
Abstract
Morin, a plant-derived flavonol, is known to be an effective inhibitor of Gram-positive bacteria. In this study, we explored the combined effect of morin with β-lactam antibiotics against methicillin-resistant Staphylococcus aureus (MRSA), a multidrug-resistant pathogen. The anti-MRSA activity of morin was investigated by the broth microdilution method, checkerboard dilution test, and time-kill curve assay. The expression of the resistant protein, penicillin-binding protein (PBP2a) encoded by mecA, was analyzed by the Western blotting method in the presence of morin and oxacillin. An increased susceptibility of MRSA toward oxacillin was observed in the presence of morin. The protein level of PBP2a was reduced when MRSA (ATCC 33591) was treated with the combination of morin and oxacillin, indicating that the combination of morin and oxacillin potentiates the killing effect against MRSA. The present study indicates that the killing effect by the combinative treatment of morin and β-lactam antibiotic is dependent on the PBP2a-mediated resistance mechanism.
Introduction
M
Among the plant flavonoids, morin (2′,3,4′,5,7-pentahydroxyflavone) was originally isolated from the members of the Moraceae family (Kapoor and Kakkar, 2012) (Fig. 1). Morin exhibits many biological activities including antioxidant (Prahalathan et al., 2012), anti-inflammatory (Fang et al., 2003), and especially osteoprotective effects on diabetic-induced osteopenia via both the antioxidant and anti-inflammatory effects. Pharmacological properties of morin such as anticancer (Gupta et al., 2013), antihyperlipedimia (Abuohashish et al., 2013), and antibacterial (Woźnicka et al., 2013) have also been reported. Gupta et al. reported that morin suppresses the signal transducer and activator of transcription 3 (STAT3) pathway in tumor cells (Gupta et al., 2013). Woźnicka et al. reported that the sulfonic derivatives of morin possess the antibacterial activity on the bacterial strains of Escherichia coli, Pseudomonas aeruginosa, and S. aureus (Woźnicka et al., 2013). However, the anti-MRSA activity of morin combined with existing antibiotic associated with the problem of antibiotic resistance was not studied.
Chemical structure of morin.
This study focuses on the development of novel antibiotics to reduce bacterial resistance in combination with conventional drugs. We investigated the enhanced antimicrobial effectiveness of morin combined with β-lactam antibiotics on MRSA.
Materials and Methods
Reagents
Ampicillin, oxacillin, and morin (C15H10O7) were purchased from Sigma-Aldrich Co. (St. Louis, MO). Difco™ Mueller–Hinton broth (MHB) was used as the nutrient media. Mouse anti-PBP2a was purchased from DiNonA Inc., Seoul, Korea.
Bacterial strains and growth conditions
Among the six strains of S. aureus used in this study, five clinical MRSA isolates were obtained from five different patients at Wonkwang University Hospital. The one remaining strain, S. aureus ATCC 33591 (methicillin-resistant strain), was commercially purchased from American Type Culture Collection, Manassas, VA. All the bacteria were stored in 30% glycerol and frozen at −70°C until use. The bacterial strains were suspended in MHB and incubated in Mueller–Hinton agar (MHA) at 37°C for 24 h.
Determination of minimum inhibitory concentration (MIC)
The MIC was determined using the broth microdilution method described by the Clinical and Laboratory Standard Institute guidelines (CLSI, 2014). Serial twofold dilutions of morin in MHB were prepared in sterile 96-well microplates and microtubes. The MRSA inocula were adjusted to the 0.5 McFarland standard (1.5×108 colony-forming units [CFU]/mL) in MHB. Final inocula were adjusted to 1.5×105 CFU/well. The MIC was defined as the lowest concentration at which visible growth inhibition was observed after 24 h incubation at 37°C.
