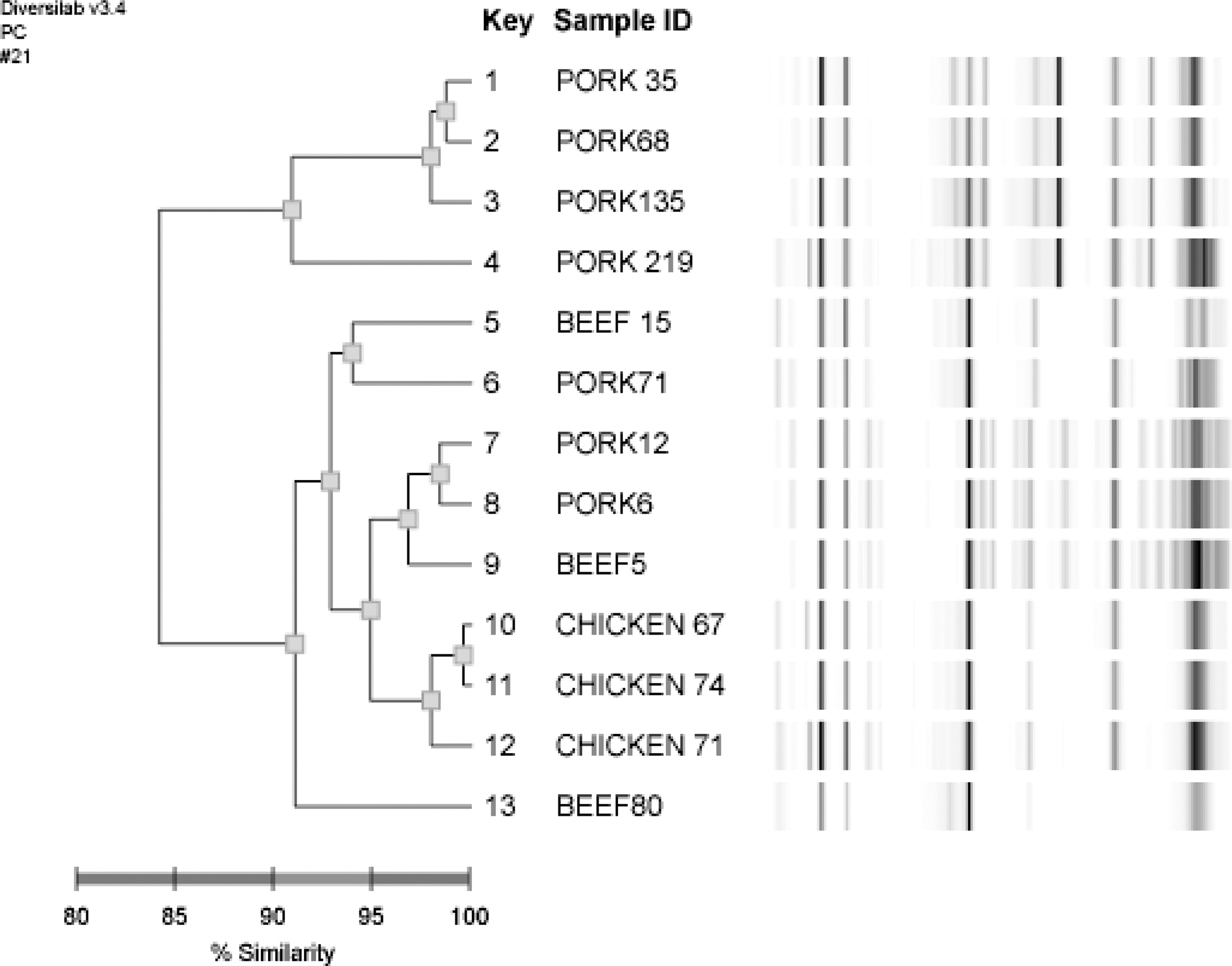
Comparison of methicillin-resistant Staphylococcus aureus strains isolated from imported and domestic meat in Korea by automated repetitive sequence-based polymerase chain reaction. The dendrogram represents the relatedness of strains based upon analyses of the amplification products using DiversiLab software (version 2.1.66).
