
Abstract
This study investigated the survival of Escherichia coli O157:H7 and Salmonella Typhimurium in finished dairy compost with different particle sizes during storage as affected by moisture content and temperature under greenhouse conditions. The mixture of E. coli O157:H7 and S. Typhimurium strains was inoculated into the finished composts with moisture contents of 20, 30, and 40%, separately. The finished compost samples were then sieved into 3 different particle sizes (>1000, 500–1000, and <500 μm) and stored under greenhouse conditions. For compost samples with moisture contents of 20 and 30%, the average Salmonella reductions in compost samples with particle sizes of >1000, 500–1000, and <500 μm were 2.15, 2.27, and 2.47 log colony-forming units (CFU) g–1 within 5 days of storage in summer, respectively, as compared with 1.60, 2.03, and 2.26 log CFU g–1 in late fall, respectively, and 2.61, 3.33, and 3.67 log CFU g–1 in winter, respectively. The average E. coli O157:H7 reductions in compost samples with particle sizes of >1000, 500–1000, and <500 μm were 1.98, 2.30, and 2.54 log CFU g–1 within 5 days of storage in summer, respectively, as compared with 1.70, 2.56, and 2.90 log CFU g–1 in winter, respectively. Our results revealed that both Salmonella and E. coli O157:H7 in compost samples with larger particle size survived better than those with smaller particle sizes, and the initial rapid moisture loss in compost may contribute to the fast inactivation of pathogens in the finished compost. For the same season, the pathogens in the compost samples with the same particle size survived much better at the initial moisture content of 20% compared to 40%.
Introduction
O
Previous studies have provided evidence that pathogens were either reduced or eliminated during the composting process (Shepherd et al., 2007; Chen and Jiang, 2014). However, there are still some possibilities for pathogens to survive during composting because of the heterogeneity of compost heaps, manure types, moisture content, and variations among different seasons (Shepherd et al., 2007; Erickson et al., 2010; Shepherd et al., 2010b). Foodborne pathogens can even survive for up to 4 months on the compost surface in animal waste compost (Shepherd et al., 2010a). Additionally, some studies have reported that pathogens reintroduced or surviving in finished compost have the ability to regrow to high levels in dairy manure–based compost under favorable conditions (Kim et al., 2010). Therefore, the contaminated compost can play an important role in the presence of foodborne pathogens on fresh produce (Oliveira et al., 2011), and there is a need to investigate how foodborne pathogens in manure or compost are transmitted to fresh produce during the preharvest stage.
Bioaerosols consist of inert particles and the attached microorganisms (Le Goff et al., 2010), and they vary greatly in size and dispersion ability (Dowd et al., 2000). A few studies have reported that bioaerosol generation, transportation, and pathogen contamination are associated with the composting operation (Fischer et al., 2008; Coccia et al., 2010; Ravva et al., 2011). Small compost particles on the compost heap surface may travel through air as aerosols from the composting site to a vegetable production site nearby. Consequently, some pathogenic cells may become airborne by adhering to compost particles (Coccia et al., 2010), which provides one of the routes for pathogen transmission on farms. Due to the heterogeneity of compost materials, it is expected that different sizes of compost particles coexist. However, no study has been carried out to examine the relationship between compost particle size and the survival of pathogens in those particles. Knowledge about pathogen survival on different compost particle sizes can be critical to assess the risk of compost aerosol contamination under the farm environment. Furthermore, pathogen survival in compost can be affected by many other factors such as moisture content, temperature, rain event, seasonality, and so on (Sidhu et al., 1999; Kim and Jiang, 2010). Therefore, the purpose of this study was to investigate the survival of E. coli O157:H7 and Salmonella in finished dairy compost during storage as affected by compost particle size, initial moisture content, and temperature under greenhouse conditions.
Materials and Methods
Preparation of finished dairy compost
The commercial Black Kow finished dairy compost (Black Gold Compost Company, Oxford, FL) was used in this study. This product contained 0.5% total nitrogen, 0.5% available phosphate (P2O2), 0.5% soluble potash (K2O), and no more than 1% chlorine. The finished compost sample was dried under a chemical hood until the moisture content was decreased to <20%. This dried sample was then prescreened using a sieve (sieve pore size, 3×3 mm) to a particle size of <3 mm to minimize sample heterogeneity. Sufficient samples were collected for the entire experiment and stored in a sealed container at 4°C until use. Initial moisture content of the finished compost sample was measured with a moisture analyzer (Model IR-35 Infrared Analyzer; Denver Instrument, Denver, CO). The pH of the compost sample was determined by adding 1 g sample in 50 mL distilled water, then stirred for 2 min, and measured by a pH meter (Orion Star 2-star meter; Thermo Scientific, Beverly, MA) (Jiang et al., 2002). Background microorganisms in the compost sample were enumerated in duplicate on tryptic soy agar (TSA; Becton, Dickinson and Company, Sparks, MD), and compost samples were also tested for the presence of either E. coli O157:H7 or Salmonella as described by the U.S. Food and Drug Administration's Bacteriological Analytical Manual.
