
Abstract
Bacteria harboring cfr, a multidrug resistance gene, have high prevalence in livestock in China and might be transmitted to humans through direct contact or via contaminated food products. To better understand the epidemiology of cfr producers in the food chain, the prevalence and genetic analysis of Staphylococcus isolates recovered from pigs, workers, and meat-handling facilities (a slaughterhouse and a hog market in Guangzhou, China) were examined. Twenty (4.5%) cfr-positive Staphylococcus isolates (18 Staphylococcus simulans, 1 S. cohnii, and 1 S. aureus) were derived from pigs (16/312), the environment (2/52), and workers (2/80). SmaI pulsed-field gel electrophoresis of 26 staphylococcal strains (22 S. simulans and 4 S. cohnii), including previously reported cfr-carrying staphylococci of animal food origin, exhibited 19 major pulsed-field gel electrophoresis patterns (A–S). Clonal spread of cfr-carrying staphylococci among pigs, workers, and meat products was detected. The genetic contexts of cfr in plasmids (pHNKF3, pHNZT2, and pHNCR35) obtained from S. simulans of swine or human origin were similar to that of Staphylococcus species isolated from human clinics and animal-derived food. The cfr-carrying S. aureus strain isolated from floor swabs of the hog market was spa-type t889 and belonged to the ST9 clonal lineage. In summary, both clonal spread and horizontal transmission via mobile elements contributed to cfr dissemination among staphylococcal isolates obtained from different sources. To monitor potential outbreaks of cfr-positive bacteria, continued surveillance of this gene in animals at slaughter and in animal-derived food is warranted.
Introduction
L
The transferable gene cfr, originally identified in the pSCFS1 plasmid from a bovine S. sciuri isolate (Schwarz et al., 2000), encodes an RNA methyltransferase that modifies the adenosine at position 2503 in 23S rRNA. Cfr mediates resistance to oxazolidinones, phenicols, lincosamides, pleuromutilins, and streptogramin A, all of which are widely used for treating human and animal infections (Long et al., 2006). To date, cfr has been detected in Staphylococcus, Enterococcus, Bacillus, Macrococcus, Jeotgalicoccus, Escherichia coli, Proteus vulgaris (Shen et al., 2013), and Streptococcus suis (Wang et al., 2013) of animal origin and is also found globally in human clinical Staphylococcus and Enterococcus isolates (Shen et al., 2013). Cfr is widely disseminated among pigs farmed in China (Shen et al., 2013) and retail meat in Guangzhou, China has also tested positive for cfr (Zeng et al., 2014). This indicates that cfr-positive bacteria may find their way into humans through direct animal–human contact or contaminated meat products. In this study, we investigated the prevalence and dissemination of cfr among staphylococcal isolates from pigs, workers, and the environment of a hog market and a slaughterhouse in Guangzhou.
Materials and Methods
Sample collection and cfr detection
From March 2011 to May 2011, a total of 569 samples were collected from a hog market and a slaughterhouse in Guangzhou. The samples included 207 pig nasal swab samples collected from the hog market (no more than 5 samples were collected from pigs at the same farm), 179 pig nasal swab samples from 1 slaughterhouse, 110 human nasal swab samples from workers at the hog market and the slaughterhouse, 31 floor swab samples from the hog market, and 42 surface swab samples from pig transportation vehicles.
All samples were cultivated in brain-heart infusion broth for 18–24 h at 37°C. The cultured broth was then streaked onto plates of mannitol salt agar for 18–24 h at 37°C, and 1 isolate per sample was selected for further study. Whole-cell DNA of all isolates was extracted using previously described methods (Louie et al., 2002). A previously described polymerase chain reaction (PCR) assay for cfr detection was performed (Kehrenberg and Schwarz, 2006). Species identification of all cfr-positive strains was performed by sequencing both gap and 16S rRNA (Ghebremedhin et al., 2008). Other resistance genes, such as ermA, ermB, ermC, msrA, msrB, vga, vgaAv, fexA, mecA, and lsa(C), were also detected by PCR (Lina et al., 1999; Louie et al., 2002; Haroche et al., 2003; Kehrenberg and Schwarz, 2006; Malbruny et al., 2011).
