
Abstract
Bulk tank milk from 80 dairy farms located in the West Java Region of Indonesia was analyzed for the presence of extended-spectrum β-lactamase (ESBL)–producing Enterobacteriaceae. Isolates from seven dairy farms were ESBL positive, and all were identified as Klebsiella pneumoniae. The isolates showed ESBL-characteristic antibiotic resistance patterns. Further analysis revealed that all K. pneumoniae isolates harbored the bla SHV gene, and two isolates were additionally positive for the bla TEM-1 and bla CTX-M-15 genes. Isolates from different farms were clonally diverse according to macrorestriction analysis. The results indicate that the relatively high frequency of ESBL-producing K. pneumoniae in bulk tank milk implies the risk that milk is both a source of local exposure and a vector contributing to the supraregional spread of antibiotic-resistant bacteria by trade.
Introduction
W
Milk payment in Java is dependent on quality parameters such as protein, fat, and total bacteria (maximum 1 million per milliliter), but stringent routine testing for inhibitors (antibiotic residues) is only implemented in some regions (Morey, 2011; Nugroho, 2012). In some regions, antibiotic residue testing of milk is done only sporadically. The lack of efficient veterinary drug control increases the risk of drug residues in milk, and enhanced selection of β-lactam-resistant bacteria may be the result (Sudarmono, 2013; Morey, 2011).
Extended-spectrum β-lactamases (ESBL) are enzymes produced by bacteria that can degrade and confer resistance to some of the most commonly used group of antibiotics, the β-lactams, which comprises the penicillins, the cephalosporins, and the monobactams (Falagas and Karageorgopoulos, 2009; Ewers et al., 2011). Recent studies in Indonesia indicated that antibiotic resistance patterns from several hospital isolates have spread, especially in Gram-negative bacteria such as Escherichia coli, Klebsiella pneumoniae, Pseudomonas aeruginosa, and Acinetobacter baumannii. Most of these were resistant both to third-generation cephalosporins and to fluoroquinolones (Moehario et al., 2009; Kuntaman et al., 2011; Radji et al., 2011; Karuniawati et al., 2013).
K. pneumoniae, in particular ESBL-producing strains, is an important cause of nosocomial infections in humans worldwide (Podschun and Ullmann, 1998; Marra et al., 2006; EFSA, 2011; Sumer et al., 2014). For Indonesia, an increase of ESBLs within clinical isolates of K. pneumoniae has been reported in recent years (Herwana et al., 2008, Kuntaman et al., 2011; Parwati et al., 2012; Saharman and Lestari, 2013). In dairy farming, K. pneumoniae is one of the most important causative agents for clinical mastitis in lactating cows, together with Escherichia coli (Hogan and Smith, 2003), and β-lactam antibiotics are the drugs of choice for treatment (Carattoli, 2008; Smet et al., 2010). Improper and excessive use of intramammary or parenteral β-lactam antibiotics may provoke bacterial adaptation. In Indonesia, there are few restrictions concerning the use of antibiotics, neither in human medicine nor in veterinary medicine. Thus, it may be suspected that the rapidly increasing incidence of multidrug-resistant bacteria could be a result of the unconscious and extensive use of antibiotics in this country (Sudarmono, 2013).
So far, no information concerning the dimension of the risk from emerging antibiotic-resistant bacteria in milk is available for West Java. Therefore, the aim of this study was to assess the frequency of ESBL-producing Enterobacteriaceae in bulk tank milk samples collected from dairy farms located in the West Java Region of Indonesia.
Materials and Methods
Milk samples
A total of 80 randomly selected raw milk samples, representing bulk tank milk of dairy farms located in the West Java region of Indonesia, were collected from November to December 2011. Farms were either smallholders (3–8 lactating cows) or medium-sized dairy farms (10–15 lactating cows), and none of these had on-site milk-cooling facilities. One sample per farm was collected directly from the milk tank. All samples were then immediately put into an ice-refrigerated box (<8°C), and reached the laboratory within 1–4 h after collection.
Microbiological analysis
For screening of ESBL-producing bacteria, 10-mL milk samples were supplemented with cefotaxime (Sigma-Aldrich, Munich, Germany) at a final concentration of 1 mg/L and incubated for 24 h at 37°C. This approach is an adaptation of a procedure recently recommended for ESBL isolation from other matrices (EFSA, 2011). Then 0.1 mL was streaked onto MacConkey agar (Oxoid, Wesel, Germany) containing 1 mg/L cefotaxime and incubated at 37°C for 24 h. For each morphologically distinct type of colony, 1 isolate was selected and subcultivated on tryptone soy (CASO) agar (Oxoid) at 37°C for 24 h.