Morphological and surface analysis of compost particles
The above compost sample was sieved into 3 portions with particle sizes of >1000, 500–1000, and <500 μm using standard sieves (#18 and 35, VWR, Radnor, PA). Then, each portion was observed under a Contour GT-K1 Optical Profiler (Bruker Corporation, Billerica, MA) using the automated turret and programmable X, Y, Z movement. The diameter, area, and surface roughness for each sample were measured in triplicate.
Bacterial cultures
Avirulent, ampicillin-resistant, and green fluorescent protein–labeled E. coli O157:H7 strain B6914 (kindly provided by Dr. Pina Fratamico at the U.S. Department of Agriculture, Agricultural Research Service—Eastern Regional Research Center) and avirulent S. Typhimurium strain 8243 (kindly provided by Dr. Roy Curtis III, Washington University, St. Louis, MO) were used. The S. Typhimurium strain was induced to be resistant to 100 μg of rifampin mL–1 (Fisher Scientific, Fair Lawn, NJ) using the gradient plate method (Smith et al., 1982). Bacterial cultures were stored at −80°C in tryptic soy broth (TSB; Becton Dickinson) with 20% glycerol.
Inoculum preparation
The frozen stock cultures of S. Typhimurium and E. coli O157:H7 were thawed and streaked on TSA supplemented with 100 μg mL–1 of rifampin (TSA-R) and ampicillin (TSA-A), respectively, and grown at 37°C for 24 h. Single colonies of S. Typhimurium and E. coli O157:H7 on TSA-R or TSA-A were transferred into 50 mL of TSB supplemented with rifampin (TSB-R) or ampicillin (TSB-A), respectively, and incubated in a shaking incubator (100 rpm, 37°C) for 24 h. The overnight cultures were washed 3 times by centrifugation at 8000 rpm for 10 min, and the final pellet was resuspended in 0.85% saline to desired cell concentrations of ≈109 CFU mL–1 by measuring the optical density at 600 nm.
Finished compost inoculation and placement
The moisture content of the above finished compost samples was measured as described above. The autoclaved tap water was used to adjust the moisture contents to 20, 30, and 40% as commonly found in compost products. The finished compost samples were inoculated with E. coli O157:H7 and Salmonella cultures, separately, at a ratio of 1:100 (vol:wt) to yield a final concentration of ≈107 CFU g–1 (Fig. 1). The moisture content was adjusted to the target initial level by adding the calculated amount of sterile tap water and inoculum. The inoculated compost samples were continually mixed by hands wearing sterile gloves (sterile nitrile gloves; Kimberly-Clark, Neenah, WI) for 15 min under a biological hood. The above compost samples of 20 and 30% moisture contents were then sieved into 3 portions: >1000, 500–1000, and <500 μm, whereas compost sample with 40% moisture content was separated into >1000 and <1000 μm portions due to large particle size formed in compost with this higher moisture content. A portion of 60-g sample was placed into a plastic tray (4×4 cm) at a depth of 3–4 cm. Duplicate samples were prepared and stored in a greenhouse up to 1 month without further adjustment of moisture content. The experiment was conducted during November–December 2011, January–February 2012, and July–August, 2012 for the late fall, winter, and summer trials, respectively. For each season, the second trial was initiated 1 week after the beginning of the first trial.
