
Abstract
Myxobolus honghuensis parasitizes the pharynx of the allogynogenetic gibel carp Carassius auratus gibelio (Bloch), an economically important food fish with large consumption rates in China. The fact that it is hard to detect the cysts formed by M. honghuensis without euthanizing the fish and the spores can reach the consumers has prompted us to investigate the effect of M. honghuensis on conventional, immunocompetent BALB/c suckling mice, in order to consider the possible pathological effects in general immunocompetent consumers by the ingestion of Myxobolus-infected fish. BALB/c suckling mice were inoculated with serial dilutions of the spore suspension and then hematological and histological studies were performed. Tests of suckling mice showed no statistically difference of fluid accumulation ratio between mice inoculated with 1.65×106 M. honghuensis spores and negative control after 8-h incubation. Spores did not disrupt intestinal histology, and no abnormal bowel movements were observed within 20 h postinoculation. No anomalous hematology parameters were documented. These results, taken together, reveal that M. honghuensis spores do not cause pathology (diarrhea and elevated fluid accumulation) in BALB/c suckling mice. However, an extrapolation is not currently feasible because of lack of a highly quantifiable risk extrapolation model, inaccessibility to data for the effect of M. honghuensis on humans, and lack of consideration of the immunosuppressed population. Our study extends the results of the effect of myxozoan on suckling mice and serves as baseline for future experiment using the same model in evaluating the myxozoan-caused foodborne disease.
Introduction
I
Myxobolus honghuensis parasitizes the pharynx of the allogynogenetic gibel carp C. auratus gibelio (Liu et al., 2012). Cysts, formed from M. honghuensis in the pharynx, are not easily detected without euthanizing the fish. Thus, allogynogenetic gibel carp infected with M. honghuensis spores may be marketed to consumers and be ingested without any prior warning.
Myxozoan spores had been observed in gastrointestinal patients' feces over the past decade (McClelland et al., 1997; Boreham et al., 1998), and a few cases involving immunosuppressed patients (Agudelo et al., 2000; Moncada et al., 2001) resulted in unproven guessing about the opportunistic nature of Myxobolus spp., the most species-rich genus in Myxosporea (Lom and Dyková, 2006). However, in all cases, the pathogenic role of myxozoan spores remained ambiguous. Therefore, several efforts were made to establish animal models of ingestion of spores. For instance, Kudoa spp. were found to be capable of provoking humoral immune responses in BALB/c mice through a subcutaneous route (Martínez de Velasco et al., 2002) or by oral administration (Martínez De Velasco and Cuéllar, 2003). Another BALB/c mice model established for M. cerebralis, the causative agent for salmonid whirling disease, focused on evaluating viability instead and did not show data for pathogenicity on mice (El-Matbouli et al., 2005). Recently, Kudoa septempunctata was found to cause more than 100 cases of foodborne disease in Japan (Ohnishi et al., 2013), and the pathogenicity role was successfully validated by inducing similar clinical symptoms (e.g., severe diarrhea, elevated fluid accumulation) in ddY suckling mice (Kawai et al., 2012). Therefore, K. septempunctata became the first reported human pathogenic myxozoan.
After the controversial findings of the possible pathogenic role of Myxobolus spp. both in immunocompetent and immunosuppressed individuals (Agudelo et al., 2000; Moncada et al., 2001) and the solid evidence that certain species do cause foodborne disease, possible ingestion of myxozoan spores through cooked or raw fish has become an important concern.
Consequently, we have developed this work to determine whether M. honghuensis shows pathogenicity for laboratory animals, in order to consider the possible pathological effects in general immunocompetent consumers by the ingestion of M. honghuensis–infected fish. Hence, the conventional, immunocompetent BALB/c suckling mice model was chosen.
Materials and Methods
Animals
Four-to-5-day-old specified-pathogen-free (SPF) BALB/c mice (mean weight, 3.65±0.80 g) were obtained from the Center for Animal Experiment of Wuhan University (Wuhan, China). Specimens (125, 25, and 27) were used to study the kinetics of fluid accumulation ratio (FA ratio), bowel movements, and hematology, respectively. All experiments were conducted in an animal biosafety level II lab at SPF levels (Institute of Virology, Chinese Academy of Sciences). The animal care protocol complied with the guidelines established by the National Institutes of Health and the International Society for Development Psychobiology (NIH, 1985).