Combination test
The checkerboard method was used to identify the interactions between morin and β-lactam antibiotic (Mun et al., 2014). The antimicrobial assay was performed with morin in combination with ampicillin and oxacillin. Serial dilutions of morin with these antibiotics were mixed in the cation-supplemented MHB. The inocula were prepared from the colonies that were grown in MHA overnight. The final bacterial concentration after the inoculation was 1.5×105 CFU/well. The MIC, determined after the incubation at 37°C for 24 h, was defined as the lowest concentration of drug, alone or in combination with other agents, that visibly inhibited the growth of bacteria. Each experiment was performed three times. The in vitro interaction between the drugs was quantified by determining the fractional inhibitory concentration (FIC). The FIC index (FICI) was calculated using the following formula:
where [A] is the concentration of drug A, MICA and FICA are the MIC and FIC of drug A, respectively, [B] is the concentration of drug B, and MICB and FICB are the MIC and FIC for drug B, respectively. The FICI thus obtained was interpreted as follows: Values <0.5 represent synergy; 0.5–0.75, partial synergy; 0.76–1, an additive effect; 1.1–4, no effect; and >4, antagonism. Finally, the different values of synergy between the two agents were calculated.
Time-kill curve assay
To perform the viability assay, the synergistic effects of morin and β-lactam antibiotics were determined on the bacterial growth in 96-well plates at 5 different points of time (0, 4, 8, 16, and 24 h) (Mun et al., 2014). The bacterial cultures incubated in MHA for 24 h at 37°C were diluted with fresh MHB to 1.5×105 CFU/mL, and the diluted cultures were preincubated at 37°C for 24 h. Aliquots (0.1 mL) of the culture were taken at 0, 4, 8, 16, and 24 h of incubation, and serial 10-fold dilutions were prepared in saline as needed. The numbers of viable cells were determined on a drug-free MHA plate after incubation for 24 h. Colony counts were performed on plates, and 30–300 colonies were enumerated. The lower limit of sensitivity of the colony counts was 100 CFU/mL. The antimicrobial agents used were considered bactericidal at the lowest concentration that reduced the original inoculum by 3 log10 CFU/mL (99.9%) for each of the indicated times. However, they were designated bacteriostatic if the inoculum was reduced by only 0–3 log10 CFU/mL. To confirm the results, the time-kill assays for each experiment were performed at least three times; the data are represented as mean data or standard deviation (SD).
Western blotting
The MRSA culture (ATCC 33591) was grown at OD600 of 0.4 in MHB for Western blot analysis. The cellular protein extracts were prepared from the bacterial harvest cells collected after 30 min of the treatment. The harvested cells from the bacterial culture were suspended in lysis buffer containing Tris-HCI (pH 7.5), and separated soluble protein was extracted by centrifugation at 10,000×g for 15 min. The cell lysate was in the supernatant. The protein concentrations were measured via the Bio-Rad (Hercules, CA) protein assay. The supernatant was transferred to a tube and 10 μL each of the soluble fraction for sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and Western blotting were used to determine the solubility of the recombinant protein of interest. Aliquots of the cellular proteins were then electrophoresed on 10% SDS-PAGE. In Western blot analyses, electrophoresed gels were transferred to Amersham Hybond™-P-membrane (GE Healthcare, Seoul, Korea). The membrane was probed with monoclonal mouse anti-PBP2a primary antibody (diluted 1:500; DiNonA Inc.) and was reprobed with anti-mouse immunoglobulin G secondary antibody (diluted 1:1000; Enzo Life Sciences, Farmingdale, NY). Loading differences were normalized with monoclonal anti-glyceraldehyde-3-phosphate dehydrogenase (diluted 1:500; Santa Cruz Biotechnology, Inc., Santa Cruz, CA). Western blot bands were visualized using a LAS-4000 mini Chemical Luminescent image.
Transmission electron microscopy (TEM)
MRSA exponential phase cultures were prepared from the overnight cultures incubated in MHB at 37°C until they reached the midlogarithmic phase of the growth. The MHB-grown exponential-phase MRSA strains were treated with one half MIC and MIC of morin for 30 min. Subsequently, 2 mL of each culture was collected by centrifugation at 10,000×g for 10 min. After removal of the supernatant, the pellets were fixed with modified Karnovsky's fixative. The specimens were examined using an energy-filtering transmission electron microscope (LIBRA 120; Carl Zeiss, Oberkochen, Germany) operated at an accelerating voltage of 120 kV. The electronic signals transmitted were recorded using a 4×4 k2 slow-scan charge-coupled device camera (Ultrascan 4000 SP; Gatan, Pleasanton, CA) attached to the electron microscope.
