
Abstract
Objective:
The objective of this study was to investigate the seroepidemiology of immunoglobulin G (IgG) antibodies against Norovirus (NoV) GII.3 and GII.4 genotypes among children younger than 5 years with acute diarrhea in Xi'an, China.
Materials and Methods:
A total of 362 serum samples were collected from diarrheal children in the Department of Digestive Diseases of Xi'an Children's Hospital between March 2009 and October 2012. Recombinant capsid proteins of NoV genotypes GII.3 and GII.4 were expressed using the baculovirus expression system. The viruslike particles (VLPs) were examined by electron microscopy and Western blot, and VLPs were used as antigens for serological IgG tests using enzyme-linked immunosorbent assays.
Results:
The overall seroprevalence for GII.4 (86.2%) was significantly higher (p<0.01) than for GII.3 (67.9%). The seroprevalence remained in a high and stable level (70.9% for GII.3 and 88.7% for GII.4) in children under 1 year of age, then dropped in the age group 12–17 months (49.3% for GII.3 and 68.1% for GII.4), and finally increased to 77.8% for GII.3 and 96.8% for GII.4 in the group >18 months. The seroprevalence in the age group 12–17 months showed more statistically significant differences than the other age groups for both GII.3 and GII.4 (p<0.05).
Conclusions:
In conclusion, seroprevalence of NoV GII.3 and GII.4 was high in young children in Xi'an, China, and the anti-GII.4-positive rates were statistically higher than GII.3 across all the age groups.
Introduction
N
NoV genome contains three open reading frames (ORFs). ORF1 codes a single polyprotein that is the precursor to the nonstructural proteins; ORF2 and ORF3 encode the single capsid protein (VP1) and the minor structural protein (VP2), respectively (Mauroy et al., 2009). NoVs are divided into six genogroups (I–VI), and each group is further classified into genotypes based on the phylogenetic analysis of polymerase or capsid region (Hasing et al., 2014), but only GI, GII, and GIV genogroups can infect humans (Li et al., 2010). In particular, GII is the most important genogroup for sporadic NoV infections in young children and outbreaks in adults. GII contains at least 17 genotypes based on pairwise distance analysis (Zheng et al., 2006). In the past decade, NoV GII.4 and GII.3 have been identified as the predominant genotypes in young children with nonbacterial acute diarrhea in China (Dai et al., 2004; Cheng et al., 2010; Hoa Tran et al., 2013). Our studies showed that GII.3 and GII.4 were the only NoV genotypes in hospitalized diarrheal children under 5 years in Xi'an, China (Zhang et al., 2011), and GII.3 and GII.4 infections in young children are significantly associated with host genetic susceptibility (Liu et al., 2014). The objective of this study was to investigate epidemiological information about NoV seroprevalence in diarrheal children in Xi'an, China through GII.3 and GII.4 VLPs expressed by the baculovirus expression system.
Materials and Methods
Study population and sample collection
Serum and fecal specimens (used in previous studies [Zhang et al., 2011; Liu et al., 2014]) were collected from hospitalized children ≤5 years of age who were clinically diagnosed with acute gastroenteritis (defined as ≥3 loose or watery stools per day without blood and mucus) and whose clinical symptoms lasted <2 weeks before admission to the Department of Digestive Diseases of Xi'an Children's Hospital between March 2009 and October 2012. Serum samples were collected once within 48 h of admission in the acute course of diarrhea. Before participation in our study, parents or guardians were asked to sign an informed consent.