Preparation of spore inocula
Allogynogenetic gibel carp were harvested from Honghu Lake, Hubei Province, China in June 2011. These fish were examined for myxosporean infection and when mature plasmodia were found, the samples were stored at −20°C until spore isolation.
The pharynxes of infected fish were homogenized by a manual glass tissue grinder and suspended in 0.1 M phosphate-buffered saline (PBS), pH 7.2, and then filtered through cotton gauze to remove aggregated tissues. The filtrate containing M. honghuensis spores was purified by sucrose gradient centrifugation.
The density of the spore suspension was determined using a hemocytometer. The final spore concentration reached 1.65×107 spores/mL. Serial dilutions (10- and 100-fold) of the spore suspension were made in PBS to perform the dose–response study. A portion of the spore suspension was heated at 90°C for 10 min (FDA, 2001) to investigate their thermostability.
Inoculation of suckling mice
Suckling mice (4–5 days old) were separated from their mother 2 h prior to inoculation. All spore suspensions were mixed with 0.01% (wt/vol) Evans blue dye before inoculation (dissolved in PBS, high-pressure steam sterilized, and then aseptically added). All mice were inoculated with 0.1 mL of spore suspensions directly into the milk-filled stomach of each animal using a 1-mL syringe equipped with 21-gauge stainless steel intramedic tubing (Bolige Industry and Trade Co., Ltd., Shanghai, China); inner diameter, 0.020 inch (≈0.51 mm); outer diameter, 0.032 inch (≈0.82 mm). Then the suckling mice were incubated at 25°C in absence of their mother until dissection. The final inoculation concentration was the following: high-dose group (1.65×106 spores/mice), middle-dose group (1.65×105 spores/mice), and low-dose group (1.65×104 spores/mice). Suckling mice without oral challenge were incubated in the same condition as the exposed group and served as a negative control. Suckling mice inoculated with equal amounts of PBS and 0.01% Evans blue served as a solvent control.
Determination and kinetics of FA ratio
Five suckling mice were sacrificed by cervical dislocation in each group at predetermined time intervals. Secretory diarrhea often reflects massive secretion of fluid and electrolytes by the small intestine (Izzo et al., 2003). The FA ratio, a parameter reflecting the degree of intestinal fluid accumulation, was measured as previously described (Bäumler et al., 1996; Ohnishi et al., 2013). Briefly, the intestinal tract, from the pylorus to the end of the rectum after the open management of the abdominal cavity, of each mouse was removed. The conglutinate viscera and mesentery were removed prior to weighing the intestinal tract. Test animals with absence of blue dye in their intestinal tracts were discarded.
FA ratio was expressed as the following: weight of intestines/remaining carcass weight. The FA ratio in each group was determined at desired time intervals: 0, 2, 4, 6, and 8 h postinoculation for negative control (n=25); and 2, 4, 6, and 8 h postinoculation for the high-dose group (n=20), middle-dose group (n=20), low-dose group (n=20), heat-treated group (n=20), and solvent control (n=20).
Bowel movements
The effect of M. honghuensis spores on bowel movements was evaluated using groups of eight or nine, 4–5-day-old BALB/c mice. Each mouse received a 0.1-mL oral administration of high-dose spore suspension or PBS and 0.01% Evans blue. Mice from the same group were then kept in 1 cage fitted with filter paper over the cage floor for 20 h at 25°C, after which an examination was conducted. Bowel movements were evaluated by calculating the ratio between the number of observed feces and the number of tested mice. The stool form on the filter paper was also recorded. Mice without inoculation treatment were used as negative control.
Feces collected during the bowel movement and FA ratio kinetics studies were checked by wet smear to observe the presence of spores. We examined the time course of appearance of spores in feces and the integrity of spores after passing through the alimentary tract of the mice.
Hematology
At 6 h postinoculation, nine suckling mice from each group—negative control (group 1), solvent control (group 2), and high-dose group (group 3)—were anesthetized with ether. Blood samples were collected via the carotid artery in tubes containing an equal amount of dipotassium ethylenediaminetetraacetic acid (EDTA-2K) and were immediately sent for hematology analysis. Red blood cell (RBC) count, white blood cell (WBC) count, hemoglobin (Hb), hematocrit (Hct), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), mean corpuscular volume (MCV), platelets, monocytes, basophils, and eosinophils counts were performed using a hematology auto-analyzer (XS-1000i, Sysmex, Kobe, Japan).