All isolates were tested for colony characteristics, Gram staining, and oxidase reaction. Gram-negative and oxidase-negative colonies were further characterized by using standard bacteriological procedures, including colony morphology, motility, lactose fermentation, malonat utilization, the production of catalase and indole, and by API20E rapid test kit (BioMérieux, Marcy l'Etoile, France). Isolates were stored in trypticase soy broth containing 20% glycerol at −20°C until further workup.
ESBL confirmation and antimicrobial susceptibility
All cefotaxime-resistant, and oxidase-negative, isolates were screened for ESBL production by the combined-disk method according to the Clinical Laboratory Standards Institute guidelines (CLSI, 2013). Zones of inhibition were determined for each isolate, using antibiotic disks each containing 30 mg of cefotaxime, ceftazidime, or cefpodoxime, either alone or in combination with 10 mg of clavulanic acid (MAST Group Ltd., Reinfeld, Germany). Results were confirmed in a microdilution assay (CLSI, 2013) by using the Micronaut-S β-lactamase VI test plates (Merlin Diagnostika, Bornheim-Hersel, Germany), which claim the detection of isolates showing a cephalosporinase phenotype (ESBL, AmpC) or a carbapenemase phenotype (K. pneumoniae carbapenemase [KPC], metallo-β-lactamase [MBL]). The isolates in which ESBL production was confirmed by phenotypical methods, the minimum inhibitory concentration (MIC) for 11 antimicrobials commonly used to treat mastitis (Table 1), were determined by the CLSI broth microdilution method using a custom-designed microtiter panel (Merlin Diagnostika). MICs were interpreted according to CLSI guidelines. A susceptible strain (E. coli ATCC 25922) and a bla SHV-positive strain (K. pneumoniae ATCC 700603) were included to monitor the performance of ESBL detection agents and as quality controls for all susceptibility tests.
For all isolates, identical MICs were obtained with the following substances: cefoxitin (<4), ampicillin (>16), cefazolin (>32), cefoperazon (>16), erythromycin (<4), oxacillin (>4), penicillin G (>8), pirlimycin (>4).
Number and location of individual farm; FB, Fapet-Bogor; C, Cipanas; KB, Kunak-Bogor.
For all isolates, identical T-value (1.0) was obtained.
3-APB, 3-aminophenylboronic acid; CEP, cefepime; CMC, cefepime (with/without) clavulanic acid; CAZ, ceftazidime; CZB, ceftazidime (with/without) 3-APB; CZC, ceftazidime/clavulanic acid; CTX, cefotaxime; CTB, cefotaxime/3-APB; C/C, cefotaxime/clavulanic acid; MER, meropenem; MEB, meropenem/3-APB; MEE, meropenem/clavulanic acid; AMC, amoxicillin/clavulanic acid 2:1; CEQ, cefquinom; K/C, kanamycin/cephalexin; MAF, marbofloxacin.
Characterization of β-lactamases by polymerase chain reaction (PCR)
ESBL-positive isolates were further analyzed for the presence of bla genes of the ESBL subtypes TEM, SHV, and CTX-M (group 1, 2, 8, 9, or 25) by PCR using primers and conditions as previously reported (Pitout et al., 1998; Batchelor et al., 2005; Woodford et al., 2006). Bacterial DNA was isolated with the DNeasy blood and tissue kit (Qiagen, Hilden, Germany) according to the manufacturer's recommendations. Two strains, K. pneumoniae ATCC 700603 (harboring a bla shv gene), and K. pneumoniae isolate (harboring both blactx -m and blatem genes) from the strain collection of the chair of dairy science, were used as standard ESBL-positive strains. A non-ESBL-producing organism (E. coli ATCC 25922) was used as negative control. PCR products were determined by electrophoresis in a 2% agarose gel (Biozym, Hessisch-Oldendorf, Germany). The molecular marker GeneRuler 100-bp DNA ladder (MBI Fermentas, St. Leon-Roth, Germany) was used.