NoV genotyping and GII.3 and GII.4 VLPs development
A 20% stool suspension was prepared in RNAse- and DNAse-free water, and 500 μL of the suspension was mixed with an equal volume of Vertrel (Miller-Stephenson, Danbury, CT). After incubation at 4°C overnight, the samples were centrifuged at 12,000×g for 15 min at 4°C. Subsequently, a 140-μL supernatant was removed and used for Norovirus RNA extraction using the QIAamp Viral RNA Mini Kit (Qiagen, Valencia, CA). Detailed methods for NoV RNA extraction, polymerase chain reaction (PCR) amplification, and genotyping has been described in our previous studies (Zhang et al., 2011; Liu et al., 2014). Based on previous results, a 2.3-kb cDNA fragment, which contains the VP1 region (NoV capsid protein) and the VP2 region encoding structural proteins of NoV GII.3 and NoV GII.4, was amplified by the primers shown in Table 1. The amplified product-cDNA fragment was cloned into a PCR®4-TOPO vector (Invitrogen, Carlsbad, CA), and the recombinant PCR®4-TOPO was transferred into a baculovirus pFastBac1 donor plasmid (Invitrogen, Carlsbad, CA). Subsequently, the recombinant bacmids were transfected into Sf9 cells (Invitrogen, Grand Island, NY) at a multiplicity infection of 1.0 using the Bac-to-Bac®Baculovirus expression system (Invitrogen, Carlsbad, CA). After a 4-day incubation of the infected Sf9 cells, VLPs secreted into the media were separated from the supernatant by centrifugation at 9790×g for 30 min at 4°C. Then the supernatant was concentrated by a vivaflow 200 ultrafiltration system (Sartorius, Gottingen, Germany) at 4°C.
The concentrated product contained VLPs that were applied to 10–50% discontinuous sucrose gradient centrifugation at 115,255×g for 1 h. The fractionated VLPs were resuspended in phosphate-buffered saline (PBS) and then pelleted by ultracentrifugation at 115,255×g for 1 h. The pellet was resuspended in PBS with proteinase-inhibitor cocktail (Pierce, Rockford, IL).The VLPs were examined by electron microscopy (EM) and Western blot and used as antigens for immunological test in this study.
Western blot
The purified GII.3 and GII.4 VLPs were transferred to a nitrocellulose membrane after being electrophoresed on a sodium dodecyl sulfate polyacrylamide gel. The membrane containing target proteins was blocked with skim milk at room temperature for 2 h. After washes, the membrane was incubated in PBS with 1:1000 diluted monoclonal anti-NoV antibodies (Kim Laboratories Inc., Rantoul, IL) overnight. On the next day, the membrane was incubated with secondary antibody (Kirkegaard & Perry Laboratories, Inc., Gaithersburg, MD) for 1 h. After washing three times with PBS, the membrane was exposed to chemiluminescent peroxidase substrate and finally displayed in a dark condition.
Detection of GII.3 and GII.4–specific IgG in sera by enzyme-linked immunosorbent assay (ELISA)
A direct ELISA was used to detect specific IgG of NoV GII.3 and NoV GII.4 in sera. The GII.3 and GII.4 VLPs examined by both EM and Western blot were used as antigens for ELISA. Polystyrene plates were coated with NoV GII.3 or NoV GII.4 VLPs (2 μg/mL) in 5% Blotto in PBS-Tween and the plates were incubated overnight at 4°C, washed 3 times, and then incubated with duplicate serum samples diluted 1:400 in blocking solution for 1 h at 37°C. After 3 washes, alkaline phosphatase–labeled rabbit anti-human IgG (Sigma-Aldrich Co., St. Louis, MO) diluted 1:2500 were applied to plates. After incubation for 30 min at 37°C, plates were washed and then 100 μL p-nitrophenyl phosphate solution (Sigma-Aldrich Co.) was added to each well. After incubation in dark conditions for 10–30 min at room temperature, the optical densities (OD) at 450 nm were measured by an ELx800 plate reader (BioTek Instruments, Inc., Winooski, VT). OD values greater than the cut-off value (add 0.1 to the OD value of the negative control) were defined as positive.
Statistical analysis
Chi-square test was used to compare NoV IgG-positive rates in different age groups by SPSS 18.0 (SPSS Inc., Chicago, IL). A significance level of 0.05 was set for the analysis.