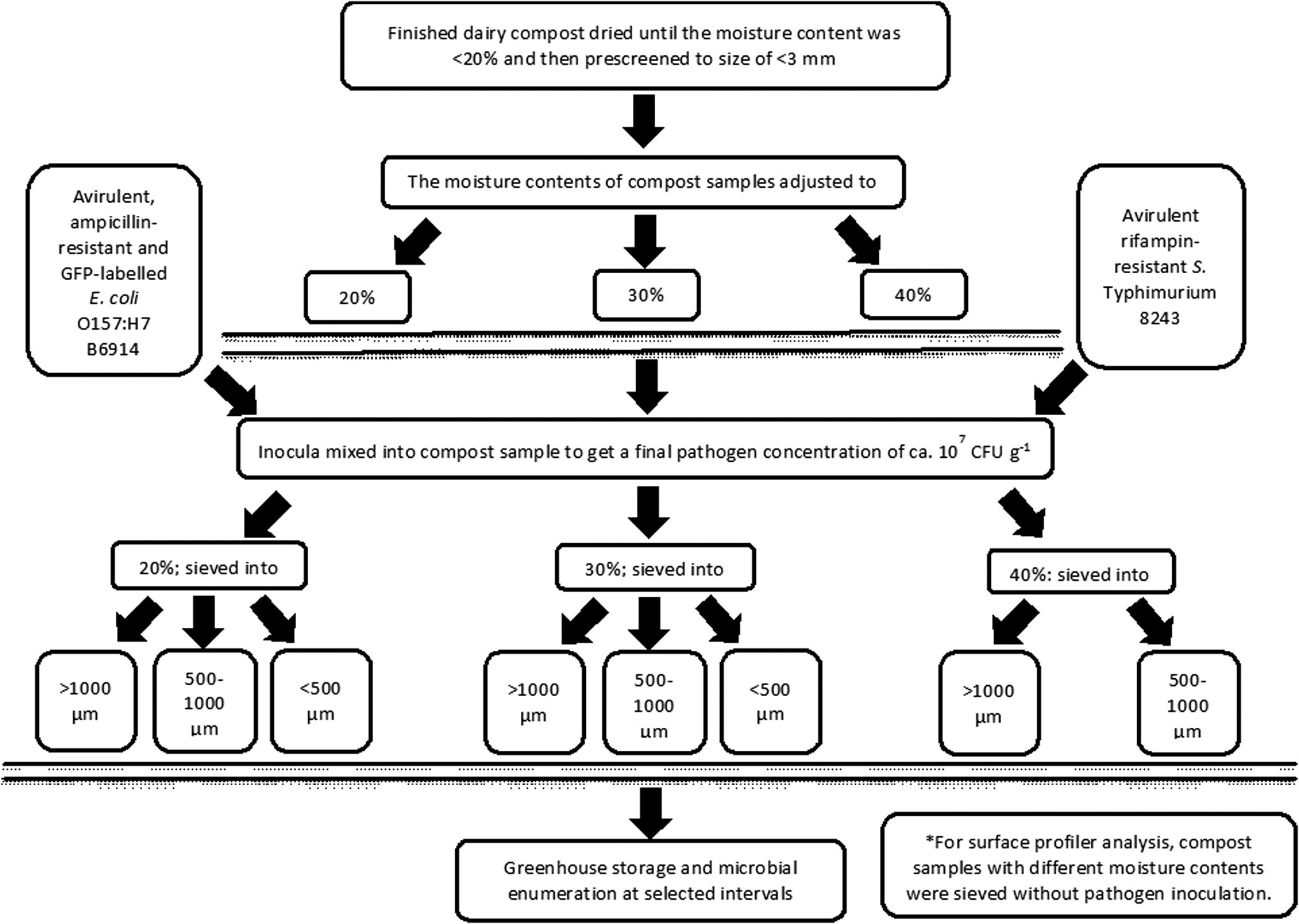
Flow chart of the experimental procedure. GFP, green fluorescent protein; CFU, colony-forming unit.
Greenhouse conditions
Temperature and relative humidity (RH) in the experimental unit in the greenhouse were measured and recorded by the remote monitoring system (Argus control system Ltd.; White Rock, BC, Canada). The greenhouse room used in this study was set to have an upper limit of 30.5°C and 700 W m–2 for temperature and light intensity, respectively. Due to the fact that the greenhouse was designed for plant growth, the low temperature control limit was set at 16°C. Forced ventilation was used to control the temperature when the temperature was too high and shading was occasionally used to control the light intensity.
Microbiological analysis
Finished compost samples were taken and analyzed on day 0, 1, 3, 5, 10, 15, 20, and 30 during storage. Briefly, 5 g of sample were mixed with 45 mL of universal pre-enrichment broth (UPB; Becton Dickinson and Company, Sparks, MD) in a Whirl-Pak stand-up bag (Nasco, Fort Atkinson, WI) with the ratio of 1:10 and shaken vigorously for 1 min. Then, the homogenates were serially diluted in sterile saline and plated on TSA-A and xylose lysine Tergitol-4 agar supplemented with 100 μg mL–1 of rifampin (XLT-4-R) in duplicate using an Autoplate 4000 spiral plater (Spiral Biotech Inc., Bethesda, MD) for E. coli O157:H7 and Salmonella, respectively. Afterwards, the plates were incubated at 37°C for 24 h and 48 h for TSA-A and XLT-4-R, respectively.
When the direct plating method failed to detect any viable cells (detection limit of 1.30 log CFU g–1), the samples were enriched in UPB at 37°C for 24 h, followed by the selective enrichment in TSB-A or Rappaport-Vassiliadis broth with rifampin (RV-R) at 37°C for E. coli O157:H7 and Salmonella, respectively. After incubation for 24 h, a loopful of secondary enrichment culture was streaked onto TSA-A and XLT-4-R for E. coli O157:H7 and Salmonella, respectively. Selected colonies grown on each selective media were confirmed by E. coli O157:H7 and Salmonella latex agglutination tests (Oxoid Inc., Basingstoke, Hampshire, UK).
