
Abstract
Salmonella enterica subsp. enterica serovar Enteritidis (Salmonella Enteritidis) is a major pathogen responsible for causing the largest number of sporadic cases and outbreaks of human salmonellosis worldwide. In this study, an outbreak of Salmonella Enteritidis involving 112 cases in Ningbo, China was investigated with a combination of genotypic subtyping methods and phenotypic analysis. The pulsed-field gel electrophoresis and multilocus variable-number tandem-repeat analysis profiles showed that most of the outbreak clinical isolates (22/23) were indistinguishable from each other and were identical to the isolates obtained from implicated mousse cakes, demonstrating that this outbreak of gastroenteritis was caused by Salmonella Enteritidis–contaminated mousse cakes. Moreover, all isolates, irrespective of source, had an identical antibiotic susceptibility pattern. Five virulence-associated genes in Salmonella pathogenicity islands and the plasmid-associated virulence genes spvB/C were present in both the food and clinical isolates. Importantly, all of these isolates can survive well under low-temperature treatment, indicating that manufacturers of foodstuffs with raw ingredients (not subjected to thermal processing) should use an effective approach to prevent or eliminate the microbial hazards to public health.
Introduction
S
In this study, all isolates from this outbreak were subtyped by pulsed-field gel electrophoresis (PFGE) and multilocus variable-number tandem-repeat analysis (MLVA), to track and to confirm that this outbreak was associated with contaminated mousse cakes. Since shaping in the refrigerator instead of traditional baking is used during the production of mousse cakes, cold tolerance of these isolates may have been a contributing factor in this outbreak, allowing the organism to persist or grow to infectious levels. Therefore, low-temperature stress tolerance was determined in this study. Also, these isolates were tested for their antibiotic resistance, virulence genes, and plasmid replicon types, to identify the pathogenic factors and potential hazards of this outbreak-associated Salmonella Enteritidis.
Materials and Methods
Salmonella Enteritidis isolates
A total of 31 Salmonella Enteritidis strains were isolated from a mousse cake–related outbreak that occurred from May 30 to June 1, 2013 in Ningbo, China. The strain associated with this outbreak consisted of 8 isolates from implicated mousse cakes (essential ingredients: eggs, butter, sugar, cream, and other flavorings; key production process: all the ingredients are mixed well and shaped in the refrigerator for at least 8 h; storage recommendations: 0–4°C for 1–2 d), and 23 isolates from 23 patients among the 112 cases who ate these cakes in the period of the outbreak and showed clinical symptoms of gastroenteritis (Table 1). It was noted that the positive rate of Salmonella Enteritidis detection among these 112 patients was only 80% because some patients had taken medicine before sampling. Moreover, the patients were sent to different hospitals, some of which failed to keep all the samples. Therefore, only 23 clinical isolates were obtained for subsequent study.
Significantly different from the survival rate of Salmonella Enteritidis reference strain ATCC 13076 as determined by Dunnett's test (p<0.05).
ND, not detected; +, positive result; –, negative result.
Antimicrobial susceptibility testing
Antimicrobial susceptibility was evaluated using the disk-diffusion method according to the criteria established by the Clinical and Laboratory Standards Institute (CLSI, 2013). The following antimicrobial agents, which represent five common types of antibiotics used to inhibit or inactivate Salmonella Enteritidis, were tested: chloramphenicol (30 μg), gentamicin (10 μg), kanamycin (30 μg), doxycycline (30 μg), norfloxacin (10 μg), nalidixic acid (30 μg), ceftazidime (30 μg), ciprofloxacin (5 μg), and trimethoprim–sulfamethoxazole (1.25/23.75 μg). Escherichia coli ATCC 25922 with known antibiotic susceptibility was used as a control strain.