Results
VLPs of NoV GII.3 and NoV GII.4
The GII.3 and GII.4 VLPs were purified by sucrose gradient centrifugation and were confirmed by Western blot (Fig. 1A and B). Purified VLPs were also observed under EM showing predominant 38-nm-diameter VLPs (Fig. 2A and B).
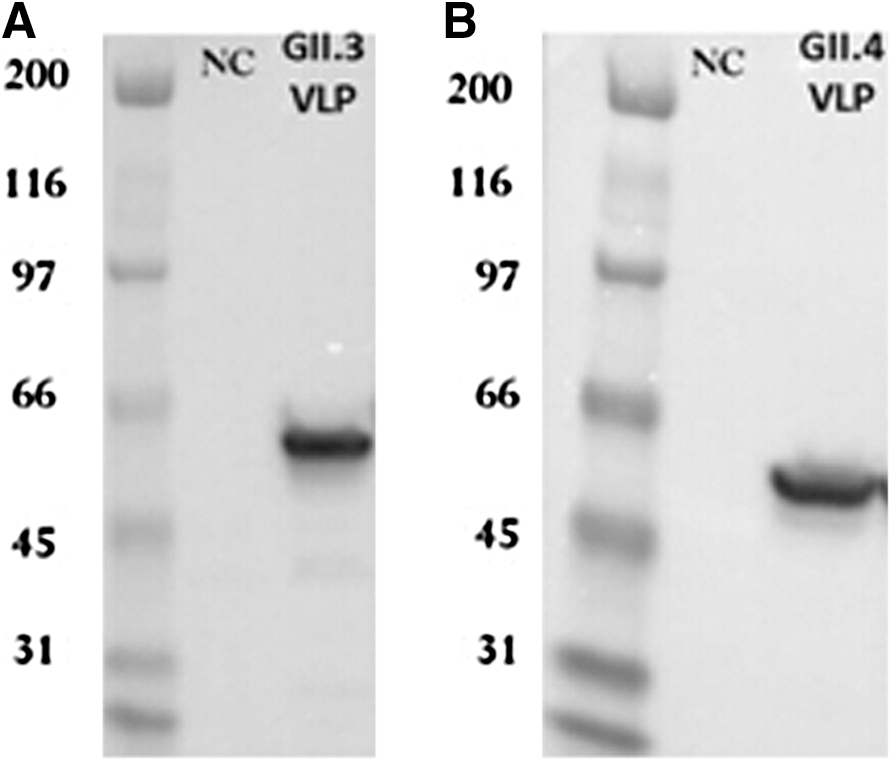
Norovirus viruslike particles (VLPs) detection on sodium dodecyl sulfate polyacrylamide gel.
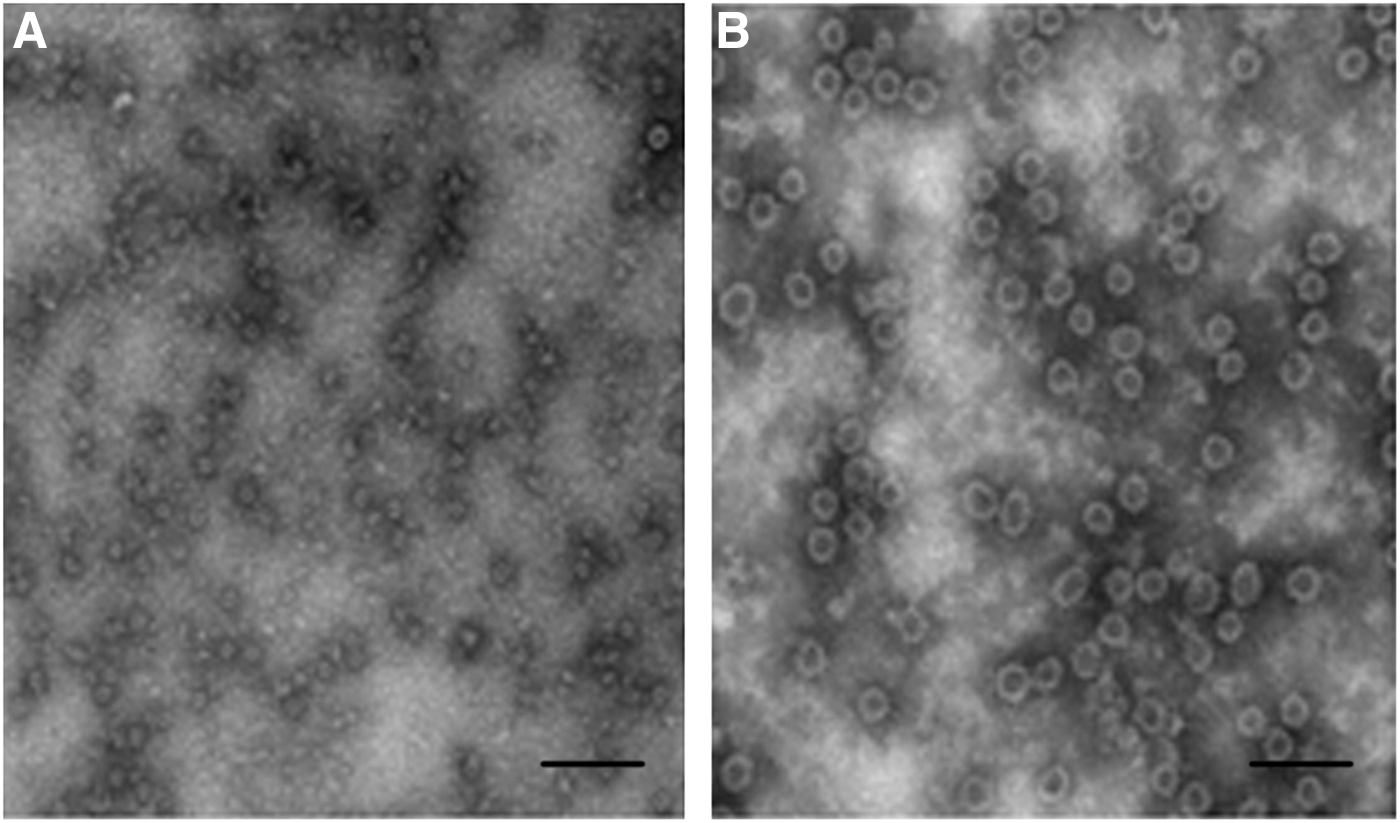
Electron microscopy of sucrose gradient centrifugation purified Norovirus viruslike particles (VLPs) stained with 1% phosphotungstic acid (pH 7.0).
Seroprevalence of NoV GII.3 and NoV GII.4 in children with diarrhea
A total of 362 serum samples (55.2% males and 44.8% females) were collected and 4 age groups were classified: 0–6 months (n=109), 7–11 months (n=121), 12–17 months (n=69), and 18 months–5 years (n=63). The median age of the children was 10 months (ranging 1.5 months–5 years). The overall positivity of anti-GII.4 in all age groups was 86.4%, significantly higher (p<0.01) than 67.9% of anti-GII.3. Of 362 collected samples, 328 tested positive for either anti-GII.3 or anti-GII.4, and 230 were positive for both anti-GII.3 and anti-GII.4.
When seroprevalence was analyzed by age groups, the positive rates of both anti-GII.3 and anti-GII.4 were relatively higher in the <6 months group (69.7% for GII.3 and 88.1% for GII.4) and the 7–11 months groups (71.9% for GII.3 and 89.3% for GII.4). Thereafter, both anti-GII.3 and anti-GII.4–positive rates showed a sharp drop in the 12–17 months group (49.3% for GII.3 and 68.1% for GII.4). Finally, both anti-GII.3 and anti-GII.4–positive rates returned to high levels again (77.8% for GII.3 and 96.8% for GII.4) in the >18 months age group. While GII.3 and GII.4–positive rates were analyzed separately, both anti-GII.3 and anti-GII.4 IgG–positive rates indicated a similar trend: The seroprevalence in the 12–17 months group was the lowest, statistically significantly different (p<0.05) from the other 2 age groups (<11 months and >18 months). Meanwhile, when the positive rates of anti-GII.3 and anti-GII.4 IgG were compared in each age group, the anti-GII.4-positive rate was always higher than that of anti-GII.3 (Fig. 3).