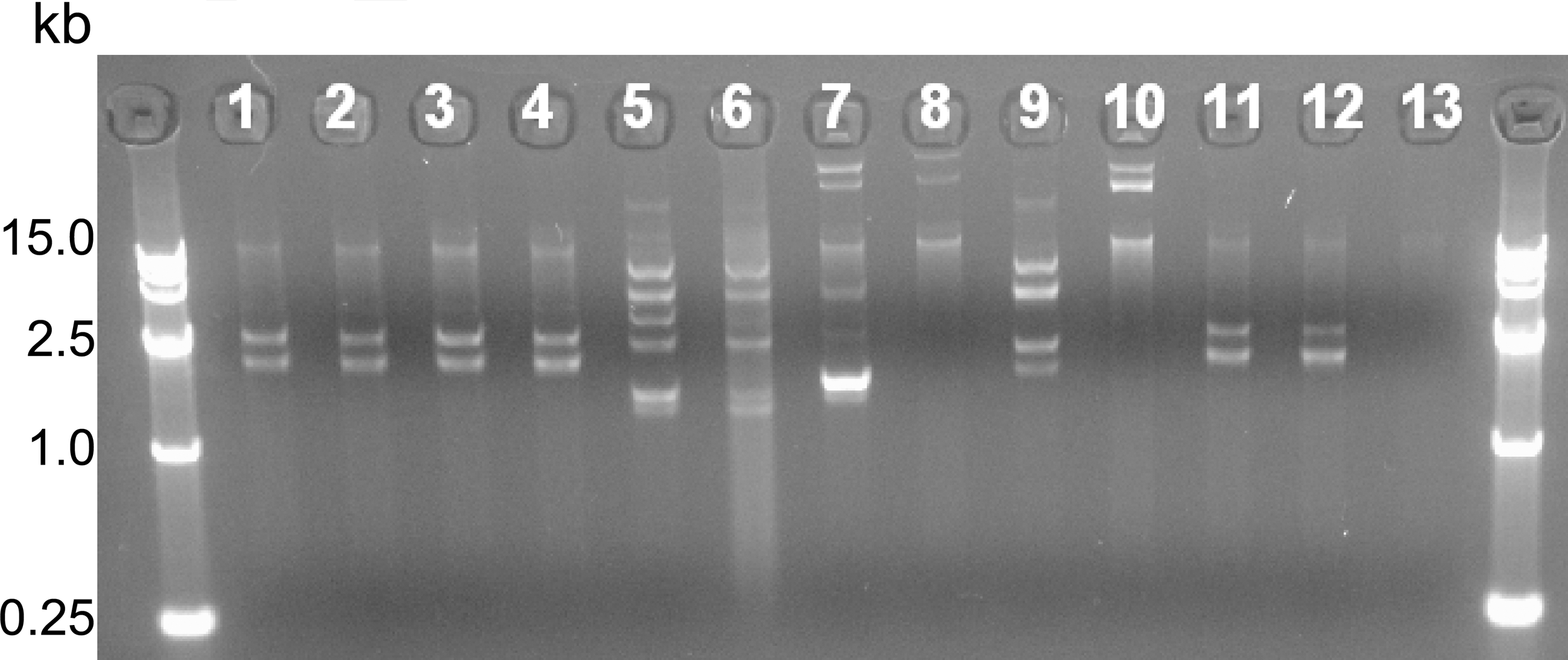
Plasmid DNA profiles of methicillin-resistant Staphylococcus aureus (MRSA) strains isolated from imported and domestic meat in Korea. First and last lanes are supercoiled plasmid DNA ladders. MRSA strains of lanes 1–13: C67, C71, P219, P71, C74, B15, P35, P68, B80, P135, B5, P6, and P12.
Discussion
ST5, ST8, ST72, and ST398 MLST types were identified from the tested samples and, unexpectedly, ST72 type was found to be a major MLST type. MRSA belonging to ST72 and SCCmecIV were isolated in a hospital in Brazil (Schuenck et al., 2009). However, our study is the first finding of the ST72-SCCmecIV MRSA clone from meat products that showed a similar antimicrobial susceptibility pattern to clinical isolates. We found that MLST and spa types of Canadian pork isolates were ST5-t002 (P71) and ST72-t148 (C74). MRSA ST5 is usually found in pigs, farm workers, and retail pork in Canada (Khanna et al., 2008; Weese et al., 2010b). However, to our knowledge, ours is the first finding of ST72 in Canadian MRSA. Furthermore, P71 was multidrug resistant to tetracycline, clindamycin, erythromycin, and quinupristin/dalfopristin, but most Canadian MRSA food isolates were reported to be highly susceptible to most classes of antibiotics (Golding et al., 2010).
Livestock-associated ST398 MRSA in Europe has been documented to be associated with human infection (Monaco et al., 2013). In our investigation, 23.1% proved to be ST398, and lukS, lukF, and lukE-lukD were positive for 2 Spanish isolates. The MRSA ST398 genotype (t2383, SCCmecV) of P135 was detected in food and clinical samples in the United States, but spa type t2383 has not been found yet (Bhat et al., 2009; Waters et al., 2011). Although S. aureus type ST398, isolated from turkey, pork, and chicken, was most common in the United States, all of them were sensitive to methicillin (Waters et al., 2011). B80 (ST8, t008, SCCmecIVA) was resistant to clindamycin, tetracycline, erythromycin, and ciprofloxacin. B80 (SCCmecIVA, ST8, t008, lukS +, and arcA +) was also very similar to USA300, except that lukF was absent.