Resistance to low-temperature stress
To determine the survival of a total of 31 Salmonella Enteritidis isolates at low temperature (–20°C and 4°C), the method described by Huang et al. (2013) was followed with minor modifications. Briefly, 0.1 mL of each overnight culture was inoculated into a 15-mL flask containing 10-mL precooled (4°C) Luria-Bertani (LB) medium (Oxoid, Cambridge, UK) at an initial population of ≈106 colony-forming units/mL. Bacteria were then stored at −20°C for 24 h. The viability of the samples was then determined by plate count. Before the viability test, the frozen samples were pre-incubated to dissolve for subsequent sampling at 37°C for 5 min. Similarly, 0.1 mL of each overnight culture was inoculated into 10 mL of phosphate-buffered saline (Shanghai Lingfeng Chemical Reagent Co. Ltd., China) and stored at 4°C for 72 h. Samples were taken to determine viability using the plate-count method. The Salmonella Enteritidis reference strain ATCC 13076 was used as a control strain. All the tests were done in triplicate. The data were analyzed by SAS Version 8.0 via one-way analysis of variance. Comparison of mean values was processed by Dunnett's test. Significance was defined when p<0.05.
PFGE assay
A total of 31 Salmonella Enteritidis isolates were genotyped by PFGE using the PulseNet protocol (Ribot et al., 2006). Briefly, 0.5 mL of each overnight culture was embedded in an agarose block, lysed, and the intact genomic DNA was digested by the restriction enzyme XbaI (Takara, Japan). These blocks were loaded into the wells of a 1% agarose gel (Lonza, ME). Electrophoresis was performed in 0.5×Tris-borate-EDTA buffer for 18 h in a CHEF DR-III (Bio-Rad, Hercules, CA) under pulsed times of 2.2 to 63.8 s. XbaI-digested Salmonella Braenderup strain H9812 DNA was used as reference DNA marker.
MLVA
Extraction of DNA, the polymerase chain reaction (PCR) system, and PCR cycling were conducted according to the method described by Lindstedt et al. (2004). PCR primers for seven variable-number of tandem repeats (VNTR) were previously described in the Laboratory Standard Operating Procedure for PulseNet MLVA (
Plasmid extraction and plasmid replicon typing
The Salmonella Enteritidis isolates were grown overnight in 5 mL LB medium (Oxoid) at 37°C with 200 rpm shaking. Plasmid DNAs were extracted by an alkaline lysis method (Feliciello and Chinali, 1993) and separated by electrophoresis on a 0.8% agarose gel with 1×Tris-acetate-EDTA buffer. In addition, plasmid replicon typing was examined by PCR assays with six singleplex primer pairs (Table 2) for six replicon types, including FIIA (FIIS), H1, L/M, ColE, IncU, and IncR.
PCR detection of virulence-associated genes
All the isolates were tested for the presence of virulence-associated genes using the primers listed in Table 2. The genomic DNAs of a total of 31 Salmonella Enteritidis isolates were extracted according to the method described by Liu et al. (2011). All the simplex PCR assays were carried out in 25-μL volumes: 2.5 μL of PCR buffer (10×, free Mg2+), 2.0 μL of MgCl2 (25 mM), 2.0 μL of dNTP (2.5 mM), 0.5 μL of each primer set (10 μM), 1.0 μL of Taq DNA polymerase (1 U/μL, Fermentas, Thermo Fisher Scientific, Lithuania), and 100 ng of template DNA. The PCR cycling was the same for all of the gene targets, consisting of an initial denaturation of 95°C for 3 min, 35 cycles of 94°C for 20 s, 58°C for 20 s, and 72°C for 30 s, with a final extension at 72°C for 5 min. Amplification products were analyzed by electrophoresis in 1.5% agarose gel, stained with ethidium bromide, and visualized under ultraviolet light.
Results
Most of the isolates (30/31) had the same PFGE pattern consisting of 13 bands, while 1 clinical isolate (SJTUF11262) was missing the approximately 55-kb band (Fig. 1). The PFGE fingerprint analysis showed that the isolates from most of the clinical (stool) samples (22/23) and all the mousse-cake samples were the same. Moreover, all of the isolates from this outbreak shared the same MLVA profile: 4-4-1-10-3-3-11. These highly similar PFGE patterns, together with the identical MLVA profile, suggest that this outbreak of gastroenteritis was caused by mousse cakes contaminated with Salmonella Enteritidis.