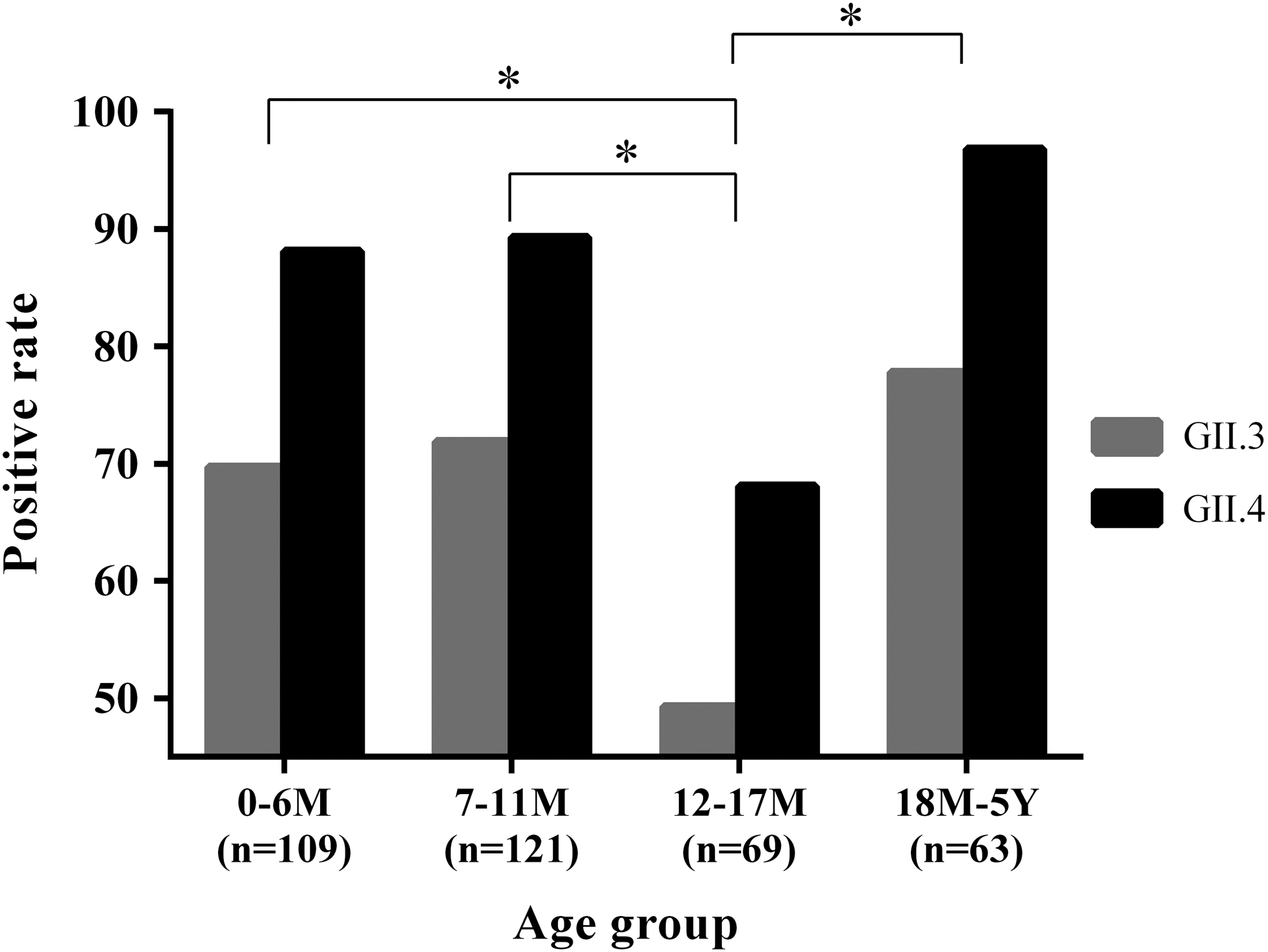
Age-related serum immunoglobulin G (IgG) antibodies to Norovirus GII.3 and GII.4 in hospitalized diarrheal children. Serum samples were tested for anti-GII.3 and anti-GII.4 IgG by enzyme-linked immunosorbent assay using homologous Norovirus viruslike particles expressed from the baculovirus expression system (*p<0.05).