Sequencing of bla genes
The ESBL-encoding genes bla
TEM, bla
SHV, and bla
CTX-M of the ESBL-positive isolates were amplified with primers and PCR conditions as described previously (Pitout et al., 1998; Batchelor et al., 2005). Resulting amplicons were purified using the PCR Purification Kit (Qiagen). Sequencing was performed at SeqLab (Goettingen, Germany). Results were evaluated using the BLAST algorithm available at
Analysis of chromosomal DNA restriction patterns by pulsed-field gel electrophoresis (PFGE)
All ESBL-positive isolates were characterized after digestion of the chromosomal DNA with the restriction enzyme XbaI by PFGE using the Chef-Dr II pulsed-field electrophoresis system (Bio-Rad, Munich, Germany) according to the PulseNet standard protocol for Gram-negative bacteria by the Centers for Disease Control and Prevention. A dendrogram was created using the Bionumerics software package version 5.1 (Applied Maths, Kortrijk, Belgium), with the DICE coefficient and the unweighted-pair group method with arithmetic means with a band position tolerance of 1%.
Results and Discussion
After the initial screening of bulk milk, a total number of 20 morphologically different, cefotaxime-resistant colonies from 16 milk samples were obtained on cefotaxime-supplemented MacConkey agar plates. Concerning the origin of ESBL-suspect isolates, there was no obvious geographic pattern; all regions under study yielded ESBL-suspect samples. The screening method, employing pre-enrichment of ESBL from milk in the presence of cefotaxime, has been successfully used for other matrices (EFSA, 2011) and consistently yielded positive results for inoculated milk samples at approximately 1 colony-forming unit (CFU)/mL (data not shown). The validity of the selective isolation method was routinely determined by recovery experiments, in which either raw milk or ultra-high-temperature-processed milk was inoculated with one of four different ESBL-positive strains of Enterobacteriaceae (K. pneumoniae ATCC 700603 and three E. coli from our Institutes' strain collection). It was therefore considered as sufficient for the rapid isolation of ESBL-suspect Enterobacteriaceae for further identification.
The isolates were then biochemically identified as A. baumannii (n=9), K. pneumoniae (n=8), Enterobacter cloacae (n=2), and Enterobacter aerogenes (n=1). Expression of ESBL was considered a threefold concentration decrease in the MIC (≥2 mg/mL) of either cephalosporins in the presence of clavulanic acid compared to its MIC when tested alone (MIC ratio≥4) (Table 1).
ESBL production, however, was phenotypically confirmed only in the K. pneumoniae isolates from seven different bulk milk tanks. For three Enterobacter spp. isolates, the production of another β-lactamase (AmpC) could be affirmed. The nine isolates of A. baumannii were negative for ESBL- and AmpC-production. One K. pneumoniae isolate per sample was further characterized, except for sample IM-12 from which two K. pneumoniae isolates with different morphology were selected. The MICs of the eight ESBL-positive K. pneumoniae isolates for various antibiotics are summarized in Table 1. All isolates were resistant to many β-lactams of importance in veterinary mastitis therapy (penicillins, cefquinome, cefazolin, cefoperazone), as well as to lincosamides (pirlimycin) and macrolides (erythromycin). Although the members of the genus Klebsiella can on occasion contain different resistance markers (Severin et al., 2010; Calbo et al., 2011), the broad resistance patterns of these isolates are remarkable.
Two isolates (IM-35 and IM-36) were simultaneously resistant to marbofloxacin, kanamycin/cephalexin, and amoxicillin/clavulanic acid. One isolate (IM-36) showed a high resistance to meropenem, a carbapenem antibiotic used in human medicine. This isolate was phenotypically confirmed to produce MBL enzymes and KPC, although carbapenem resistance and the encoding genes were not further characterized. Carbapenem resistance has been increasingly reported among Enterobacteriaceae, often isolated from clinical specimens, mainly attributed to MBL and KPC (Nordmann et al., 2009).