Statistical analysis
For each season, there were two separate trials, and experiments were conducted in duplicate for each trial. Total bacterial counts were converted to log CFU g–1 on dry-weight basis, and the log reductions of each pathogen within first 5 days and after 30 days in each trial were subjected to analysis of variance, followed by the least significant difference analysis with Statistical Analysis System (SAS, version 9.1; SAS Institute Inc., Cary, NC). The difference among treatments was considered significant with a p-value of<0.05. In this study, 3 factors being examined were seasonality (summer, late fall, and winter), moisture content (20, 30, and 40%), and particle size (>1000, 500–1000, and <500 μm). Also, the correlation between pathogen reduction and dehydration rate within 5 days in different seasons was analyzed using SAS. A linear regression model was used to predict the change of E. coli O157:H7 and Salmonella reduction according to the dehydration rate in different seasons.
Results
pH, microbiological, and morphological and surface analysis of finished compost
Initial pH values for finished compost samples used in late fall, winter, and summer trials were 7.93±0.09, 7.66±0.01, and 7.77±0.04, respectively. The levels of background mesophilic microorganisms in these samples ranged from 6 to 8 log CFU g–1. All compost samples for these trials were negative for both E. coli O157:H7 and S. Typhimurium.
The finished compost samples of particle sizes of >1000, 500–1000, and <500 μm had an average diameter of 2037, 884, and 257 μm, respectively, and the average surface roughness (Ra) of 112.66, 76.61, and 26.45 μm, respectively. Ra was defined as the arithmetic average of the absolute values of the heights of roughness irregularities (Ho et al., 2002). The average areas were 1.19×106, 2.91×105, and 3.70×104 μm2 for particle sizes of >1000, 500–1000, and <500 μm, respectively. Data of the profiler analysis are summarized in Table 1. The results for sample particle sizes were consistent with the sieve pore sizes, which were 500 and 1000 μm.
Ra, represents the surface roughness. It is the arithmetic average of the absolute values of height difference.
Area and volume represent the measurement per particle.
Diameter is the average measurement of particle length.
Greenhouse conditions