Molecular typing
Genetic diversity of cfr-positive staphylococcal isolates was determined by performing SmaI pulsed-field gel electrophoresis (PFGE) according to previously reported protocols (McDougal et al., 2003). Seven previously described cfr-positive staphylococci, including four S. simulans and three S. cohnii isolates, collected from pork and chicken meat sold in Guangzhou in 2012, were selected for comparison (Zeng et al., 2014). Comparison of PFGE patterns was performed with BioNumerics software (Applied Maths, Sint-Martens-Latem, Belgium) using the Dice coefficient (1.5% optimization and 1.0% tolerance). Patterns were clustered using the unweighted pair-group method with arithmetic mean. A similarity cut-off of 80% was used to define a PFGE cluster (McDougal et al., 2003). Different PFGE clusters were indicated by capital letters in alphabetical order.
The cfr-carrying S. aureus strain was characterized by spa typing (Shopsin et al., 1999) and multilocus sequence typing [MLST] (Enright et al., 2000) according to previously described procedures. The spa type was assigned using the Ridom web server (
Transformation and hybridization experiments
Plasmid DNA was extracted using the E.Z.N.A. Plasmid DNA Midi Kit (Omega, Norcross, GA) following the manufacturer's instructions. The resulting plasmid DNA was transferred into the RN4220 S. aureus recipient strain by electroporation at 2.5 kV using 0.2-cm cuvettes and a Gene Pulser apparatus (Bio-Rad, Munich, Germany). Transformation experiments were performed using two randomly selected cfr-positive S. simulans strains of swine and human origin with different PFGE patterns. Transformants were selected on brain-heart infusion agar supplemented with 10 μg/mL florfenicol, and the presence of cfr was further confirmed by PCR. Southern blot hybridization was performed using plasmid DNA of the original strains to identify the location of cfr. Extracted plasmid DNA fragments were transferred to Hybond N+ membranes (Amersham Biosciences, Piscataway, NJ) and hybridized with a nonradioactively labeled cfr-specific probe using a DIG High Prime DNA Labeling and Detection Kit (Roche Diagnostics, Mannheim, Germany) according to the manufacturer's instruction.
Antimicrobial susceptibility testing
The original cfr-positive isolates, recipient strain S. aureus RN4220, and transformants were tested for their susceptibility to clindamycin, chloramphenicol, florfenicol, tiamulin, valnemulin, linezolid, vancomycin, oxacillin, tetracycline, gentamicin, ciprofloxacin, and erythromycin using the agar dilution method. In addition, isolates were investigated for resistance to quinupristin/dalfopristin (15 μg) using the agar disc diffusion method. Both susceptibility tests were performed and evaluated using protocols recommended by the VET01-S2 (CLSI, 2013a) and M100-S23 (CLSI, 2013b) of the Clinical and Laboratory Standards Institute (Wayne, PA). S. aureus reference strains ATCC 29213 and ATCC 25923 were used for quality control.
Genetic environment of the cfr gene
To identify the sequences surrounding cfr, partial nucleotide sequences of cfr-carrying plasmids were determined using an inverse PCR strategy and PCR mapping using primers corresponding to the sequences of the cfr-carrying plasmids pHK01 (GenBank accession number KC820816), pSS-01(accession no. JQ041372), and pSS-03 (accession no. JF834911; Supplementary Table S1; Supplementary Data are available online at
Nucleotide sequence accession number
The nucleotide sequences of a 7320-bp fragment of the pHNKF3 plasmid, a 9880-bp segment of pHNCR35, and a 5086-bp segment of pHNZT2 were deposited in GenBank under the accession nos. KF861984, KF861983, and KF861985, respectively.
Results
Identification of the cfr gene in Staphylococcus isolates
From a total of 569 samples, 444 Staphylococcus isolates (165 from hog market pigs, 147 from slaughterhouse pigs, 80 from workers, 29 from surface swabs from pig transport vehicles, and 23 from hog market floor swabs) were recovered. Among them, cfr was present in 20 (4.5%) Staphylococcus isolates obtained from pigs (n=16), the environment (n=2), and workers (n=2; Table 1). S. simulans (n=18) was the predominant species isolated. One isolate each of S. cohnii from pig and S. aureus from hog market floor swabs were also isolated (Table 2).
C, chromosome; P, plasmid.
MIC, minimum inhibitory concentration; CLI, clindamycin; CHL, chloramphenicol; FFC, florfenicol; TIA, tiamulin; VAL, valmulin; VAN, vancomycin; LZD, linezolid.
Q-D, quinupristin/dalfopristin. The antimicrobial susceptibility of quinupristin/dalfopristin was determined by the agar disk-diffusion method.
OXA, oxacillin; TET, tetracycline; GEN, gentamycin; CIP, ciprofloxacin; ERY, erythromycin. All the isolates are susceptible to vancomycin.