All MRSA strains were resistant to penicillin, but only two strains harbored penicillin-resistance genes, either blaZ only or both blaZ and p480. Lee et al. (Lee et al., 2001) also reported that about 20% of penicillin-resistant staphylococci could not amplify blaZ. Several studies report that most tetracycline-resistant MRSA possess tetM gene (Trzcinski et al., 2000; Schmitz et al., 2001). However, in our work, both tetK and tetL were present in all tetracycline-resistant MRSA, whereas only half of the MRSA contained tetM. B80 was resistant to ciprofloxacin (MIC=8 μg/mL) among our collection and the combination of a Ser84 to Leu mutation in gyrA and an Asp291 to Gly in norA was found to be responsible for the resistance. All MRSA strains except P219 amplified PCR products, with three primer pairs extending from cadD-F3 to cadX-R8. Community-acquired MRSA and methicillin-sensitive S. aureus (MSSA) have exhibited similar results (Massidda et al., 2006; Takano et al., 2008).
Bbp that was positive for only highly virulent isolates (Vancraeynest et al., 2004) was present in all MRSA. MRSA isolated from humans was reported to have map/eap associated with extracellular adherence (Campbell et al., 2008). B80 also carried map/eap. SdrD and sdrE, which were positive for all our MRSA isolates, have been detected only in human clinical MRSA (Campbell et al., 2008). To the best of our knowledge, this is a new finding of MRSA strains from food that contained sdr genes. It is noteworthy that B15 harbored an uncommon type of enterotoxin, sec-see-seg-sei-sel-sem-sen-seo-sep. The coexistence of seg, sei, sem, sen, and seo in the MRSA demonstrated that these genes are within the enterotoxin gene cluster (Shukla et al., 2010). P71 and P6 isolates harbored all of lukS-lukF-lukE-lukD-lukM. No one has reported yet that any single MRSA or MSSA isolate from human or animal possesses lukS-lukF-lukE-lukD-lukM. MRSA USA300 has been reported to have bsaB (Escobar et al., 2013). Among our collection, only P6 carried the gene, but this isolate (SCCmecV, ST72, t324, lukF+ , lukS+, sek-, seq- ) did not share the same molecular profile with USA300 (SCCmecIV, ST8, t8, lukF+ , lukS+, sek+, seq+ ). Both community- and hospital-acquired MRSA strains in the United States and Spain have been reported to carry ear and efb (Shukla et al., 2010; Lim et al., 2012). They were present in 77% in most of our collection. IcaA is known to play a critical role in biofilm production in S. aureus (Satorres and Alcaraz, 2007). All of our MRSA isolates amplified an icaA PCR product of 936 bp. Additionally, compared to a reference strain, S. aureus ATCC 25923, all MRSA isolates had a great ability for biofilm formation (data not shown). V8 serine protease was positive for all of our MRSA strains. Campbell et al. (Campbell et al., 2008) reported that 91% of S. aureus isolates from patients with skin infections were positive for the V8. All of our isolates contained the efb. arcA was first reported in the whole genome sequence of USA300 MRSA and may play a role in virulence and survival (Diep et al., 2006). Until now, arcA has been identified in ST8-MSSA, ST8-MRSA-SCCmecIV, ST5-MRSA-SCCmecII, ST59-MRSA-SCCmecII, ST97-MRSA-SCCmecV, and ST1-MRSA-SCCmecIVA (Ellington et al., 2008). For the first time, we report that arcA was detected in two specific genotypes, ST72-SCCmecV (P219) and ST72-SCCmecIVA (P12).
Whereas plasmid DNA sizes of C67, C71, P219, P71, B5, and P6 isolates were similar, the antimicrobial susceptibility profiles of resistance phenotypes did not share the same profile (Fig. 2). Dendrogram analysis of the rep-PCR showed that MRSA from different countries were genetically diverse, but the poultry isolates exhibited the highest similarity index (Fig. 1).
With the rapid economic growth and free-trade agreements, Korea has recently seen a significant increase of imported meat (Shida and Hwang, 2007). Results from our study indicate that the imported meat may serve as a reservoir of multiple antimicrobial-resistant MRSA. Furthermore, these MRSA isolates could carry various toxin genes previously known only in clinical MRSA. Therefore, our study suggests that there is a need for comprehensive monitoring for MRSA within meat imported into Korea, and that people who handle meat products must put food safety precautions into practice.
Footnotes
Acknowledgments
We thank Drs. Carl E. Cerniglia, John B. Sutherland, and Fatemeh Rafii for critical review of the manuscript. This work was supported by a grant (13161MFDS923) from the Ministry of Food and Drug Safety, Korea. The views presented here do not necessarily reflect those of the U.S. Food and Drug Administration.
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.