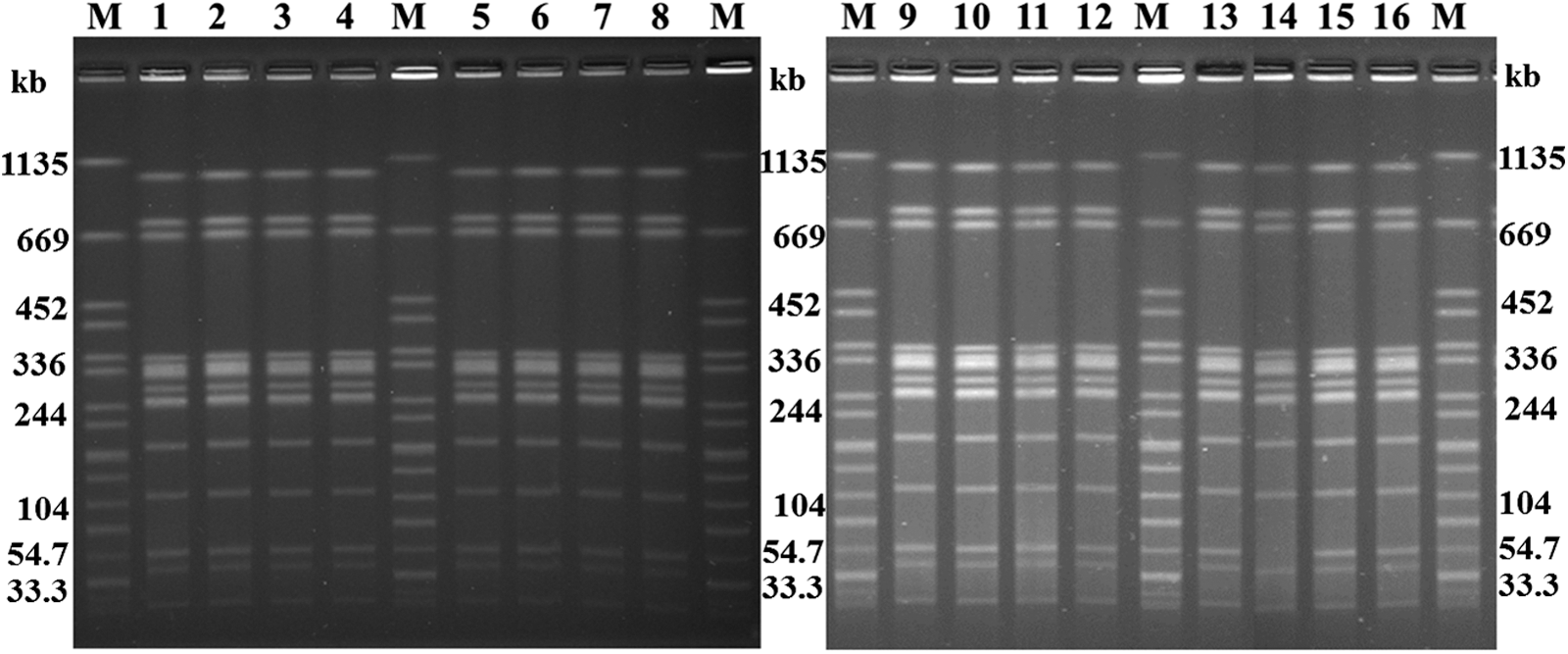
Pulsed-field gel electrophoresis fingerprints of selected Salmonella Enteritidis isolates. M, Salmonella Braenderup strain H9812; Lanes 1–8, Salmonella Enteritidis food isolates (SJTUF11248-SJTUF11253; SJTUF11265, SJTUF11266); Lanes 9–16, Salmonella Enteritidis clinical strains (SJTUF11241-SJTUF11244; SJTUF11261-SJTUF11263).