To simulate the surface condition for finished compost during outdoor storage, three greenhouse trials were performed during November–December 2011 (late fall trial), January–February 2012 (winter trial), and July–August 2012 (summer trial). The average values of temperature in the greenhouse were 21.3, 21.7, and 28.4°C for late fall, winter, and summer trials, respectively, while the average values of RH in the greenhouse were 41.3, 36.0, and 64.4% for late fall, winter, and summer trials, respectively. Figure 2 presents the average temperature, RH, and light intensity inside the greenhouse on a typical day of each trial.
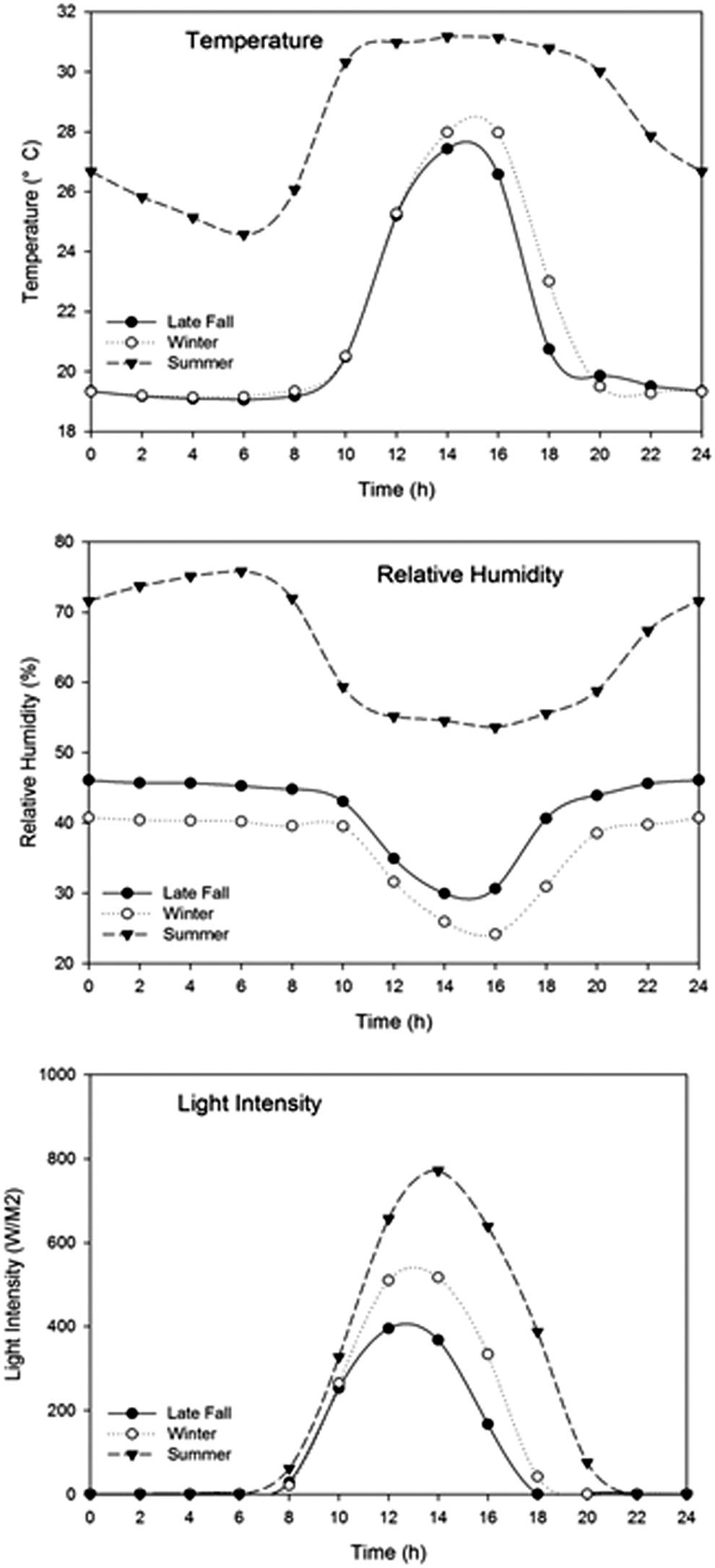
Greenhouse temperature, relative humidity and light intensity, in a typical day. The data were recorded hourly by the remote monitoring system starting at midnight. For each trial, data for each parameter of every experimental day were averaged.
Changes of moisture content in compost stored in different seasons
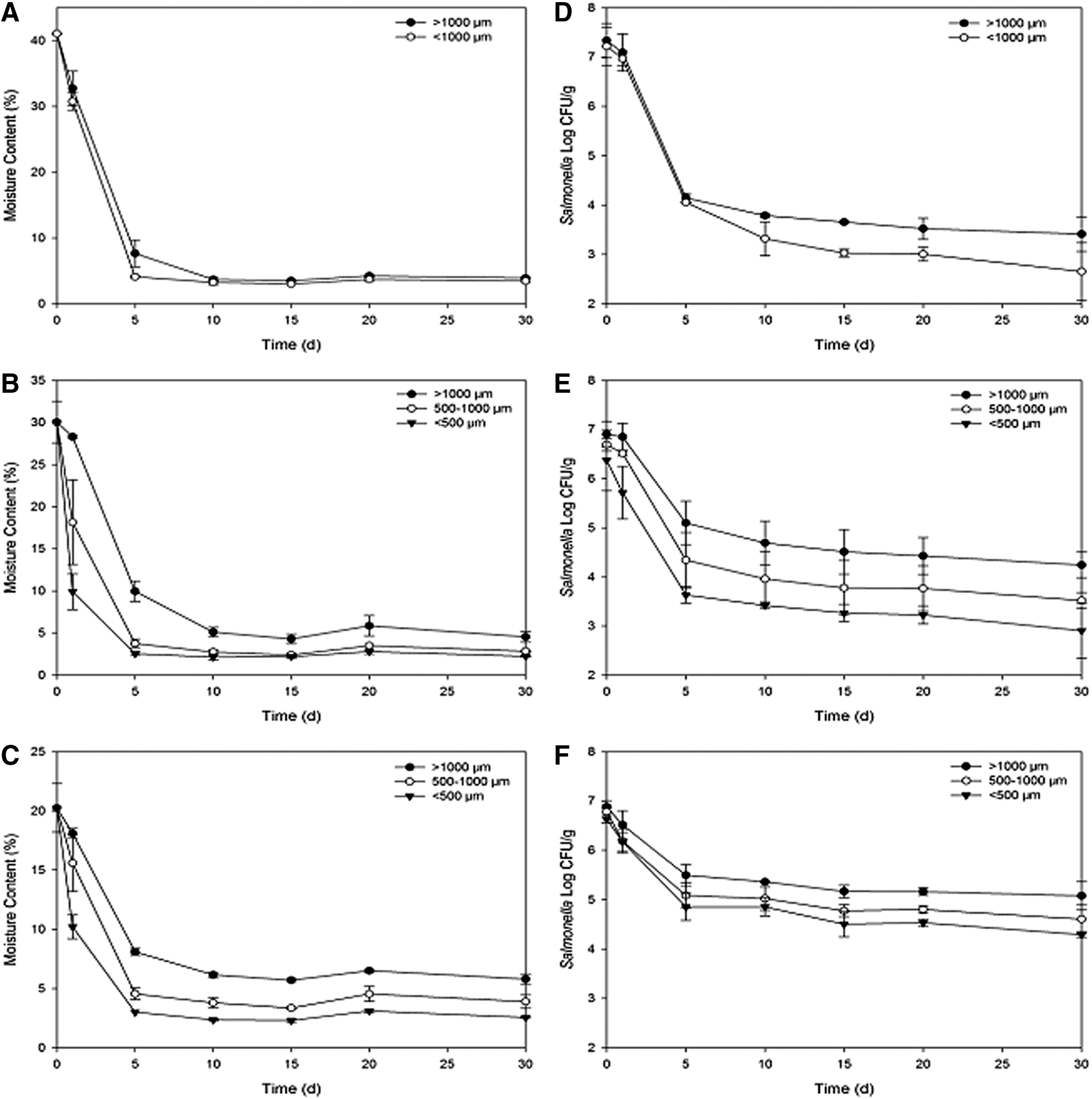
The experiments for all trials were conducted with no further moisture adjustment after the compost samples were placed in the greenhouse. The moisture change in each sample during 1-month storage is shown in Figure 3 for the late fall trial only. In general, the moisture contents of compost samples were reduced from 40, 30, and 20% to <10% within 5 days during late fall and winter trials. After 5 days, the moisture content stayed below 10% in these trials. For the summer trial, the moisture contents of compost samples with <500 μm particle size reduced from the initial moisture contents of 30 and 20% to <10% within 5 days, whereas for compost samples with >1000-μm particle size, the moisture content increased ≈5% within 1 day, followed by a rapid decline, and then stayed at 5–25% from days 5 to 30 (data not shown).
Changes in moisture content
For the same initial moisture level, the moisture content of compost samples with larger particle sizes tended to decrease more slowly than those with smaller particle sizes (Fig. 3 and Table 2). For example, in the compost samples with >1000-μm particle size and 30% initial moisture content in the late fall trial, it had a dehydration rate of 4.02% per day on average within the first 5 days, as compared to that of 5.26 and 5.50% per day in the compost samples of particle sizes of 500–1000 and <500 μm, respectively. For samples with 30% initial moisture content in the winter trial, there were dehydration rates of 4.48, 4.99, and 4.90% per day in the compost samples of >1000, 500–1000, and <500-μm particle sizes, respectively. Moreover, in the compost samples with 30% initial moisture content in the summer trial, the >1000, 500–1000, and <500-μm samples had dehydration rates of 1.69, 3.82, and 4.70% per day within 5 days, respectively. Clearly, the finished compost samples with the same particle size and moisture level dried out much more quickly in the late fall and winter trials than the summer trial.