Results
Anti-MRSA activity of morin
Antimicrobial susceptibility tests of morin against six strains of S. aureus were performed using the standard broth microdilution method. The MICs of morin for each of the tested strains are listed in Tables 1 and 2. The growth of S. aureus was inhibited in the range of concentrations from 125 to 500 μg/mL morin.
FICI: <0.5, synergy; FICI: 0.5–0.75, partial synergy.
MIC, minimum inhibitory concentration; FIC, fractional inhibitory concentration; FICI, FIC index.
FICI, <0.5, synergy; FICI, 0.5–0.75, partial synergy.
MIC, minimum inhibitory concentration; FIC, fractional inhibitory concentration; FICI, FIC index.
Increased susceptibility to β-lactam antibiotic in MRSA
The combined effects of morin with β-lactam antibiotics ampicillin and oxacillin were tested on MRSA strains by using the checkerboard dilution assay. The antibacterial effects of morin and morin combined with the antibiotics are listed in Tables 1 and 2. The antibacterial activity of morin markedly reduced the MICs of these antibiotics against S. aureus strains. The MICs of morin in combination with ampicillin and oxacillin were reduced 2- to 8-folds and 2- to 16-folds, respectively.
Time-kill curve assay
The synergistic effects of morin with selected antibiotic oxacillin on MRSA were confirmed by the time-kill curve assay. Figure 2 shows that within a 24-h incubation period neither morin nor oxacillin alone induced the cell death. However, when used together, the combination of morin and oxacillin caused rapid inhibition in a time-dependent process during an observation period of 24 h. The combination of one half MIC morin and one half MIC oxacillin showed a reduction in the bacterial count by 3 log10 CFU/mL (DPS-1 and DPS-3) after 24 h. The enhanced effects of the combination of morin and oxacillin sharply reduced the number of CFUs.
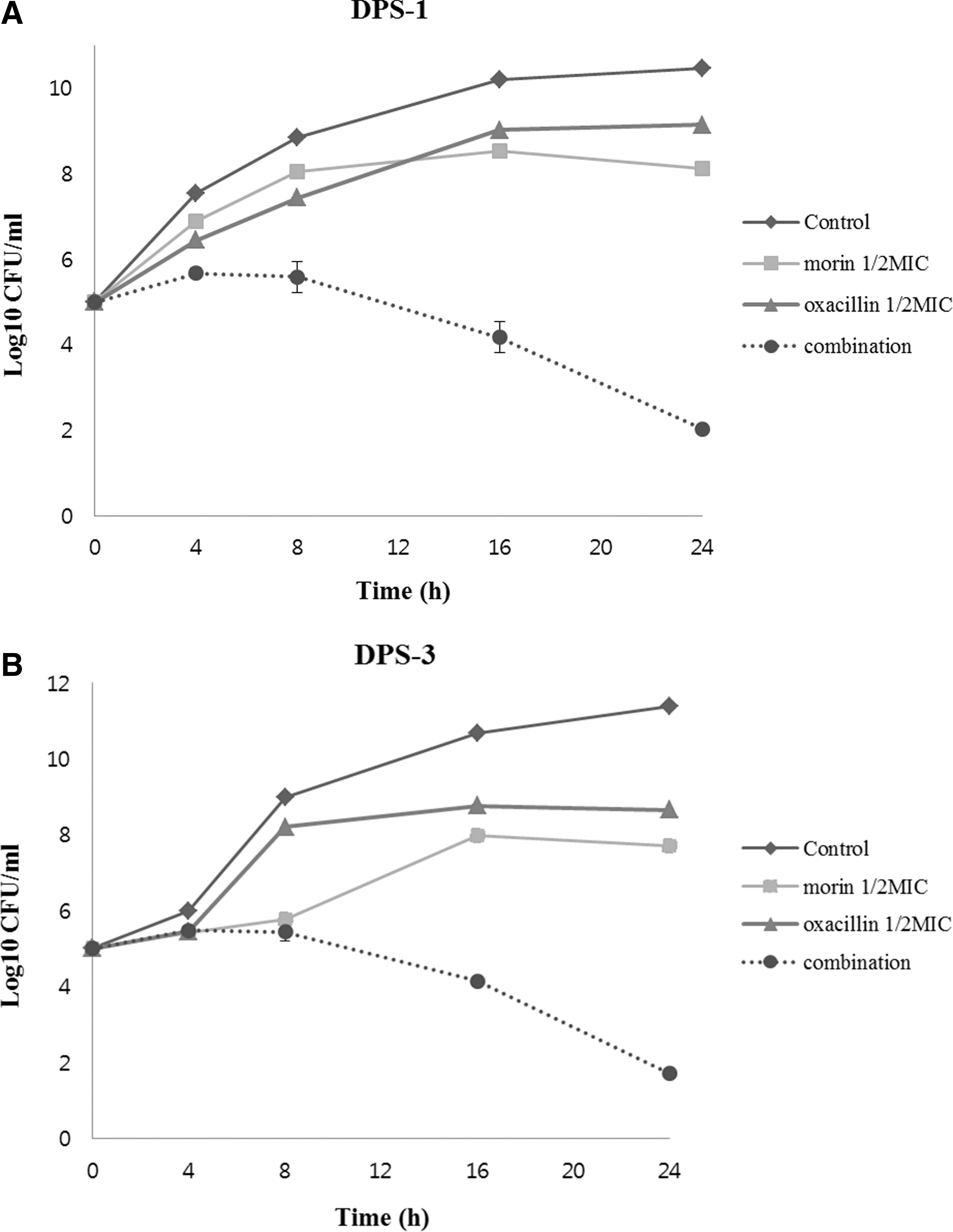
Time-kill curves of methicillin-resistant Staphylococcus aureus (
Expression of the PBP2a determinants in MRSA ATCC 33591 treated with morin
The protein level of PBP2a was analyzed by the Western blotting method. In the Western blot, the PBP2a protein was induced after the induction with oxacillin (32 μg/mL). The protein