All stool isolates and food isolates had the same antibiotic susceptibility pattern. They were susceptible to all the drugs tested. Meanwhile, no differences were found in the growth rates in LB medium or LB supplemented with 20% sucrose (the typical sucrose concentration in mousse cakes) and motility of these isolates (data not shown). After 24-h freezing treatment, the survival rates of these Salmonella Enteritidis isolates were approximately between 10% and 30% (Table 1), meaning that the population of all these isolates was reduced by <1 log after freezing (–20°C). Moreover, the isolates displayed more resistance (p<0.05) to low temperature than the Salmonella Enteritidis reference strain ATCC 13076, which demonstrated an approximate 3.4% survival rate (Table 1). After refrigeration storage (4°C for 72 h, the general time period between manufacture and consumption of mousse cakes), the survival rates of these Salmonella Enteritidis isolates exhibited no significant (p>0.05) population reduction (data not shown).
Large plasmids were present in all of the Salmonella Enteritidis isolates except for the clinical isolate SJTUF11262 (selected strains are shown in Fig. 2). The results of replicon typing showed that all isolates carried a ColE plasmid (Table 1). All of the isolates also carried an IncFIIA incapability group plasmid except the clinical isolate SJTUF11262 (Fig. 3A and Table 1). Thus, the missing plasmid in clinical isolate SJTUF11262 probably belonged to the IncFIIA incapability group.
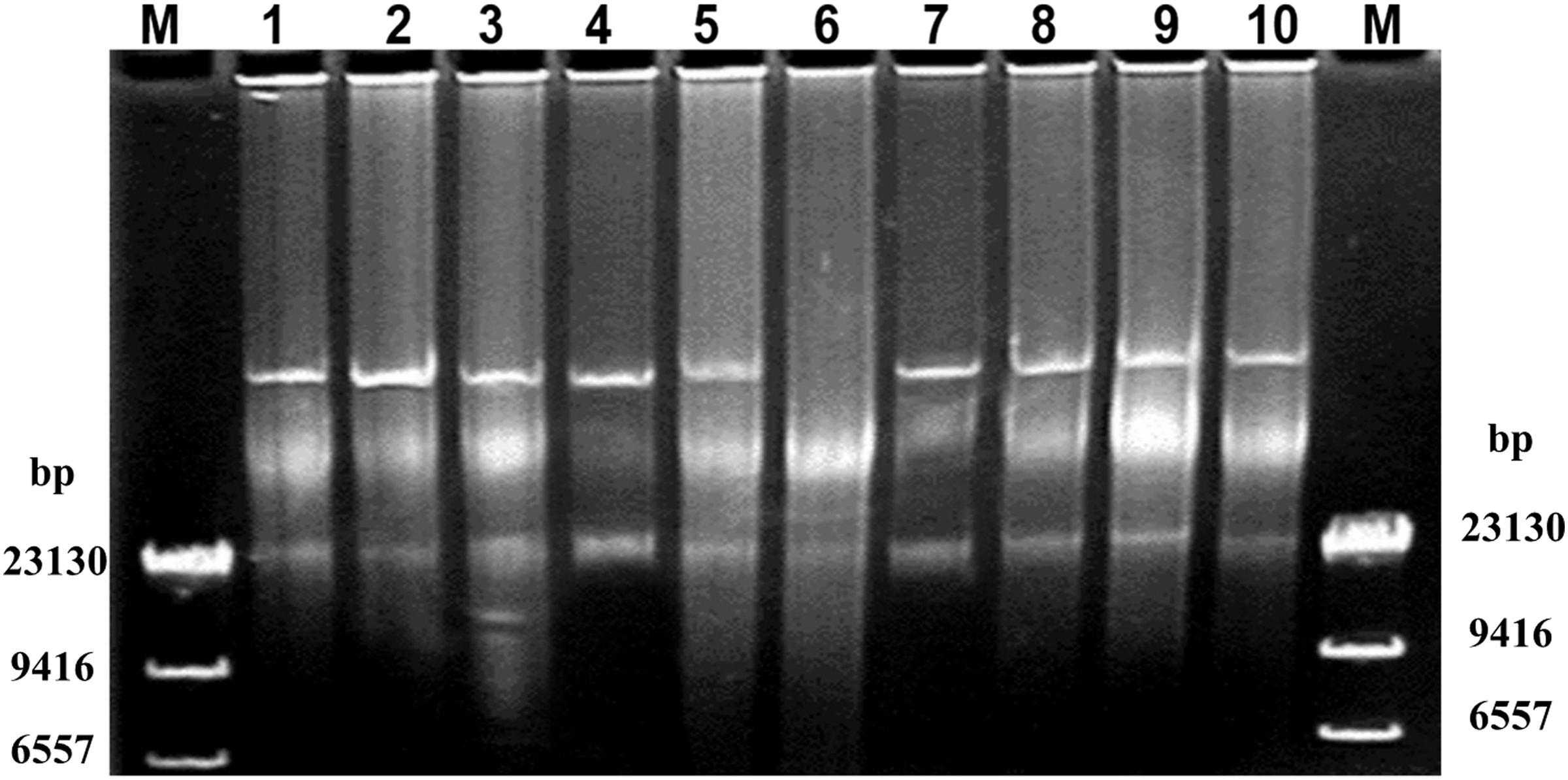