Intestinal histopathology
Mice were killed after recording of bowel movements and measurement of FA ratios. The intestines were collected and then fixed in PBS-formalin (10%, pH=7.2) for 24–48 h, before being routinely processed to create 4-μm paraffin-embedded sections stained with hematoxylin and eosin for histological analysis.
Statistical analysis
The statistical differences of FA ratios and bowel movement numbers between negative control and exposed groups were evaluated using Student t-test or Mann–Whitney U test in combination with the Levene test to evaluate the homogeneity of variances. All results were expressed as means±SD and probability level of p<0.05 was considered significant. Statistical analysis was accomplished by Statistica 6.0 (2001; Statsoft, Tulsa, OK) and Graphpad Prism 5 software package (GraphPad Software, Inc., San Diego, CA).
Results
Kinetics of FA ratio
All the inoculated and controlled animals survived throughout the 8-h experimental period. In the negative control group, FA ratios remained stable during the experimental period as shown in Figure 1. Meanwhile, Figure 1 also indicates that no statistically significant differences (p<0.05) in FA ratios were found between the experimental groups and the negative control group throughout the incubation period.
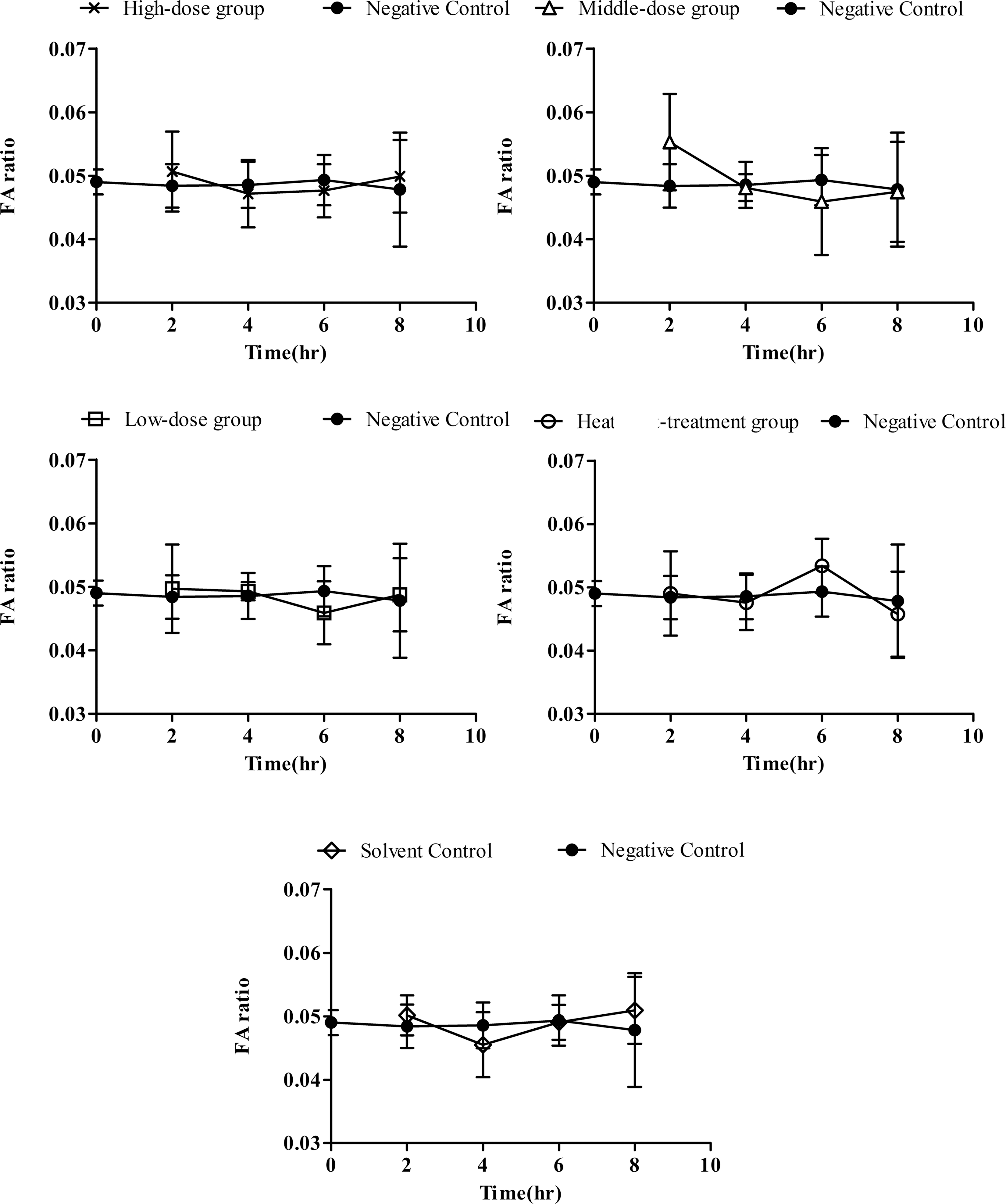
Kinetics of fluid accumulation (FA) ratio in high-dose group (1.65×106 spores/mice), middle-dose group (1.65×105 spores/mice), low-dose group (1.65×104 spores/mice), heat-treatment group (1.65×106 spores/mice), solvent control (phosphate-buffered saline and 0.01% Evans blue) and negative control. Means of FA ratios±SD for five mice are shown. No significant differences (p<0.05) were found at any time interval at any group.