Data are expressed as means±SD of two trials. Means of different uppercase letters in the same column within each season and moisture content are significantly different (p<0.05) according to the least significant difference (LSD) test. Means of different lowercase letters in the same column with the same particle size in each season are significantly different (p<0.05) according to the LSD test.
The dehydration rates were calculated for all compost samples within 5 days, but for winter trial, dehydration rates within 3 days were presented as well.
Dehydration rate is expressed as moisture content reduction/days of storage.
E. coli, Escherichia coli; NT, not tested.
Survival of S. Typhimurium and E. coli O157:H7 in finished compost
The initial inoculation levels for S. Typhimurium and E. coli O157:H7 were at 6.5–7.5 log CFU g–1. Under greenhouse conditions, both S. Typhimurium and E. coli O157:H7 populations declined rapidly within 5 days, followed by a gradual reduction during the 1-month storage. Thus, the log reduction in the first 5 days was used to analyze the inactivation rates of both pathogens.
Table 2 summarizes the pathogen reductions and dehydration rates in finished compost samples during the initial 5 days of storage for three seasons as affected by compost particle size and initial moisture content. Table 3 summarizes the pathogen reduction in finished compost samples during entire storage as affected by compost particle size and initial moisture content. Figure 2 presents the changes in moisture content and Salmonella reductions in compost particles for the late fall trial. Generally, within each season, as the compost particle size increased, the pathogen inactivation rate decreased; and as the initial moisture content increased, the pathogen inactivation rate increased for the compost particle with the same size with some exceptions for the winter trial. Salmonella population decreased by 1.80, 2.35, and 2.74 log CFU g–1 within 5 days for the compost samples (30% initial moisture content) of particle sizes of >1000, 500–1000, and <500 μm, respectively, as compared to the pathogen reductions of 1.39, 1.70, and 1.78 log CFU g–1 for the compost samples with 20% initial moisture content, respectively (Table 2).