Plasmid analysis of selected Salmonella Enteritidis isolates. M, HindIII-digested λ DNA; Lanes 1–10, Salmonella Enteritidis strains SJTUF11241, SJTUF11243, SJTUF11247, SJTUF11255, SJTUF11258, SJTUF11262, SJTUF11263, SJTUF11264, SJTUF11266, SJTUF11268.
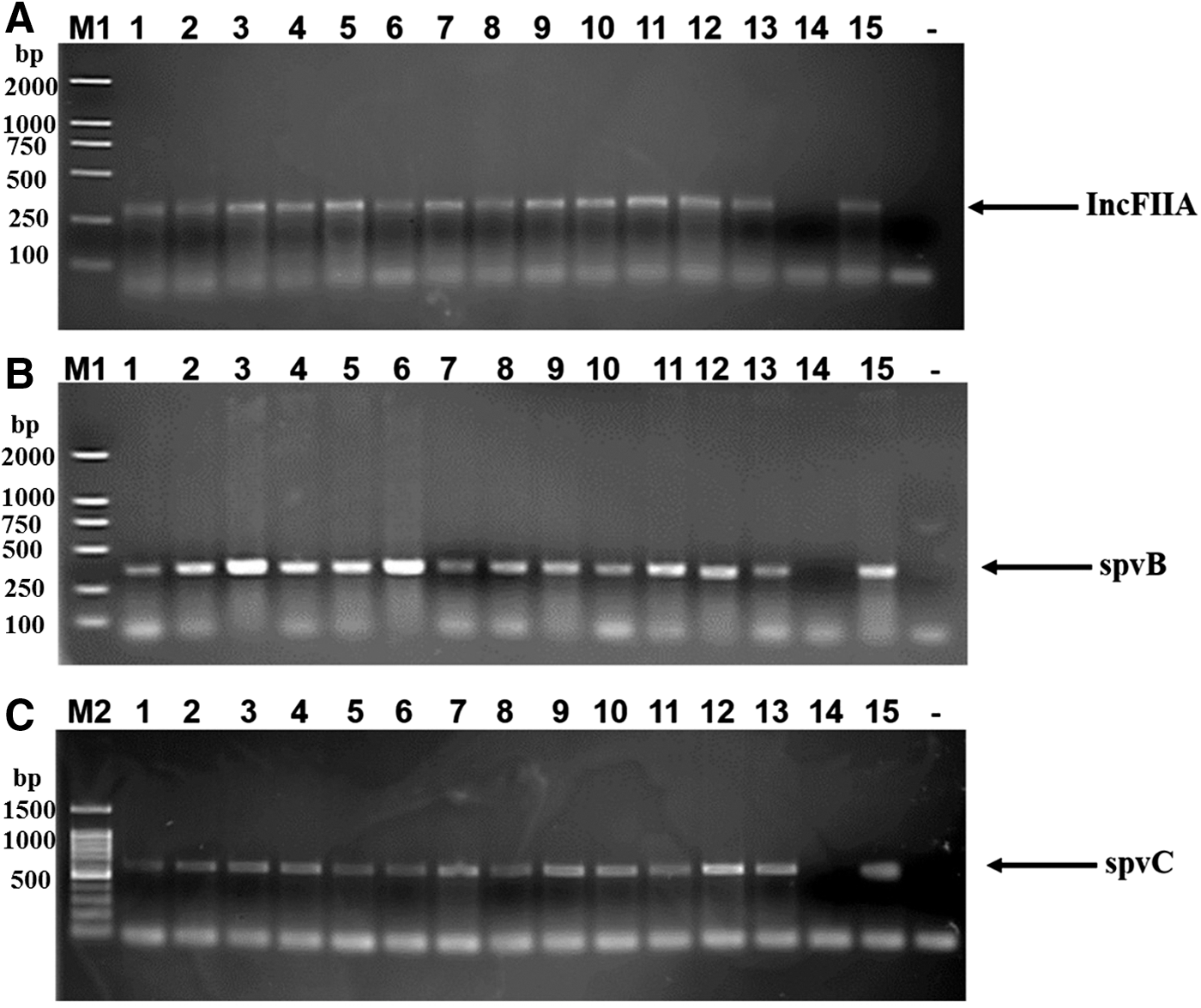
Polymerase chain reaction detection of
All isolates possessed invH (SPI-1), sopE (SPI-1), ttrB (SPI-2), rhuM (SPI-3), and spi4H (SPI-4) genes as revealed by PCR (Table 1). Most isolates (n=30, 96.8%) possessed the spvB and spvC genes, while clinical isolate SJTUF11262 lacked both the spvB and spvC genes (Table 1 and Fig. 3B and C).
Discussion
A number of factors may have contributed to the occurrence of this outbreak, including inappropriate manufacturing and storage processes, the use of contaminated ingredients, and cross-contamination by food handlers. A refrigeration or freezing treatment instead of baking is used during production and storage of mousse cake and its raw ingredients. Many reports show that raw ingredients such as eggs and poultry that are stored under refrigeration can be a serious food safety threat, because Salmonella outbreak isolates can survive very well under refrigeration and frozen storage (Pintar et al., 2007; Anderson et al., 2010; Pradhan et al., 2012). Also in the current study, the survival rates of these Salmonella Enteritidis isolates at the extremely low temperature treatment (freezing at −20°C) showed that they had greater cold tolerance than the reference strain ATCC 13076 (Table 1). After freezing, less than a 1-log decline of bacterial population was found in