Molecular typing
PFGE analysis of the 26 cfr-positive staphylococci (22 S. simulans and 4 S. cohnii) obtained from pigs, the environment, humans, and animal food products revealed that the 22 S. simulans isolates from diverse sources exhibited 15 different PFGE patterns (A–O), whereas the 4 S. cohnii isolates of swine and animal food origin exhibited 4 different PFGE patterns (P–S; Fig. 1; Supplementary Fig. S1). Thus, cfr dissemination might not be due to the spread of a specific Staphylococcus clone. However, some isolates obtained from different sources had similar PFGE patterns (Fig. 1). Interestingly, one porcine isolate and one strain of human origin showed two similar patterns (C1 and C2) with a difference of a single band (Fig. 1). In addition, one pork isolate (TLD22) and three pig isolates showed three similar patterns (B1, B2, and B3). The cfr-positive S. aureus strain BKED was shown to belong to spa-type t889 and clonal lineage ST9.
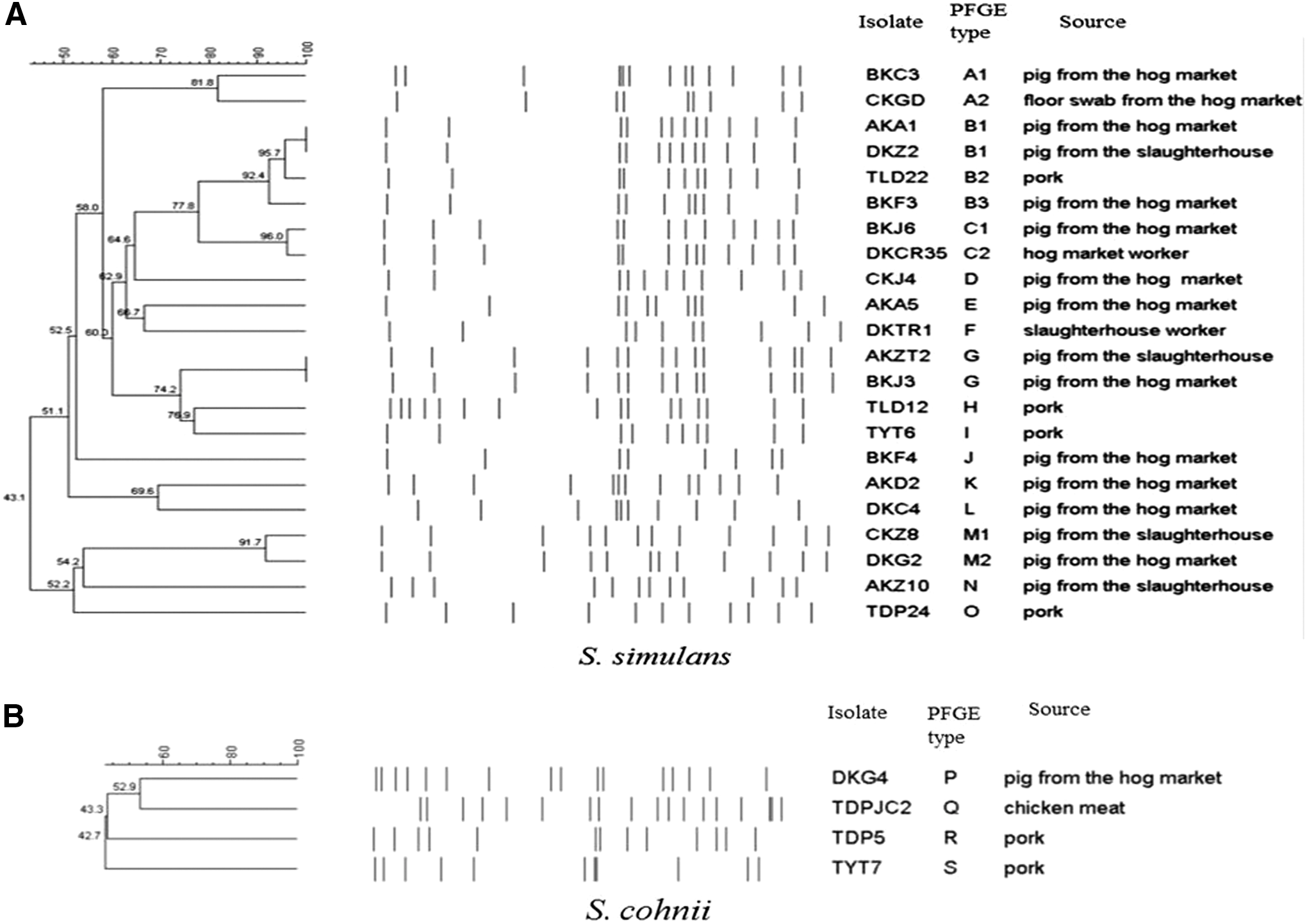
Dendogram of the pulsed-field gel electrophoresis profile of SmaI-digested genomic DNA of 25 cfr-positive staphylococcal strains recovered from pigs, environment, workers, and retail meat.