Bowel movements
There was no difference (p<0.05) in FA ratio between the high-dose group and the negative control group. Also, there were no significant differences between the number of feces collected from the three experimental groups. All groups had a similar ratio of the number of observed feces to the number of tested mice as follows: high-dose group, 0.75 (6/8), solvent control, 0.78 (7/9), and negative control, 0.75 (6/8) (Table 1). There were no mortalities in the solvent control and negative control groups, though 2 mice from the high-dose group died after 20-h incubation. The necropsy showed serious stomach hemorrhage due to accidental injury during inoculation in one dead mouse. In the other one, the absence of Evans blue dye in the gastrointestinal tract (stomach and intestines) indicated a failed inoculation. Considering these results, the two dead mice were excluded from statistical analysis. Like the FA ratio, no abnormal bowel movements (e.g., watery diarrhea, mucoid, or pasty discharges) were observed (Fig. 2).
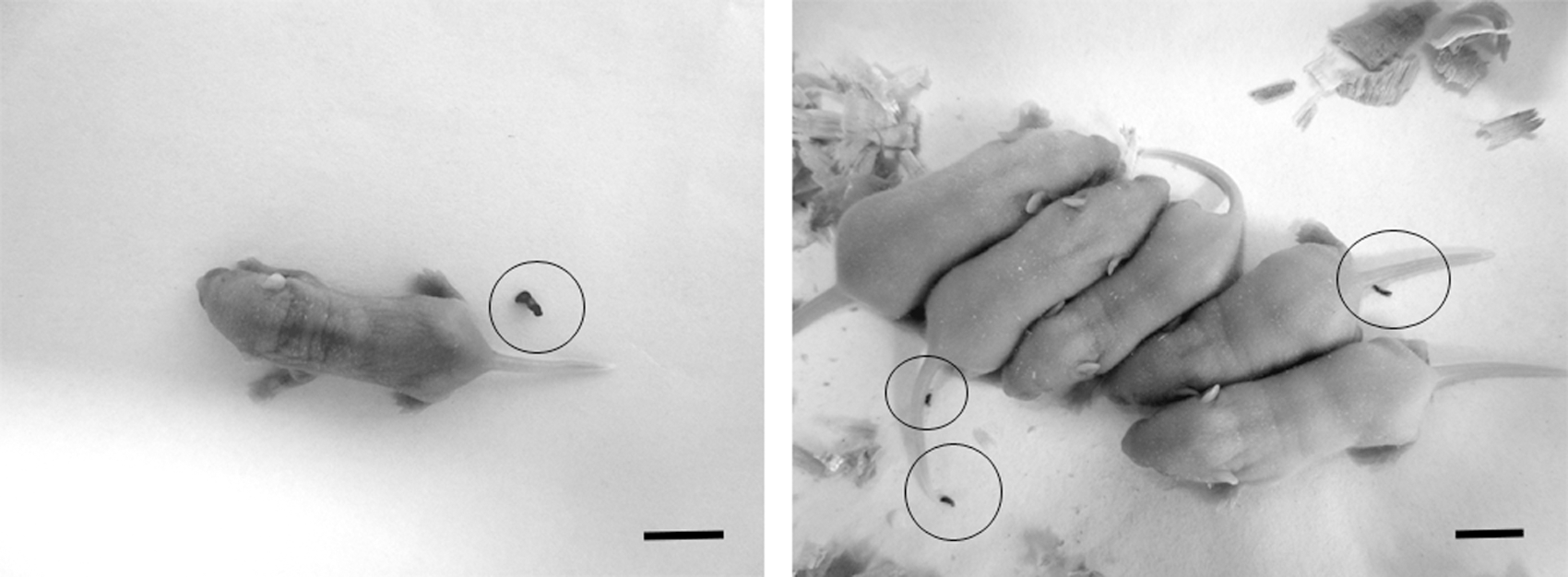
Suckling mice from high-dose (1.65×106 spores/mice) group (left panel) and negative control (right panel) both excreted normal feces after 20-h incubation at 25°C. Each circle in the panel represents a feces. Scale bars=10 mm.