Data are expressed as means±SD of two trials. Means of different uppercase letters in the same column within each season and moisture content are significantly different (p<0.05) according to the east significant difference (LSD) test. Means of different lowercase letters in the same column with the same particle size in each season are significantly different (p<0.05) according to the LSD test.
E. coli, Escherichia coli; NT, not tested.
Both Salmonella and E. coli O157:H7 in finished compost samples with larger particle size survived better than those with smaller particle sizes. Within the same season, these pathogens in the compost samples with the same particle size survived better at the initial moisture content of 20% when compared with 40%. During the initial 5 days of storage (Table 2), for the late fall trial, the log reduction of Salmonella in the compost sample with 40% moisture content was significantly higher (p<0.05) as compared to 30 and 20% moisture contents when particle size was >1000 μm. For the winter trial, the log reductions of Salmonella in compost sample of the particle size of <500 and 500–100 μm were significantly higher as compared to the particle size of >1000 μm (p<0.05) when moisture content was 30%. The log reductions of E. coli O157:H7 in the compost sample became higher as particle size decreased (p<0.05) when moisture content was 30%. For the summer trial, the log reductions of Salmonella in compost sample with the particle size of >1000 μm were significantly lower as compared to the particle size of <1000 μm (p<0.05) when moisture content was 40%.
For the late fall trial, the log reduction of Salmonella in compost sample with the particle size of <500 μm was significantly higher (p<0.05) as compared to the particle size of >1000 μm when moisture content was 30% (Table 3). The log reduction of Salmonella in compost sample with 30% moisture content was significantly higher (p<0.05) as compared to 20% moisture contents when particle size was <500 μm. The log reduction of Salmonella in compost sample with 40% moisture content was significantly higher (p<0.05) as compared to 20% moisture contents when particle size was >1000 μm. For the winter trial, the log reduction of Salmonella in compost samples of the particle sizes of <500 and 500–100 μm were significantly higher (p<0.05) as compared to the particle size of >1000 μm when moisture content was 20%.
Salmonella was studied for the summer, late fall, and winter trials, whereas E. coli was tested only in the summer and winter trials. Based on the overall pathogen reduction data, even though E. coli was not tested in late fall, Salmonella inactivation rates were affected by three seasons differently. Overall, both pathogens were inactivated relatively faster in winter than in summer.
The correlations between pathogen reduction and particle size or moisture content were not high (R 2<0.70). The correlation between the dehydration rate of compost and Salmonella log reduction in late fall, Salmonella log reduction in summer, or E. coli O157:H7 log reduction in summer were 0.98, 0.90, and 0.79, respectively (Table 4). However, the correlations between the dehydration rate of compost and Salmonella and E. coli O157:H7 log reduction in winter were −0.05 and −0.12, respectively. The correlation results showed that in late fall and summer trials, the relationship of pathogen reduction and dehydration rate can be described as a linear regression model, whereas in the winter trial, the change in the pathogen log reduction was not linear with the dehydration rate.
R2 represents the ability of dehydration rate to predict the log reductions of pathogens.
X variable represents the value log reduction of pathogen change when the dehydration rate rise by 1%/d.
p-Value represents the significance of X variable.
E. coli, Escherichia coli.
Discussion
In this study, the pathogen survival curves showed a rapid decline of bacterial population in the finished compost, followed by an extensive tailing after the first 5 days of storage time regardless of season and initial moisture content. Kim et al. (2010) found that after 7 days, the populations of E. coli O157:H7, Salmonella spp., and Listeria monocytogenes in dairy compost stayed stable, showing an obvious tailing effect. This effect may be explained by the fact that more sensitive populations of these pathogens were killed by harsh environmental stresses, while a few resistant cells were able to survive and persist under the same stresses. The prolonged survival of pathogens in compost may also be attributed to the low nutrients/starvation and desiccation stress adaptation phenomenon (Shepherd et al., 2010b; Chen et al., 2013; Chen and Jiang, 2014).
Moisture content of compost is considered to be an important factor affecting pathogen survival and growth (Sidhu et al., 1999). It is well documented that a fast dehydration process leads to an efficient microbial inactivation (Antheunisse and Arkesteijndijksman, 1979). In our study, the relationship between dehydration rate and pathogen inactivation within 5 days supported this assumption (Table 2). After initial rapid moisture loss in finished compost samples, pathogens were exposed to constant desiccation stress. Since the pathogen cells can become adapted to the dry condition after 5 days of storage in a greenhouse, these cells may develop the ability to survive for an extended period of time in dry compost samples (Crowe et al., 1992; Potts, 1994). The lower level of overall survival of pathogens in the winter trial might be explained by the higher dehydration rate compared to other two seasons (Table 2). It should be noticed that, for the samples with particle size of >1000 μm, pathogens generally died more rapidly in samples with higher initial moisture content. This may be due to the fact that the higher initial moisture content in compost samples caused more rapid dehydration in the first 3 or 5 days of storage (Table 2).