Based on PCR analysis, all eight isolates harbored SHV genes (Fig. 1). When these were sequenced, three were identified as SHV-2a, two as SHV-36, and one each as SHV-1, SHV-11, and SHV-28, respectively. Two isolates (IM-35 and IM-36), which were SHV-28 and SHV-11 positive, respectively, both co-produced TEM-1 penicillinase and CTX-M-15. Although the SHV group of genes has most frequently been reported for ESBL-producing K. pneumoniae isolates, co-occurrence of TEM and SHV enzymes has also been described for ESBLs from dairy herds (Hammad et al., 2008; Locatelli et al., 2009). More recently, a rapidly growing number of reports on positive results for the third group of ESBL, the CTX-M enzymes, have been observed (Woodford, 2010). CTX-M-positive Enterobacteriaceae isolates have been found both in companion animals and in livestock (Carattoli, 2008; Smet et al., 2010; Ewers et al., 2011). CTX-M-15 is considered to have spread worldwide, especially in Europe (Carattoli, 2008; Rogers et al., 2011), and to be the most common source of resistance to broad-spectrum cephalosporins in E. coli, and to a lesser extent in K. pneumoniae (Poirel et al., 2013). A relative increase of the percentage of CTX-M-15-positive isolates in sick animals, as compared to healthy animals, has been pointed out by Smet et al. (2010).
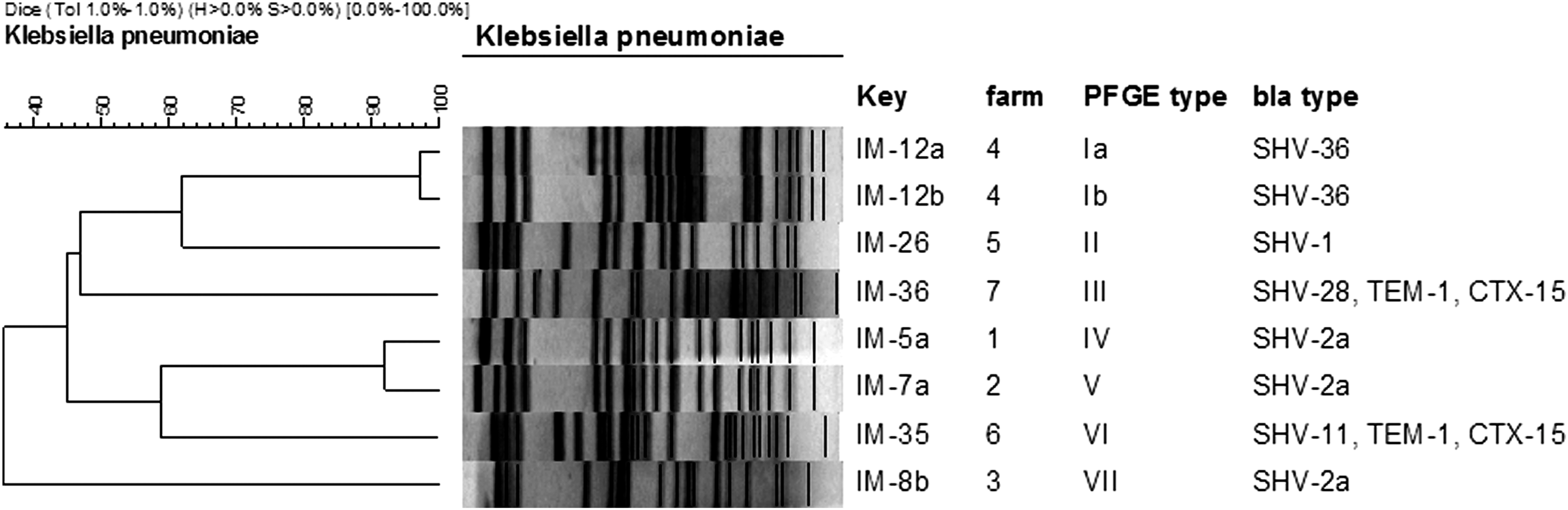
Pulsed-field gel electrophoresis (PFGE) pattern of eight Klebsiella pneumoniae isolates from seven different farms in the West Java region of Indonesia, digested with the XbaI restriction enzyme. The dendrogram was constructed with the BioNumerics software 5.1 (Applied Maths NV, Sint-Martens-Latem, Belgium) choosing the Dice coefficient setting both tolerance and optimization at 1%. The horizontal scale on the left side (100–40) indicates the level of similarity in percent among fingerprints. The 95% of similarities indicates the minimal levels for defining clusters and subtypes, respectively.
The two CTX-M-15 isolates, which were both from farms in the Cipanas region, but were otherwise unrelated to each other, indicate that this type of ESBL-producing K. pneumoniae requires specific attention. Strikingly similar findings were reported in clinical isolates from Javanese hospitals. For clinical isolates collected at two governmental teaching hospitals in Java, Indonesia, E. coli and K. pneumoniae were the dominant Enterobacteriaceae, and CTX-M-15 was the most frequent ESBL type in both species. For K. pneumoniae isolates, the bla SHV-type (54.3%) and the bla CTX-M type enzymes were each detected in more than 50% of all isolates (Severin et al., 2010, 2012). Studies from other countries confirm the importance of bla CTX-M genes in K. pneumoniae (Locatelli et al., 2010; Dahmen et al., 2013; Ohnishi et al., 2013; Schmid et al., 2013; Randall et al., 2014).