Abnormal stool form was defined as observation of abnormal stool form (loose/watery stools, lumpy/hard stools, or defecation strain).
Phosphate-buffered saline and 0.01% Evans blue.
FA ratio, fluid accumulation ratio, calculated by all the tested animals in each group.
Feces from 2, 4, 6, 8, and 20 h postinoculation were collected and checked by wet smear. M. honghuensis spores were found in feces from all challenge groups except control group feces. Almost all the spores found in the feces were intact, and no more than 5% of spores extruded the polar filaments (Fig. 3). Fecal spores counted after 4 h were observed to be less, which increased up to a peak number after 8 h and gradually decreased after 20 h postinoculation.

Light micrographs of mature spores of Myxobolus honghuensis in orally challenged BALB/c suckling mice unstained fecal wet smear. Feces were collected 8 h postchallenge.
Hematology
All hematological parameters from tested animals sacrificed at 6 h postinoculation are shown in Table 2. In group 3 (high-dose group), most of the indexes (e.g., RBC, WBC, Hb, Hct, MCH, MCHC, MCV) were not statistically different from group 1. The spore inoculation treatment had no effect on circulating platelets, basophils, or eosinophils numbers. The monocytes level was lower in group 3, while the basophils and eosinophils were at normal level. In group 2, all the indexes were similar to group 1.
p<0.05, significantly different from negative control.
Red blood cell count (RBC), white blood cell count (WBC), hemoglobin (Hb), hematocrit (Hct), corpuscular hemoglobin (MCH), corpuscular hemoglobin concentration (MCHC), and corpuscular volume (MCV) are abbreviated parameters recorded in the table.
Intestinal histopathology
In Figure 4, light micrographs of hematoxylin and eosin–stained intestinal tissue sections from negative control, solvent control, and high-dose group (4 h and 20 h postinoculation) are shown. No obvious differences in brush border, crypt cell, epithelium, mucosa, or submucosa were observed. The intestine villi morphology remained characteristic and distinct. Normal intraepithelial lymphocytes were found in lamina propria, and there was no evidence of inflammation (Fig. 4A–C).
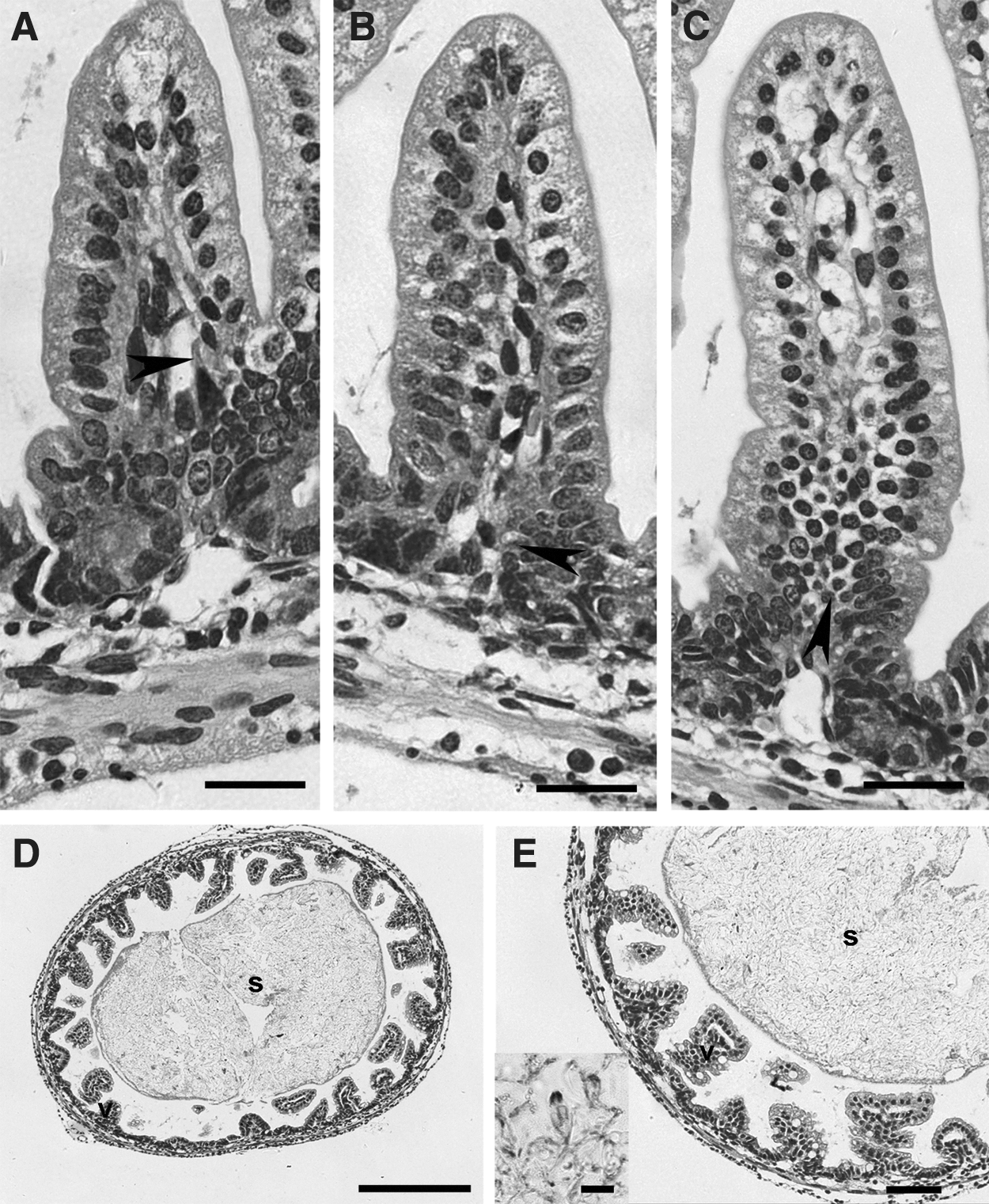
Histopathology of hematoxylin and eosin–stained 4-μm-thick intestinal tissues. (A–C) Scale bars=20 μm; (D) scale bars=250 μm; (E) scale bars=10 μm (inset), 50 μm (outer panel).
Chyme-like formations, containing numerous M. honghuensis spores, were observed within the intestine in samples fixed 4 h postinoculation (Fig. 4D and E). Intact spores with two polar capsules could be found inside the intestine. There was no observable tissue attachment, reaction, or degeneration of the digestive tract where the parasites were located (Fig. 4E). No spores were observed in the intestine samples fixed at 20 h postinoculation.