level was reduced by adding morin at a concentration of 250 μg/mL. The analysis indicated that morin reduced the secretion of PBP2a from MRSA (ATCC 33591), as shown in Figure 3.
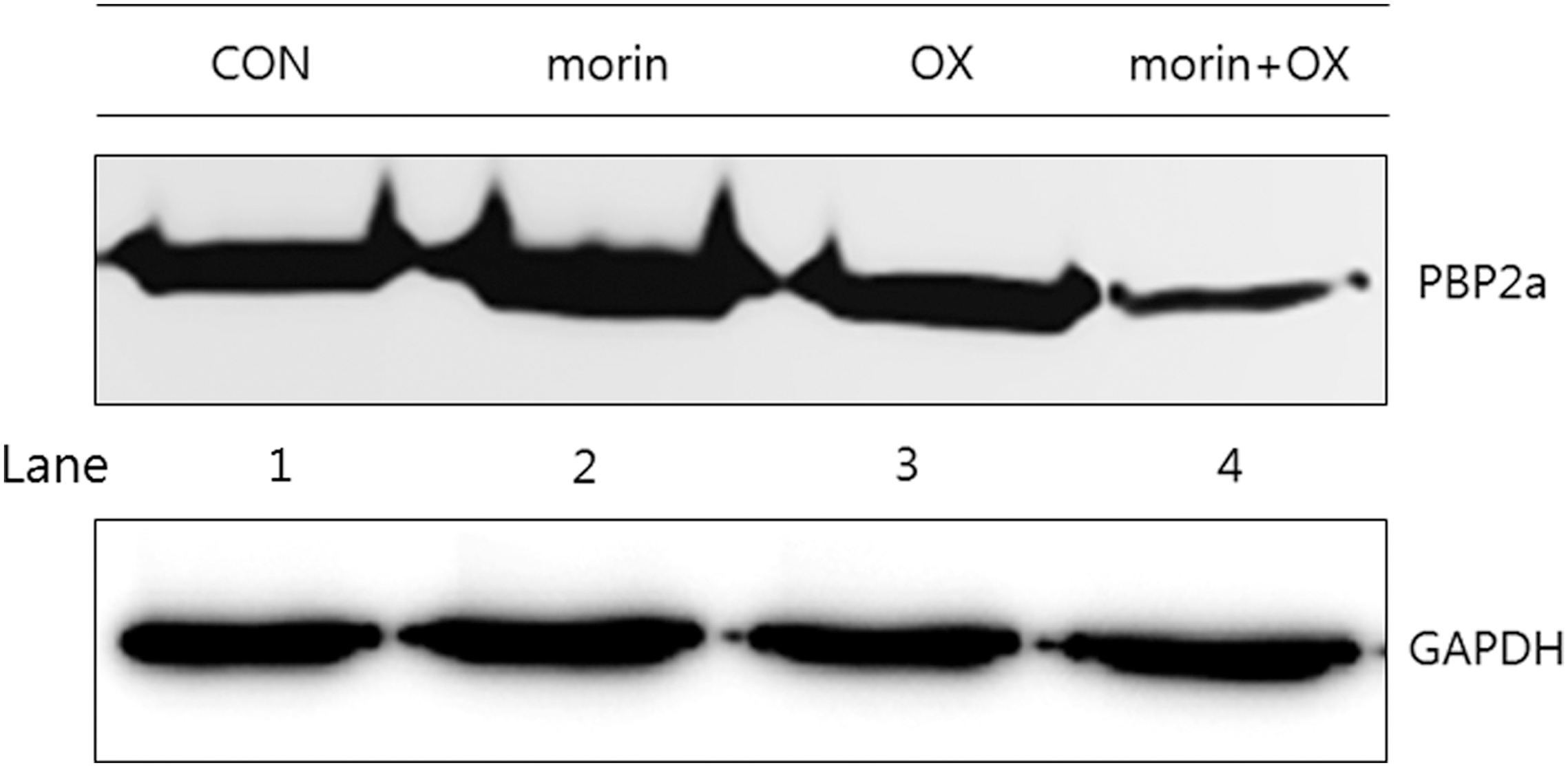
Effect of morin on the expression of penicillin-binding protein 2a (PBP2a) against methicillin-resistant Staphylococcus aureus ATCC 33591. Western blot analysis was used to represent PBP2a production treatment with oxacillin (OX) (32 μg/mL) and morin (250 μg/mL) alone (lane 2 and 3) and in combination (lane 4). Control (CON) was used without treatment drugs (lane 1). GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
TEM images of the MRSA strains
The TEM images of the MRSA strains confirmed a change in the ultrastructure of the bacteria following the exposure to morin. In the untreated MRSA strains, intact septa were evident. After exposure to the one half MIC of the morin, membrane damage and disruption of the midline of the MRSA strains were evident. The MIC of the morin-exposed MRSA cells appeared irregular, rougher, and partially enlarged with cytoplasmic disruption (Fig. 4).