all of these isolates. Moreover, after storage at refrigerator temperatures (4°C) for 72 h, the bacterial population of these isolates remained almost unchanged (data not shown). Furthermore, short-term room temperature storage (at least 2 h) in the hands of the consumer was verified in this outbreak. These results suggest that mousse cake (a type of condensing dessert) manufacturers should make every effort to avoid introducing pathogenic bacteria such as Salmonella Enteritidis into their products because storage at room temperature by accident could give Salmonella Enteritidis an opportunity for logarithmic/secondary growth. In addition, it has been frequently reported that Salmonella Enteritidis infections are commonly associated with eggs (Patrick et al., 2004; Zhang et al., 2011). Eggs are indispensable ingredients for mousse cakes; moreover, raw eggs are generally used in cake making in China, and thus might have been the vehicle of infection for this outbreak. Unfortunately, implicated eggs were unavailable for further analysis because the processing site had been cleaned and the ingredients had been discarded. We cannot rule out the possibility of cross-contamination from one or more food handlers. Thus, mousse cake manufacturers should be educated to regard the risks of foodborne pathogenic bacteria during producing foods not subjected to thermal process. For example, pasteurized eggs should be used, raw products should be kept separate from the cooked ingredients, and all the food handlers should wash their hands after handling unprocessed ingredients.