Localization of cfr
Southern blot hybridization indicated that cfr was harbored in plasmid DNA (17 isolates) and chromosomal DNA (3 isolates). In addition, plasmid DNA was obtained from S. simulans DKCR35 of human origin and from S. simulans AKZT2 of swine origin and then successfully transferred to S. aureus RN4220. Plasmids obtained from resultant transformants were designated as pHNCR35 (human origin) and pHNZT2 (swine origin).
Antimicrobial resistance and resistance determinants
Antimicrobial susceptibility testing showed that all 20 cfr-positive isolates were resistant to clindamycin, tetracycline, and erythromycin and showed elevated minimum inhibitory concentrations for chloramphenicol, florfenicol, tiamulin, and valnemulin; however, isolates were susceptible to vancomycin. Of the 20 cfr-positive isolates, 3 were considered resistant and 10 were shown to have intermediate resistance to quinupristin/dalfopristin. Additionally, nine (45%), 14 (70%), nine (45%), and 14 (70%) isolates exhibited resistance to linezolid, gentamicin, oxacillin, and ciprofloxacin, respectively.
The two transformants (S. aureus RN4220:pHNCR35 and RN4220:pHNZT2) exhibited resistance to clindamycin, chloramphenicol, and linezolid and showed elevated minimal inhibitory concentrations of florfenicol, tiamulin, and valnemulin compared with the S. aureus recipient strain RN4220. For the assay using quinupristin/dalfopristin discs, the growth inhibition zone diameters of transformants were 7–8 mm smaller than those of the recipient RN4220. This finding indicated cfr prevalence and expression in transformants (Table 2). In addition, the two transformants were resistant to erythromycin, but susceptible to tetracycline, ciprofloxacin, and oxacillin. Transformants carrying the plasmid pHNZT2 also showed resistance to gentamicin.
The 20 cfr-positive isolates and the transformant RN4220:pHNZT2 carried fexA. In addition, all of the 20 cfr-positive isolates and the 2 transformants carried 1–3 erythromycin resistance genes, ermA, ermB, and ermC (Table 2). Five strains harbored vga or vgaAv. All of the nine oxacillin-resistant isolates harbored mecA and were therefore considered to be methicillin-resistant coagulase-negative staphylococci.
Genetic environment of cfr in the plasmids pHNKF3, pHNCR35, and pHNZT2
The genetic structure surrounding cfr was assessed using inverse PCR combined with PCR mapping. A 7.3-kb cfr-containing fragment of plasmid pHNKF3 carried by S. simulans BKF3 was sequenced. Sequencing results showed that a Tn4001-like transposon, including the aminoglycoside resistance gene aacA-aphD flanked by an incomplete IS256 and IS256-like element, was upstream of cfr. In addition, another IS256-like element was present in the downstream region of cfr. This 7.3-kb cfr-containing fragment showed 99% identity to plasmid pSS-01 (accession no. JQ041372) obtained from an S. cohnii strain recovered from a pig in Shandong, China, and plasmid p7LC (accession no. JX910899) obtained from a human S. epidermidis isolate from the United States (Fig. 2).