Discussion
In this study, seroprevalence of two NoV genotypes (GII.3 and GII.4) was investigated in hospitalized children under 5 years of age with acute diarrhea in Xi'an, China. The overall seroprevalence in all ages for GII.3 and GII.4 were 67.9% and 86.2%, respectively.
NoV seroprevalence was reported in several countries, and the positive rates varied between geographic locations. For example, anti-GII.4-positive rates were 63.4% (Finland) (Nurminen et al., 2011), 94.5% (Korea) (Son et al., 2013), 66.0% (Japan) (Kobayashi et al., 2009), 72.3% (Portuguese) (Mesquita et al., 2012), 74.1% (France) (Nicollier-Jamot et al., 2003), and 71.1% (Guangzhou of China) (Dai et al., 2004). In comparison, anti-GII.3-positive rates were 76.3% (Korea) (Son et al., 2013), 37.5% (Japan) (Kobayashi et al., 2009), 51.5% (Guangzhou, China) (Dai et al., 2004), and 91.0% (Beijing, China) (Jing et al., 2000). In this study, the overall positive rates of anti-GII.3 and anti-GII.4 in children <5 years (median: 10 months) were 67.9% and 86.2%, respectively, which are in the range of the aforementioned positive rates. The variable seroprevalence of NoV is likely associated with several factors. First, in different locations of the world, hygiene and socioeconomic status are significantly different, and this may lead to a greater NoV seroprevalence. Second, previous molecular epidemiological studies (Cho et al., 2014; Lu et al., 2014) indicated that predominant NoV genotypes varied in different countries, and this probably resulted in an altered NoV seroprevalence. Third, aforementioned studies used varied NoV VLPs to detect serological IgG response; some (Nicollier-Jamot et al., 2003; Kobayashi et al., 2009; Son et al., 2013) applied homologous and others (Jing et al., 2000; Dai et al., 2004; Nurminen et al., 2011; Mesquita et al., 2012) used heterologous antigens. Therefore, the presence of cross-reactive antibodies against epidemic NoV genotypes may lead to biased results. Fourth, the genotype-specific herd immunity in infants and young children and dynamic antigenic mutations are related factors affecting the endemic NoV genotype (Sakon et al., 2014). In recent years, several studies have shown that histo–blood group antigens are associated with the susceptibility to NoV infection. Blood group O had an increased and blood group B had a decreased risk of NoV infection (Hutson et al., 2002; Rockx et al., 2005). The study from our group indicated that NoV-specific IgG titers among children with homozygous and heterozygous secretor were higher than the titer of homozygous weak secretor children (Liu et al., 2014), suggesting that secretor status plays an important role in seroprevalence. In addition to these major factors, other factors such as age span, developing gut microflora, evolving immune system, and sex ratio (Peasey et al., 2004) may also impact NoV seroprevalence.