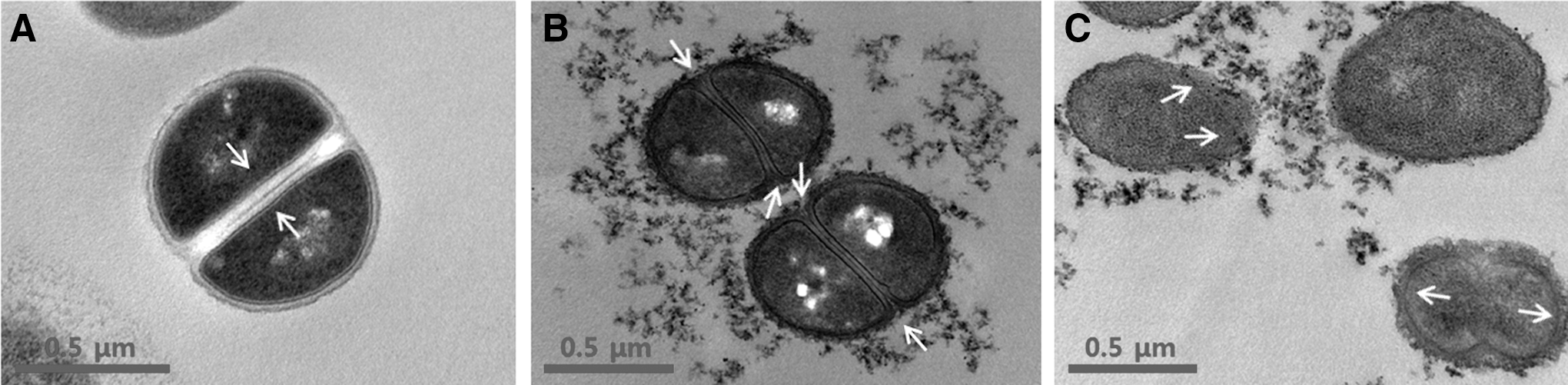
Transmission electron microscopy images of methicillin-resistant Staphylococcus aureus (MRSA) after 30 min of morin treatment.
Statistical analysis
All experiments were performed more than three times. These data from the experiments were presented as the mean±SEM.
Discussion
In this study, the in vitro ability of the combination of morin with ampicillin and oxacillin to kill MRSA isolates was analyzed by the viability assay and Western blot assay methods. The antibacterial mechanism of β-lactam antibiotic was found to be linked to the inhibition of cell-wall synthesis by direct binding to PBP by inactivating its transpeptidase activity (Kitano and Tomasz, 1979; Brudzynski and Sjaarda, 2014), thereby generating spheroplast phenotype with upregulated expression of the genes involved in spheroplast lysis (Brudzynski and Sjaarda, 2014). However, β-lactam antibiotics exhibit activity against the Gram-positive organisms, because some organisms produce β-lactamase enzyme that breaks the β-lactam ring, thereby destroying the activity of the antibiotic (Nester et al., 1998). Therefore, our prevailing purpose was to examine an alternative anti-MRSA drug that can reduce the antibiotic resistance to prevent or treat MRSA diseases.
The results of the bacterial growth curve assay indicate that the combination of morin with oxacillin remarkably suppressed the growth of MRSA. The drastic reduction in the cell viability test clearly showed that the combination of oxacillin and morin can reverse the β-lactam antibiotic resistance in a MRSA. Hence, we expected that morin affects the level of PBP2a protein, encoded by the gene mecA. In the Western blot analysis, morin together with oxacillin lowered the expression of the PBP2a protein in MRSA. When the PBP2a level is lowered or inactive, MRSA is not protected against morin, and MRSA is killed. Therefore, this study confirms that morin potentiates the effect of β-lactam antibiotic against MRSA.
The morphological analyses of the morin-induced MRSA have shown concentration-dependent phenotype changes. Morin affected the cell wall and/or cell membrane. Antibiotic treatment is known to cause cellular changes such as membrane damage and extrusion of cytoplasmic contents (Lehrer et al., 2003; Al-Habib et al., 2010). This study indicates that plant flavonoid morin has potent antibacterial effect on MRSA strains, and this activity potentiated β-lactam antibiotic activity against MRSA. Our forthcoming focus will be to elucidate the mechanism of the increase in the susceptibility of morin to β-lactam antibiotic on MRSA.
Footnotes
Acknowledgment
This study was supported by the Basic Science Research Program through the National Research Foundation of Korea (NRF), funded by the Ministry of Education, Science and Technology (2012-0004337), and by the “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ00962202)” Rural Development Administration, Korea.
Disclosure Statement
No competing financial interests exist.