Discussion
To date, only a limited number of studies have evaluated possible human pathogenicity and/or allergenicity of myxozoans (Martínez de Velasco et al., 2002, 2008; Martínez De Velasco and Cuéllar, 2003; Ohnishi et al., 2013). However, myxozoan-related disease has received attention because of the recent outbreak of food poisoning in Japan related to K. septempunctata (Iwashita et al., 2013). This parasite can also cause diarrhea and elevated FA ratios in 4–5-day-old ddY suckling mice, which can be inhibited through heat treatment (Ohnishi et al., 2013). Conversely, we did not observe intestinal fluid elevation in suckling mice after exposure to M. honghuensis spores. Furthermore, no anomalous FA ratios were observed in mice that received heat treatment at 90°C for 10 min, which could be explained by the absence of initial pathogenicity mentioned above. Mechanisms of K. septempunctata pathogenesis have recently been demonstrated to release sporoplasms, and to invade and damage intestinal tissue (Kawai et al., 2012). In our study, no tissue attachment, reaction and degeneration, or sporoplasm release were observed. Among the chyme-like formations found in the intestines, it was difficult to distinguish whether the polar filaments of intact spores were extruded or not. We demonstrated the extruding ability of polar filaments by our viability procedure; however, since most spores found in the feces did not extrude polar filaments, it was thought that M. honghuensis spores kept polar filaments detruded until defecation. These results were consistent with previous findings that spores can tolerate passage through the vertebrate gut without loss of infectivity to its natural oligochaete host (El-Matbouli and Hoffmann, 1991). Furthermore, attachment and binucleated sporoplasm penetration of the gut epithelial cells were necessary processes for natural host infection, and spores would lose their infectivity if polar filaments were prematurely extruded (Azevedo et al., 2011). Differing polar filament behavior between K. septempunctata and M. honghuensis during epithelial invasion could be the underlying mechanism driving K. septempunctata pathogenicity. We speculated that K. septempunctata spores, when incidentally swallowed by human or mouse instead of its assumed natural annelid worm host, could prompt the sporoplasm-penetrating process and cause damage to intestinal cells, whereas the same process would not be triggered by M. honghuensis spores. The normally shaped stools and stable number of feces observed in the high-dose groups were similar to the negative control. This differed from the abnormal bowel movements in mice inoculated with a K. septempunctata spores suspension described previously (Kawai et al., 2012), and intuitively supports the viewpoint that M. honghuensis spores are not pathogenic for BALB/c suckling mice.