Although a direct linkage between ESBL-producing K. pneumoniae in bulk tank milk and in hospitals in Java seems to be unlikely, further investigations on the possible interrelation or interaction between both environments via milk production and distribution should be done in the future, either to confirm or to exclude implications of dietary ESBLs on human health.
PFGE analysis of ESBL-producing K. pneumoniae isolates from the seven different dairy farms yielded a markedly different pulsotype for each bulk milk tank. This indicates that the ESBL isolates have contaminated the bulk milk independently, each one in its own environment (Fig. 1). In contrast, the two isolates obtained from the same bulk tank milk sample (IM-12a and IM-12b) showed a closely related restriction profile. Whether or not this is the result of a clonal spread of K. pneumoniae–producing ESBLs in this farm remains unclear. Members of the genus Klebsiella are frequently isolated from environment and food samples, particularly from cattle with mastitis, soil, feces, and water among several sources within a dairy herd, and, together with E. coli, are probably the most important raw milk–related Enterobacteriaceae worldwide (Paulin-Curlee et al., 2008; Locatelli et al., 2010; Verbist et al., 2011).
Nevertheless, it remains remarkable that all ESBL producers were K. pneumoniae and not E. coli. On a worldwide basis, E. coli and nontyphoidal Salmonella are by far the most frequently reported ESBL producers in food-producing animals and food of animal origin in general, and specifically in cattle (Smet et al., 2010; EFSA, 2011). Although ESBL enzymes are frequently found in K. pneumoniae, this species has a small share in the overall number of ESBLs in Enterobacteriaceae recovered from food-producing animals or food of animal origin (EFSA, 2011; Dahmen et al., 2013). On the other hand, our results are not fully unprecedented. For bovine mastitis isolates from Japan, monitored over a period of 5 years, Ohnishi et al. (2013) reported that CTX-M-2–positive K. pneumoniae alone accounted for 63% of all ESBLs, and other Klebsiella spp. added another 12%. In contrast, ESBL-producing E. coli were found with a frequency of only 20%. Locatelli et al. (2010) studied isolates of K. pneumoniae from Italian mastitis cases and also found a high number of ESBLs, with bla SHV and bla TEM being the predominant genes.
Because our study is the first one for ESBLs in bulk tank milk from a tropical country, this could indicate that there is a different situation for Indonesia than has been reported for dairy environments from other countries, in particular European countries. So far, most studies dealing with ESBL-producing Enterobacteriaceae in cattle and bovine products had a primary focus on E. coli (Smet et al., 2010; Schmid et al., 2013), and this may have contributed to a relative underestimation of the relevance of K. pneumoniae.
Most of the domestically produced milk in Indonesia is thermally processed in dairy plants (Morey, 2011; Vanzetti et al., 2013). Such products should be essentially free from Enterobacteriaceae, if recontamination can be avoided. However, some of the bulk milk is pasteurized locally, using high-temperature short time (HTST, 72–75°C, 15–30 s) conditions (Slette and Meylinah, 2012; Vanzetti et al., 2013). Considering the risk of exceptionally high total bacteria numbers in bulk tank milk (>106 CFU/mL) in Indonesia, it is questionable whether or not HTST treatment is always sufficient, under real-life conditions, to fully eliminate Enterobacteriaceae. Therefore, pasteurized milk could at least be an occasional source of exposure with ESBLs.
If ESBL-producing K. pneumoniae from raw milk could find habitats at the farm site, at the KUDs, and in the dairy processing plant, this could contribute to a spread of resistant isolates not only among all those working in such environments. Furthermore, this presents a permanent risk of recontamination of pasteurized milk with these bacteria.
In conclusion, the relatively high frequency (8.75%) of ESBL-producing K. pneumoniae in raw bulk tank milk from West Java imply the risk that milk is both a source of local exposure and a vector contributing to the supraregional spread of antibiotic-resistant bacteria by trade.
Footnotes
Acknowledgments
The technical expertise of Cornelia Borchardt and Annette Schaus-Früauff is gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.