Comparison of anti-GII.3 and anti-GII.4 positive rates in all the age groups showed that GII.4 seroprevalence was significantly higher than GII.3, suggesting that GII.4 infection is more common than GII.3 in children under 5 years of age in Xi'an, China. This conclusion is consistent with our previous NoV molecular epidemiological results in Xi'an (Zhang et al., 2011; Liu et al., 2014) and studies from other countries (Cheng et al., 2010; Menon et al., 2013; Son et al., 2013).
NoV seroprevalence studies using baculovirus-expressed VLPs have been performed in several countries (Smit et al., 1999; Jing et al., 2000; Dai et al., 2004), but the majority of these studies were reported in the 1990s. In those early studies, all used Norwalk virus (GI.1) and Mexico virus (GII.2) capsid antigens to detect NoV (no matter what epidemic NoV genotypes) serological antibodies and indicated that the prevalence of NoV antibodies followed an age-related pattern. Children appear to acquire antibodies against NoV at an early age, and seroprevalence remains high at older ages; however, NoV GI.1 and GII.2 are not epidemic genotypes anytime and anywhere in the world; therefore, application of heterologous antigens to detect antibodies against epidemic NoV strains may not represent homologous antibody response. To study the virus specific immune response, a homologous antigen should be used. In this study, we used NoV GII.3 and GII.4 VLPs to detect homologous IgG immune response and observed higher positive rates of both GII.3 and GII.4 antibodies in the younger age group (<11 months) compared with children between 12 and 17 months, and NoV antibodies remain at high positive rates in older children (>17 months) thereafter. Interestingly, two NoV genotypes demonstrated a same pattern which was also similar to other studies (Nurminen et al., 2011; Son et al., 2013). The high and stable NoV seroprevalence at an early age (<11 months) revealed that children may have obtained NoV antibodies through their mothers (Gray et al., 1993; Numata et al., 1994) such as breast-feeding (Khodayar-Pardo et al., 2014). After the antibodies waned, the seroprevalence dropped, and that is why lower seroprevalence was detected in the 12–17 months group. Thereafter, in children >17 months, anti-GII.3 and GII.4-positive rates reached high levels.
Immunity to NoV infection is poorly understood. Because NoV cannot be cultivated and there is no small-animal model, our limited knowledge of NoV immunology has been derived from human challenge, outbreak, and pediatric diarrhea studies (Reeck et al., 2010). Human challenge studies suggest that infection with 1 strain of NoV provides short-term (<6 months) (Matsui et al., 2000), but not long-term (>24 months) protection from future infection or illness for the same strain. Infection with one strain may or may not partially protect from illness induced by a second strain, particularly if they belong to separate genogroups (Fankhauser et al., 2002; Rockx et al., 2002). The role of NoV-specific antibodies is also not known. Conflicting reports suggest that the presence of NoV-specific serum antibodies increase, decrease, or does not change the risk of re-infection with NoV.
The major limitation of this study was that all serum samples were collected in the acute course of illness and no follow up samples were obtained. It is likely that the antibodies against the contemporary GII.3 and GII.4 infections have not yet been mounted because NoV specific antibodies are usually detected after 5 days of infection when clinical symptoms end (Brinker et al., 1998, 1999; Iritani et al., 2007). It will be interesting to examine NoV-specific IgG response in serum samples during recovery phase after acute infection and compare it with the antibodies of the acute phase. In the future, we will collect follow-up blood samples and explore cellular immune response in addition to humoral immune response.
Conclusions
Seroprevalence of NoV GII.4 and GII.3 was high in young children, and the anti-GII.4-positive rates were statistically higher than GII.3 across all the age groups. These findings are consistent with NoV molecular epidemiological studies, and it suggests that NoV infection is very common and young children are susceptible to NoV infections, although there is a high NoV seroprevalence.
Footnotes
Acknowledgments
The authors gratefully acknowledge the generous assistance of the staff members of the Department of Digestive Diseases of Xi'an Children's Hospital. This project supported by the National Natural Science Foundation of China (Grant No. 81101333).
Disclosure Statement
No competing financial interests exist.