Rapid methods for tracking of outbreak isolates are needed for food safety surveillance and clinical diagnosis. PCR-based methods have shown promise over the past few decades, which has had a revolutionary impact on the diagnosis and epidemiology of foodborne diseases. MLVA is a PCR-based subtyping method using seven multiplex primer panels for amplifying repeated sequences from seven different loci. An important advantage of MLVA used in outbreak studies is that the data can be obtained in a shorter amount of time (<24 h) than from methods such as PFGE. Moreover, the MLVA data can easily be shared among laboratories by a standardized allele string or MLVA type number. The MLVA type number acquired in this outbreak would be compared with that of other outbreaks to identify the prevalent MLVA type. In addition, PFGE (XbaI) analysis was performed and similar profiles were found for these isolates (Fig. 1). Although PFGE is currently the gold standard for subtyping of Salmonella Enteritidis, it requires skilled personnel, and it usually takes longer than 24 h to complete. Since MLVA is easier and faster than PFGE (Kurosawa et al., 2012), MLVA may be used as the preferable subtyping method. However, differences in plasmids cannot be displayed by MLVA, suggesting that a combination of two or more genotyping methods that rely on different parameters may be required to better distinguish among isolates in an outbreak.
In this study, all isolates harbored the SPI-related genes (Table 1), which are important virulence factors that help the pathogen in adhesion and invasion mechanisms. For example, SOP proteins (SOP A–E) serve as effector proteins (Wallis and Galyov, 2000; van Asten and van Dijk, 2005) that are involved in the pathogenesis of salmonellosis. In addition, the inv A–H gene products function during invasion of the intestinal mucosa (Fluit, 2005; Chuanchuen et al., 2010) and are important systemic infection-related factors of Salmonella. Moreover, some virulence genes are located on large virulence plasmids, such as pSTV that encodes the spv operon involved in bacterial replication in extraintestinal sites (Gebreyes et al., 2009; Hur et al., 2011). In addition, the spvB/C genes were observed in most of the isolates (Fig. 3 and Table 1). Taken together, the presence of these virulence or virulence-related genes suggests that these isolates could give rise to public health problems if they were dispersed in the general human population.
Bichler et al. (1994) demonstrated that the spvB/C genes are associated with a plasmid of 54–57 kb in Salmonella Enteritidis. In our study, most of the isolates carried an Inc/FIIA plasmid of this size. These results correlated with a previous study by Rychlik et al. (2006). It is puzzling that one clinical isolate (SJTUF 11262) did not carry this common 55-kb Inc/FIIA plasmid. Whether or not the loss of this plasmid (or spvB/C genes) could influence the infection or/and pathogenicity of this isolate needs further investigation.
Conclusions
Antimicrobial susceptibility testing and two genotyping schemes (MLVA and PFGE) were employed to examine the relatedness of the outbreak-associated clinical and food isolates. Phenotypic analysis data and subtyping results showed that this Salmonella Enteritidis outbreak in Ningbo (China) was caused by Salmonella Enteritidis–contaminated mousse cake. Furthermore, the possession of virulence-associated genes in these outbreak strains indicated that these isolates could cause infection in humans. This study provided further insights into the use and handling of raw ingredients during the manufacture of foods without thermal processing, as Salmonella Enteritidis can survive very well under refrigerated and frozen treatment and improper storage could support its survival and growth, which may be a serious food safety threat.
Footnotes
Acknowledgments
This research was supported by the National Natural Science Foundation of China (31230058) and the Ministry of Science and Technology of China (2012AA101601, 2012BAK17B10, and 2012BAD29B02).
Disclosure Statement
No competing financial interests exist.