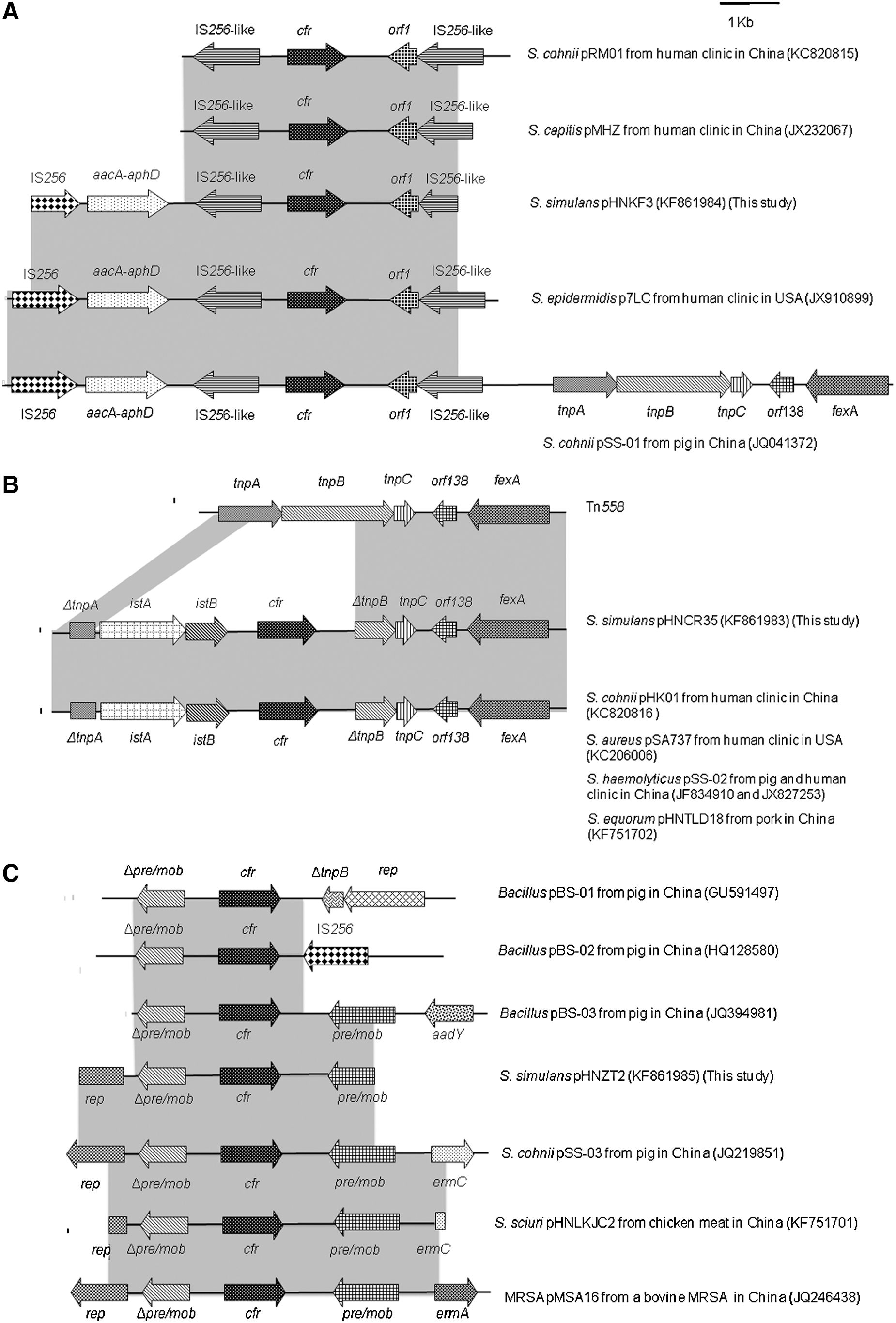
Genetic environment of cfr in plasmids (pHNKF3, pHNCR35, and pHNZT2) and their structural comparison with other similar plasmids. The arrows indicate the positions and directions of the transcription of the genes. Regions of >98% homology are shaded in gray. Δ indicates a truncated gene. 1-kb distance scale is displayed in the upper right corner of Fig. 2A.
In plasmid pHNCR35 from human S. simulans, a 9880-bp region encompassing cfr was sequenced, and a Tn558 variant was identified. The segment containing an IS21-558 insertion and cfr was integrated into the Tn558 element, replacing parts of transposase genes tnpA and tnpB. This 9.9-kb region containing cfr on plasmid pHNCR35 was identical to the corresponding region of plasmids pHK01 (accession no. KC820816) found in one S. cohnii strain from a Chinese patient, pSA737 (accession no. KC206006) from clinical linezolid-resistant S. aureus in the United States, pHNTLD18 (accession no. KF751702) from the TLD18 S. equorum strain of animal food origin in Guangzhou, China, pSS-02 (accession no. JX827253) carried by two S. cohnii and one S. haemolyticus isolates from swine (Wang et al., 2012), and a clinical linezolid-resistant S. haemolyticus strain from China (Fig. 2).