Temperature is considered as another factor influencing pathogen survival and growth in finished compost (Singh et al., 2010). In this study, the average ambient temperatures in all trials were lower than the optimum growth temperature for Salmonella and E. coli O157:H7. Pietronave et al. (2004) investigated the survival and growth of seeded Salmonella and E. coli O157:H7 in finished compost with different indigenous microflora, moisture contents, and temperature conditions. Their results demonstrated that in nonsterilized samples with 10% moisture content, Salmonella stored at room temperature had 0.74 log lower population compared to those stored at 37°C, and the difference was smaller (0.17 log) when the moisture contents were 40%. In agreement with this published study, our results also revealed a larger difference in the reduction of Salmonella population in samples with 20% moisture content among 3 seasons than in samples with 40% moisture content, suggesting the temperature as a significant factor on pathogen survival in compost samples of different initial moisture contents. In this study, the greenhouse facility was designed for supporting the plant growth; therefore, the lowest temperature was set to 16°C. Due to such limitation in greenhouse setting, the average temperatures in late fall and winter trials were very close. Our results of the winter trial may have some implications for some regions with warm winters, such as California rather than South Carolina. In addition to average temperature, other parameters such as light intensity, humidity, and temperature fluctuations between day and night varied a lot among three seasons tested. As a result, the pathogen inactivation rates were different, suggesting some of the above factors could affect the pathogen survival in the finished compost. In the present study, the seasonality effects on pathogen survival were mainly influenced by relative humidity and light intensity, which could affect the dehydration rates of finished compost samples.
Particle size is also a key factor for the pathogen carriage and transfer. On compost sites, aerosols can be generated from the surface of the compost pile and dispensed as small airborne aerosols (Gralton et al., 2011). The influence of particle sizes of finished compost samples on the survival of pathogens was also examined in this study. Our results demonstrated that pathogens died the most rapidly in the compost samples with particle size of <500 μm, although some differences in population reduction among different particle sizes for Salmonella and E. coli O157:H7 were not significant (p>0.05) for the same treatment. Some previous studies have determined the relationship between the matrix size of soil sediments and the pathogen survival profile in them. Howell et al. (1996) conducted a laboratory-scale study to determine the survival of fecal bacteria in the sediment of agricultural water. They reported that the mortality rate of fecal bacteria decreased as the particle size decreased, which are contradictory to our findings. This difference may be attributed to the fact that these sediment samples were investigated in an aqueous environment, whereas our compost samples were exposed to air under greenhouse conditions. Since pathogens can attach to the particle surface, the complex or rough surface structure can prevent pathogenic cells from being washed away by wind or water. In this study, the “Ra” results from the surface profiles suggested that larger particles had larger average surface area per particle than smaller particles. The increase in particle size can also lead to more nutrients and pathogens being shielded among or inside particles, less water loss, higher air-holding capacity, and less temperature fluctuation during compost storage.
The primary challenge in understanding the survival of foodborne pathogen–laden compost bioaerosols lies in delineating different factors influencing the survival characteristic. The present study investigated the survival profiles of foodborne pathogens in compost by taking into consideration compost particle size, moisture content, and temperature. Since our study was conducted under greenhouse conditions, the results may not reflect the actual situation of outdoor compost storage in composting facilities. Nonetheless, the data provide some valuable information about the possibilities of pathogen survival in finished compost as affected by particle size, moisture content, temperature, relative humidity, and light intensity. Since pathogens surviving and persisting on the surface of a compost heap during storage may be disseminated into nearby produce fields via aerosols (Fischer et al., 2008), the surface of finished compost may be treated with physical covering to prevent compost particles from forming an aerosol (Shepherd et al., 2011). Therefore, the composting facilities should be located at a certain distance away from fresh-produce farms. However, to simulate the real-world condition of composting facilities, further studies are needed to investigate the ability and risk of the transfer of compost particle of different particle sizes in open-air environments.
In conclusion, our results suggested that finished compost samples with larger particle sizes supported pathogen survival for an extended time as compared with compost samples with smaller particle sizes. In addition, for the same particle size, the pathogen survived better in compost with lower moisture content. Under greenhouse conditions, a higher dehydration rate was found to be a critical factor contributing to the initial rapid inactivation of pathogens. In consideration of the extended survival of a few pathogenic cells in finished compost particles, the risk of compost bioaerosols as one of the possible sources of produce contamination needs to be further assessed under field conditions.
Footnotes
Acknowledgments
We would like to thank Dr. James Rieck (Department of Mathematical Sciences, Clemson University) for assistance with statistical analysis. This research was supported by a grant from Center for Produce Safety, University of California at Davis.
Disclosure Statement
No competing financial interests exist.