Despite the 20-h maximum incubation time, the majority of spores present in the mice feces were intact. The results support the common view that myxozoan spores can remain intact after passing through the vertebrate gut (El-Matbouli et al., 1992; McClelland et al., 1997; Boreham et al., 1998). Our histopathology results showed that no clusters of spores were present in intestine 20 h postinoculation. Therefore, the number fluctuation of spores in feces could be explained by regular defecation patterns: Most M. honghuensis spores would be excreted 8 h after inoculation, and few of them would reside in the intestinal tract 20 h after inoculation, thus excluding the possibility of parasite residing and chronic disease development.
The nonpathogenicity of M. honghuensis for suckling mice was further confirmed by hematology parameters, which showed no abnormalities except for the decreased monocytes level in the high-dose group. We prefer to consider it an error instead of leukopenia because the high-dose group (group 3) showed relatively higher coefficient of variation (SD/mean, CV%) compared with the negative control (group 1) and solvent control (group 2), which means data with higher variability and less stability.
It is unlikely that mice strains used here would cause false-negative results because many mice strains, including ddY, BALB/c, Swiss-Webster, CF1, ICR, and CFW, have been fittingly applied for the suckling mice model previously (Nishibuchi et al., 1983; Kothary and Richardson, 1987; Audicana et al., 2002; Izzo et al., 2003; Ohnishi et al., 2013). Furthermore, the relatively stable fluid accumulation ratio in our negative group supports the utility of this BALB/c suckling mice test in parasitology research.
Conclusions
Our present data suggest that M. honghuensis spores do not induce diarrhea and elevate fluid accumulation in BALB/c suckling mice. These results, combined with the absence of clinical symptoms and observation of normal hematology and histology, demonstrate that M. honghuensis is not pathogenic for BALB/c suckling mice. However, it is too early to say that the present insights can be applied to humans with current data in hand for the following reasons: (1) lack of a highly quantifiable risk extrapolation model that outlines the steps leading from estimate of myxozoan pathogenicity in mice to that in human population; (2) inability to access data for the effect of M. honghuensis on humans; and (3) in most risk assessments, the human population is considered as a whole and little or no efforts have been devoted to protect the rare individual, namely, the immunosuppressed population in this case. The former two limitations could be partly solved by establishing a stringent mathematical model that will account for all of the elements required to risk extrapolation based on this suckling mice model with the availability of clinical information from human pathogenic myxozoans K. septempunctata. On the other hand, the possible pathogenic role of Myxobolus spp. in immunosuppressed patients must be elucidated and a corresponding humanized immunodeficient mouse model will provide promising solutions for this in future work. Taken together, this study not only demonstrates nonpathogenicity of M. honghuensis in BALB/c suckling mice, but also serves as a baseline for future experiments using the same model in evaluating the newly emerged myxozoan-caused foodborne disease that deserves attention.
Footnotes
Acknowledgments
We thank Sean Roon (Oregon State University, U.S.A.) for his valuable suggestions and critical reading of the manuscript. This work was financially supported by New Century Excellent Talents in University (NCET-12-0866) and the Fundamental Research Funds for the Central Universities (2012PY004, 2013PY023).
Disclosure Statement
No competing financial interests exist.