A 5086-bp segment carrying cfr of the plasmid pHNZT2 was also obtained and sequenced to discover an incomplete rep, Δpre/mob upstream of cfr and a partial pre/mob downstream of cfr. This segment was completely identical to the corresponding region of plasmids pSS-03 (accession no. JQ219851) carried by an S. cohnii strain of swine origin, pMSA16 (accession no. JQ246438) extracted from a bovine MRSA strain, and pHNLKJC2 (accession no. KF751701) from an S. sciuri isolate from animal food in Guangzhou (Fig. 2).
Discussion
The distribution cfr among staphylococcal isolates simultaneously collected from animals at slaughter, the surrounding environment, and people in close contact with them has not been previously studied. We screened 444 staphylococcal isolates collected from pigs, workers, and the environment in a hog market and a slaughterhouse to assess cfr prevalence. The data indicated that cfr was present in 5.1%, 3.8%, and 2.5% of staphylococcal isolates obtained from pigs, the environment, and workers, respectively. These data may provide lower estimates for cfr prevalence in pigs at slaughter because the staphylococcal isolates used in this study were randomly selected without any antimicrobial selection. A previous study showed that 22.1% of porcine staphylococcal isolates (screened with selective media containing 10 μg/mL florfenicol) obtained from swine farms in China harbored cfr (Wang et al., 2012). The high prevalence of Cfr-producing Staphylococcus among pigs may be responsible for the high cfr prevalence (18.6%) in retail meat samples as reported in our previous study (Zeng, 2014).
The majority of the cfr-positive staphylococcal isolates were clonally unrelated. However, the high similarity among PFGE profiles of S. simulans isolates obtained from different sources indicated that clonal spread of cfr-carrying S. simulans had occurred among pigs, humans, the environment, and animal-derived food. The cfr flanking sequences in plasmids from porcine or human S. simulans strains characterized in this study were highly homologous to those that were previously obtained from humans, animals, and foods, indicating that horizontal transmission mediated by mobile genetic elements had occurred and may be the primary cfr dissemination mechanism among staphylococcal isolates. The transmission of cfr-carrying plasmids between staphylococci from pigs and humans has already occurred (Cui et al., 2013). A previous study indicated that ingestion of contaminated meat might result in the dissemination of resistant bacteria between food, animals, and humans (Leverstein-van Hall, 2011). The emergence and dissemination of cfr-carrying staphylococcal strains and plasmids among pigs at slaughter, retail meat, and the environment is worrisome and requires continued vigilance, given the possibility that cfr can be transferred from animals to humans through the food chain and via fomites and potentially induce further human-to-human spread in the community (Geenen et al., 2010).
The S. aureus strain BKED belonged to spa-type t889 and was assigned to the MLST-type ST9. ST9 is frequently associated with pigs. Although ST9 is a minor S. aureus MLST in Europe, Canada, and the United States, it is identified as the predominant sequence type in China (Cui et al., 2009; Boost et al., 2012). To date, among all cfr-positive S. aureus isolated so far, only one porcine methicillin-susceptible S. aureus ST9 collected from Germany and one bovine MRSA ST9 collected from China have been reported (Shen et al., 2013). Previous studies indicate that there is a significant risk of colonization by S. aureus ST9 in pig-handlers (Cui et al., 2009). Several cases of human infections with MRSA ST9 have been reported in Miami, FL (Chung et al., 2004) and Guangzhou, China (Liu et al., 2009). The ST9 S. aureus strain analyzed in this study might have been colonized in pigs. Although the origin of this isolate remains unclear, the transfer of cfr-carrying S. aureus ST9 from swine to humans through a contaminated environment and further spread among humans cannot be precluded since the most common route of S. aureus transmission from animals to humans is through direct contact with animals and their surrounding environment (Geenen et al., 2010).
Our findings indicated that cfr dissemination among staphylococcal isolates obtained from different sources occurred via both clonal spread of cfr-positive strains and through horizontal transfer mediated by mobile elements. These data indicate the potential for cfr transfer from animals to humans through food or fomites. Thus, continuous surveillance of cfr dissemination in staphylococcal isolates obtained from animals and food is needed to safeguard public health.
Footnotes
Acknowledgments
This study was supported in part by the National Key Basic Research Program of China (No. 2013CB127200), the Program for Changjiang Scholars and Innovative Research Team in University (IRT13063), the Guangdong Natural Science Foundation (No. S2012030006590), the Foundation for the Author of Guangdong Excellent Doctoral Dissertation (No. sybzzxm201225), and the fund for Training of PhD Students from the Ministry of Education of China (201044041100).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.